Recombinant Expression of a Novel Fungal Immunomodulatory Protein with Human Tumor Cell Antiproliferative Activity from Nectria haematococca

Abstract
:1. Introduction
2. Results and Discussion
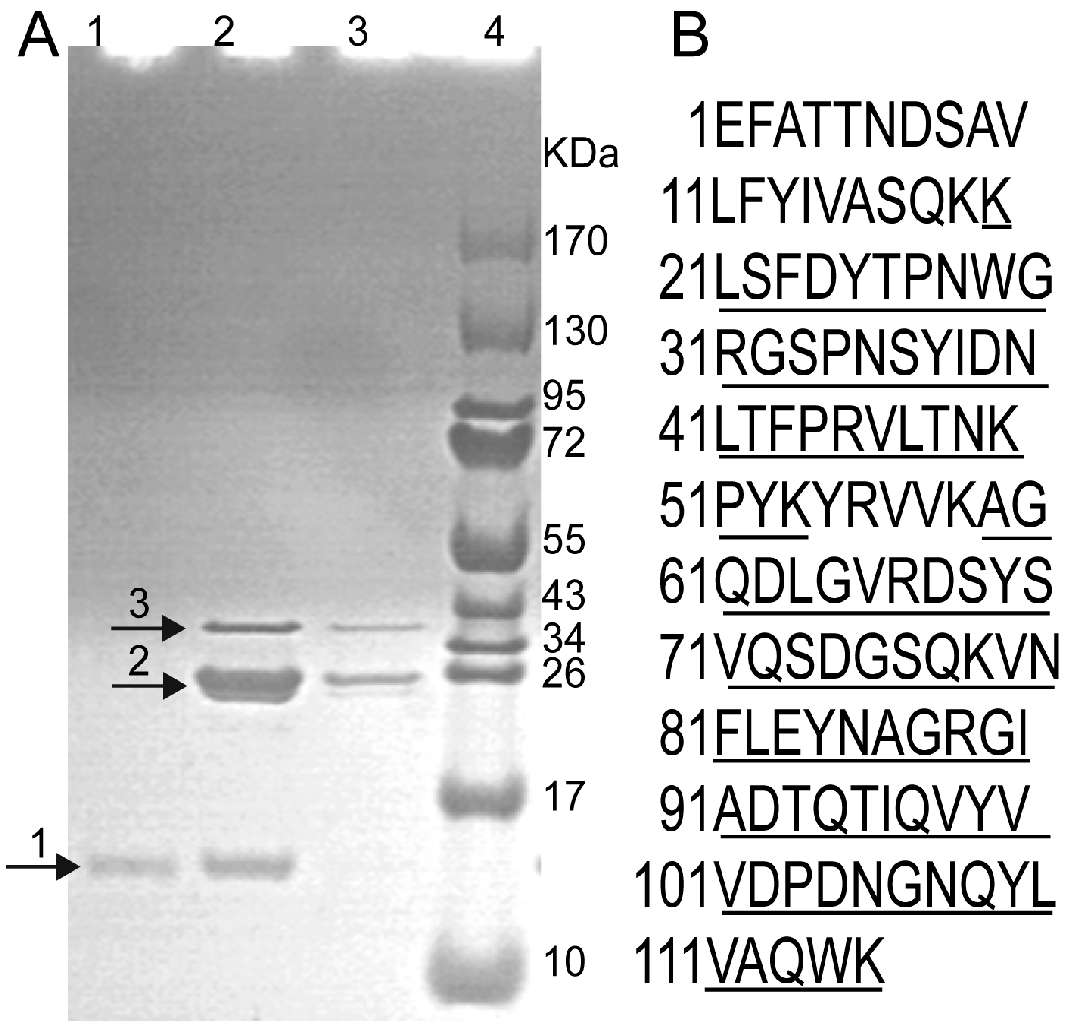
2.1. Expression, Purification and Identification of rFIP-nha

2.2. Hemagglutination Activity of rFIP-nha
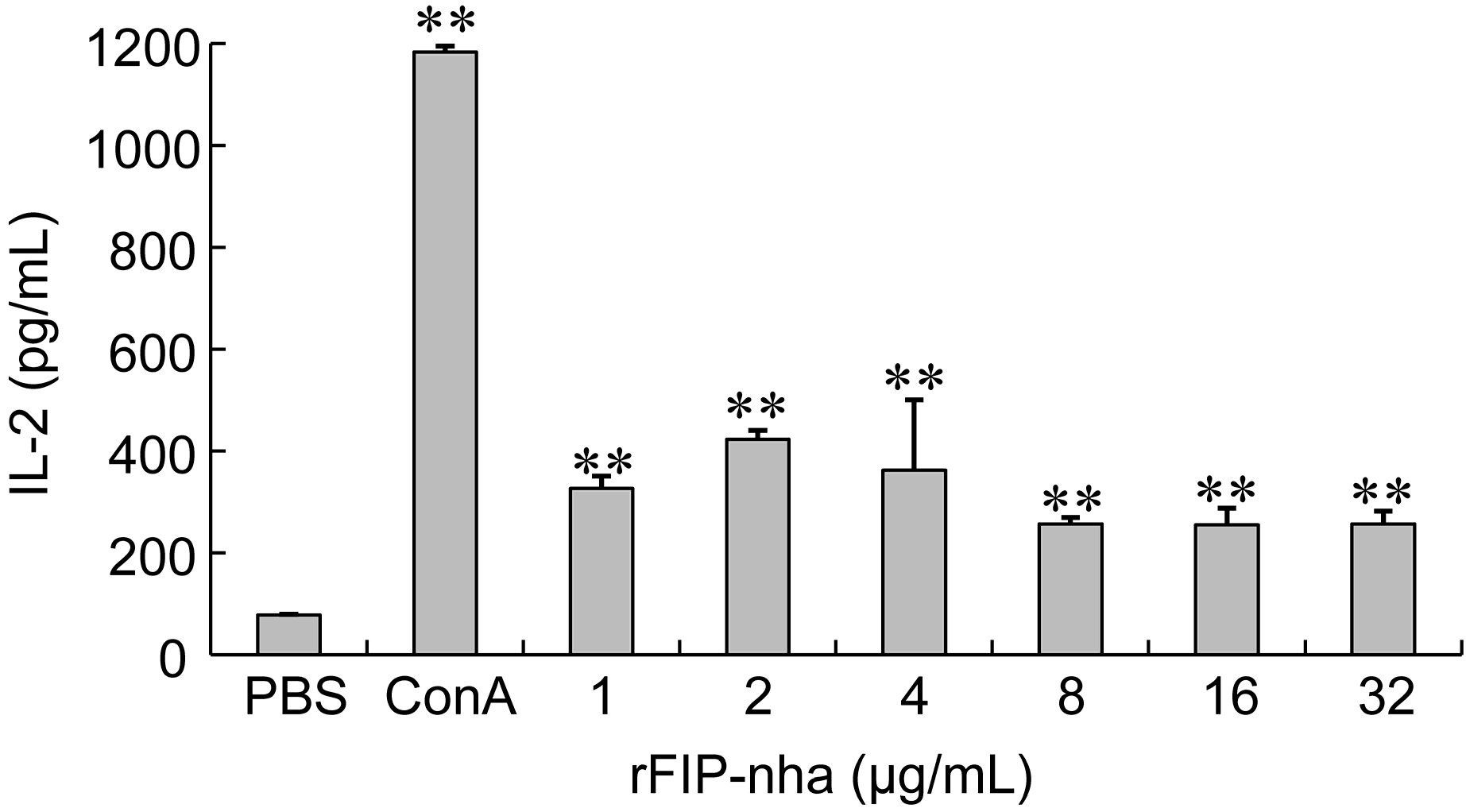
2.3. The Immunomodulatory Activity Assay of rFIP-nha

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Concentration (μg/mL) | PBS (A570) | ConA (A570) | LPS (A570) |
|---|---|---|---|---|
| Control | - | 0.16 ± 0.01 | 0.33 ± 0.03 | 0.32 ± 0.01 |
| rFIP-nha | 1 | 0.26 ± 0.02 a | 0.35 ± 0.01 | 0.38 ± 0.02 |
| 2 | 0.33 ± 0.02 b | 0.41 ± 0.00 a | 0.43 ± 0.00 a | |
| 4 | 0.31 ± 0.01 b | 0.40 ± 0.00 a | 0.40 ± 0.01 a | |
| 8 | 0.25 ± 0.00 a | 0.35 ± 0.00 | 0.35 ± 0.01 | |
| 16 | 0.25 ± 0.00 a | 0.34 ± 0.00 | 0.34 ± 0.00 | |
| 32 | 0.24 ± 0.01 a | 0.33 ± 0.01 | 0.33 ± 0.02 |

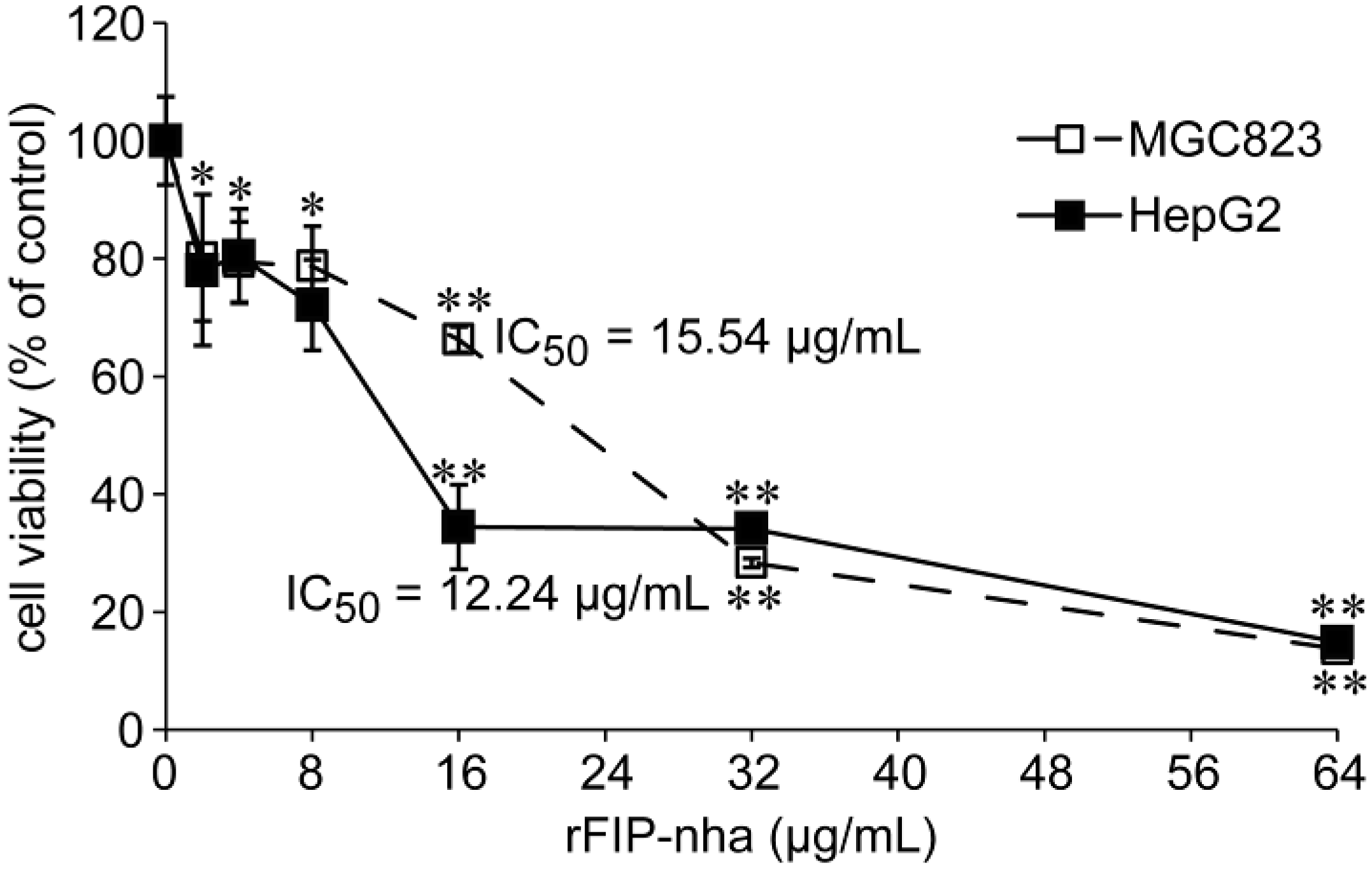
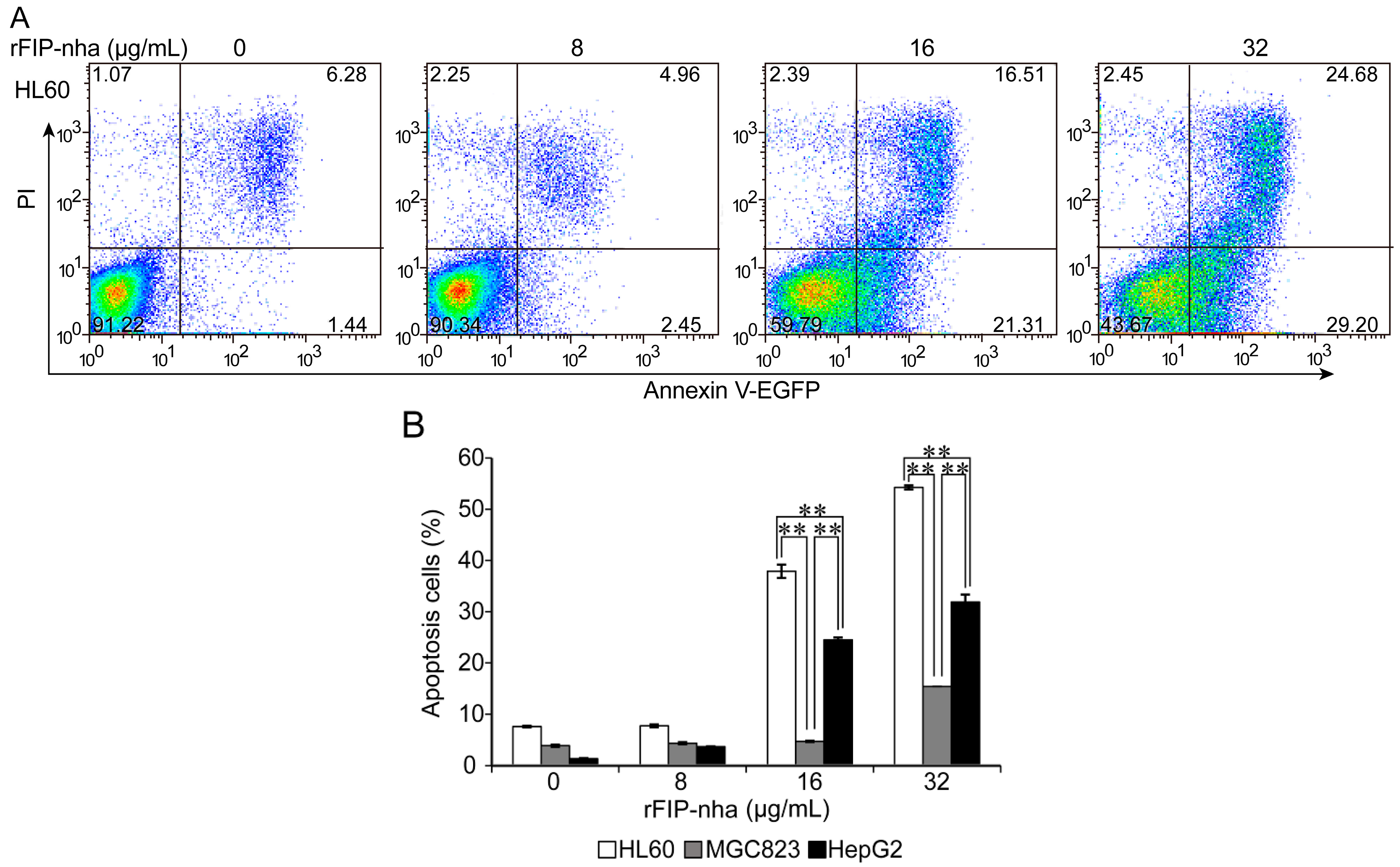
2.4. The Antitumor Activity Assay of rFIP-nha on Human Tumor Cells


3. Experimental Section
3.1. Expression and Purification of rFIP-nha
3.1.1. Gene Synthesis and Construction of the Expression Vector
3.1.2. Protein Expression and Purification
3.1.3. MALDI-MS Analysis
3.2. Hemagglutination Test
3.3. The Immunomodulatory Activity Assay of rFIP-nha
3.3.1. Mouse Splenocyte Proliferation Assay
3.3.2. IL-2 Release Assay
3.4. The Antitumor Activity Assay of rFIP-nha
3.4.1. Tumor Cell Proliferative Inhibitory Analysis
3.4.2. Tumor Cell Apoptosis Assay
3.5. Statistical Analysis
4. Conclusions
Supplementary Materials
Supplementary Files
Supplementary File 1Acknowledgments
Conflicts of Interest
References
- Risberg, T.; Lund, E.; Wist, E.; Kaasa, S.; Wilsgaard, T. Cancer patients’ use of nonproven therapy: A 5-year follow-up study. J. Clin. Oncol. 1998, 16, 6–12. [Google Scholar]
- Wang, S.Y.; Hsu, M.L.; Hsu, H.C.; Tzeng, C.H.; Lee, S.S.; Shiao, M.S.; Ho, C.K. The anti-tumor effect of Ganoderma lucidum is mediated by cytokines released from activated macrophages and T-lymphocytes. Int. J. Cancer 1997, 70, 699–705. [Google Scholar] [CrossRef]
- Zhu, M.; Chang, Q.; Wong, L.K.; Chong, F.S.; Li, R.C. Triterpene antioxidants from Ganoderma lucidum. Phytother. Res. 1999, 13, 529–531. [Google Scholar] [CrossRef]
- Ko, J.L.; Hsu, C.I.; Lin, R.H.; Kao, C.L.; Lin, J.Y. A new fungal immunomodulatory protein, FIP-fve isolated from the edible mushroom, Flammulina velutipes and its complete amino acid sequence. Eur. J. Biochem. 1995, 228, 244–249. [Google Scholar] [CrossRef]
- Kino, K.; Yamashita, A.; Yamaoka, K.; Watanabe, J.; Tanaka, S.; Ko, K.; Shimizu, K.; Tsunoo, H. Isolation and characterization of a new immunomodulatory protein, Ling Zhi-8 (LZ-8) from Ganoderrna zucidiurn. J. Biol. Chem. 1989, 264, 472–478. [Google Scholar]
- Hsu, H.C.; Hsu, C.I.; Lin, R.H.; Kao, C.L.; Lin, J.Y. Fip-vvo, a new fungal immunomodulatory protein isolated from Volvariella volvacea. Biochem. J. 1997, 323, 557–565. [Google Scholar]
- Paaventhan, P.; Jeremiah, S.J.; See, V.S.; Shai, V.; Howard, R.; Kaw, Y.C.; Prasanna, R.K. A 1.7Å structure of Fve, a member of the new fungal immunomodulatory protein family. J. Mol. Biol. 2003, 332, 461–470. [Google Scholar] [CrossRef]
- Huang, L.; Sun, F.; Liang, C.Y.; He, Y.X.; Bao, R.; Liu, L.X.; Zhou, C.Z. Crystal structure of LZ-8 from the medicinal fungus Ganoderma lucidium. Proteins 2008, 75, 524–527. [Google Scholar] [CrossRef]
- Van der Hem, L.G.; van der Vliet, J.A.; Frans, C.; Bocken, M.; Kino, K.; Hoitsma, A.J.; Tax, W.J.M. Ling Zhi-8: studies of a new immunomodulating agent. Transplantation 1995, 60, 438–443. [Google Scholar]
- Murasugi, A.; Tanaka, S.; Komiyama, N.; Iwata, N.; Kino, K.; Tsunoo, H.; Sakuma, S. Molecular cloning of a cDNA and a gene encoding an immunomodulatory protein, Ling Zhi-8, from a fungus, Ganoderma lucidium. J. Biol. Chem. 1991, 266, 2486–2493. [Google Scholar]
- Haak-Frendscho, M.; Kino, K.; Sone, T.; Jardieu, P. Ling Zhi-8: A novel T cell mitogen induces cytokine production and upregulation of ICAM-1 expression. Cell Immunol. 1993, 150, 101–113. [Google Scholar] [CrossRef]
- Wang, P.H.; Hsu, C.I.; Tang, S.C.; Huang, Y.L.; Lin, J.Y.; Ko, J.L. Fungal immunomodulatory protein from Flammulina velutipes induces interferon-gamma production through p38 mitogen-activated protein kinase signaling pathway. J. Agric. Food Chem. 2004, 52, 2721–2725. [Google Scholar] [CrossRef]
- Liao, C.H.; Hsiao, Y.M.; Hsu, C.P.; Lin, M.Y.; Wang, J.C.; Huang, Y.L.; Ko, J.L. Transcriptionally mediated inhibition of telomerase of fungal immunomodulatory protein from Ganoderma tsugae in A549 human lung adenocarcinoma cell line. Mol. Carcinog. 2006, 45, 220–229. [Google Scholar] [CrossRef]
- Liao, C.H.; Hsiao, Y.M.; Sheu, G.T.; Chang, J.T.; Wang, P.H.; Wu, M.F.; Shieh, G.J.; Ko, J.L. Nuclear translocation of telomerase reverse transcriptase and calcium signaling in repression of telomerase activity in human lung cancer cells by fungal immunomodulatory protein from Ganoderma tsugae. Biochem. Pharmacol. 2007, 74, 1541–1554. [Google Scholar] [CrossRef]
- Chang, H.H.; Hsieh, K.Y.; Yeh, C.H.; Tu, Y.P.; Sheu, F. Oral administration of an Enoki mushroom protein FVE activates innate and adaptive immunity and induces anti-tumor activity against murine hepatocellular carcinoma. Int. Immunopharm. 2010, 10, 239–246. [Google Scholar] [CrossRef]
- Lin, W.H.; Huang, C.H.; Hsu, C.I.; Lin, J.Y. Dimerization of the N-terminal amphipathic α-helix domain of the fungal immunomodulatory protein from Ganoderma tsugae (Fip-gts) defined by a yeast two-hybrid system and site-directed mutagenesis. J. Biol. Chem. 1997, 272, 20044–20048. [Google Scholar] [CrossRef]
- Lin, Y.W.; Yeh, C.H.; Huang, Y.H.; Lai, Y.C.; Chang, J.G.; Ko, J.L. A new immunomodulatory protein from Ganoderma microsporum inhibits epidermal growth factor mediated migration and invasion in A549 lung cancer cells. Process Biochem. 2010, 45, 1537–1542. [Google Scholar] [CrossRef]
- Chang, H.H.; Yeh, C.H.; Sheu, F. A novel immunomodulatory protein from Poria cocos induces Toll-like receptor 4-dependent activation within mouse peritoneal macrophages. J. Agric. Food Chem. 2009, 57, 6129–6139. [Google Scholar]
- Li, F.; Wen, H.; Liu, X.; Zhou, F.; Chen, G. Gene cloning and recombinant expression of a novel fungal immunomodulatory protein from Trametes versicolor. Protein Expr. Purif. 2012, 82, 339–344. [Google Scholar] [CrossRef]
- Li, Q.Z.; Wang, X.F.; Zhou, X.W. Recent status and prospects of the fungal immunomodulatory protein family. Crit. Rev. Biotechnol. 2011, 31, 365–375. [Google Scholar] [CrossRef]
- Wang, X.F.; Su, K.Q.; Bao, T.W.; Cong, W.R.; Chen, Y.F.; Li, Q.Z.; Zhou, X.W. Immunomodulatory effects of fungal proteins. Curr. Top. Nutraceutical Res. 2012, 10, 1–11. [Google Scholar]
- Bastiaan-Net, S.; Chanput, W.; Hertz, A.; Zwittink., R.D.; Mes, J.J.; Wichers, H.J. Biochemical and functional characterization of recombinant fungal immunomodulatory proteins (rFIPs). Int. Immunopharmacol. 2013, 15, 167–175. [Google Scholar] [CrossRef]
- Beigia, L.; Karbalaei-Heidaria, H.R.; Kharrati-Kopaeib, M. Optimization of an extracellular zinc-metalloprotease (SVP2) expression in Escherichia coli BL21 (DE3) using response surface methodology. Protein Expr. Purif. 2012, 84, 161–166. [Google Scholar] [CrossRef]
- Qin, X.; Ding, Y.X.; Shang, C.H.; Jiang, C.; Zhao, M.W. Functional expression of LZ8, a fungal immunomodulatory protein from Ganoderma lucidium in Pichia pastoris. J. Gen. Appl. Microbiol. 2008, 54, 393–398. [Google Scholar] [CrossRef]
- Jiao, L.; Li, X.; Li, T.; Jiang, P.; Zhang, L.; Wu, M. Characterization and antitumor activity of alkali-extracted polysaccharide from Enteromorpha intestinalis. Int. Immunopharmacol. 2009, 9, 324–329. [Google Scholar] [CrossRef]
- Wermerskirchen, A.S.; LaTocha, D.H.; Clarke, B.L. Adrenocorticotropic hormone controls Concanavalin A activation of rat lymphocytes by modulating IL-2 production. Life Sci. 2000, 67, 2177–2187. [Google Scholar] [CrossRef]
- Wu, C.M.; Wu, T.Y.; Kao, S.S.; Ko, J.L.; Jinn, T.R. Expression and purification of a recombinant Fip-fve protein from Flammulina velutipes in baculovirus-infected insect cells. J. Appl. Microbiol. 2008, 104, 1354–1362. [Google Scholar] [CrossRef]
- Jinn, T.R.; Wu, C.M.; Tu, W.C.; Ko, J.L.; Tzen, J.T. Functional expression of FIP-gts, a fungal immunomodulatory protein from Ganoderma tsugae in Sf21 insect cells. Biosci. Biotechnol. Biochem. 2006, 70, 2627–2634. [Google Scholar] [CrossRef]
- Ou, C.C.; Hsiao, Y.M.; Wu, W.J.; Tasy, G.J.; Ko, J.L.; Lin, M.Y. FIP-fve stimulates interferongamma production via modulation of calcium release and PKC-alpha activation. J. Agric. Food Chem. 2009, 57, 11008–11013. [Google Scholar] [CrossRef]
- Ou, C.C.; Hsiao, Y.M.; Wang, W.H.; Ko, J.L.; Lin, M.Y. Stability of fungal immunomodulatory protein, FIP-gts and FIP-fve, in IFN-γ production. Food Agric. Immunol. 2009, 20, 319–332. [Google Scholar] [CrossRef]
- Li, Q.Z.; Wang, X.F.; Chen, Y.Y.; Lin, J.; Zhou, X.W. Cytokines expression induced by Ganoderma sinensis Fungal Immunomodulatory Proteins (FIP-gsi) in Mouse Spleen Cells. Appl. Biochem. Biotechnol. 2010, 162, 1403–1413. [Google Scholar] [CrossRef]
- Li, Q.Z.; Huang, L.; Wang, X.F.; Li, X.S.; Wu, S.Q.; Zhou, X.W. Flammulina velutipes induces cytokine gene expression in mouse spleen cells. Curr. Top. Nutraceut. Res. 2011, 9, 111–118. [Google Scholar]
- Liang, C.Y.; Zhang, S.Q.; Liu, Z.Y.; Sun, F. Ganoderma lucidum immunomodulatory protein (Lz-8) expressed in Pichia pastoris and the identification of immunocompetence. Chin. J. Biotech. 2009, 25, 441–447. [Google Scholar]
- Lin, J.W.; Hao, L.X.; Xu, G.X.; Sun, F.; Gao, F.; Zhang, R.; Liu, L.X. Molecular cloning and recombinant expression of a gene encoding a fungal immunomodulatory protein from Ganoderma lucidum in Pichia pastoris. World J. Microbiol. Biotechnol. 2009, 25, 383–390. [Google Scholar] [CrossRef]
- Wu, C.T.; Lin, T.Y.; Hsu, H.Y.; Sheu, F.; Ho, C.M.; Chen, E.I. Ling Zhi-8 mediates p53-dependent growth arrest of lung cancer cells proliferation via the ribosomal protein S7-MDM2-p53 pathway. Carcinogenesis 2011, 32, 1890–1896. [Google Scholar] [CrossRef]
- Lin, C.C.; Yu, Y.L.; Shih, C.C.; Liu, K.J.; Ou, K.L.; Hong, L.Z.; Chen, J.D.; Chu, C.L. A novel adjuvant Ling Zhi-8 enhances the efficacy of DNA cancer vaccine by activating dendriticcells. Cancer Immunol. Immunother. 2011, 60, 1019–1027. [Google Scholar] [CrossRef]
- Liao, C.H.; Hsiao, Y.M.; Lin, C.H.; Yeh, C.S.; Wang, J.C.; Ni, C.H.; Hsu, C.P.; Ko, J.L. Induction of premature senescence in human lung cancer by fungal immunomodulatory protein from Ganoderma tsugae. Food Chem. Toxicol. 2008, 46, 1851–1859. [Google Scholar] [CrossRef]
- Lin, C.H.; Hsiao, Y.M.; Ou, C.C.; Lin, Y.W.; Chiu, Y.L.; Lue, K.H.; Chang, J.G.; Ko, J.L. GMI, a Ganoderma immunomodulatory protein, down-regulates tumor necrosis factor alpha-induced expression of matrix metal loproteinase 9 via NF-kappaB pathway in human alveolar epithelial A549 cells. J. Agric. Food Chem. 2010, 58, 12014–12021. [Google Scholar] [CrossRef]
- Chang, Y.C.; Hsiao, Y.M.; Wu, M.F.; Ou, C.C.; Lin, Y.W.; Lue, K.H.; Ko, J.L. Interruption of lung cancer cell migration and proliferation by fungal immunomodulatory protein FIP-fve from Flammulina velutipes. J. Agric. Food Chem. 2013, 61, 12044–12052. [Google Scholar]
- Marshall, N.A.; Galvin, K.C.; Corcoran, A.M.; Boon, L.; Higgs, R.; Mills, K.H. Immunotherapy with PI3K inhibitor and Toll-like receptor agonist induces IFN-gamma+IL-17+ polyfunctional T cells that mediate rejection of murine tumors. Cancer Res. 2012, 72, 581–591. [Google Scholar] [CrossRef]
- Li, F.; Wen, H.; Zhang, Y.; Aa, M.; Liu, X. Purification and characterization of a novel immunomodulatory protein from the medicinal mushroom Trametes versicolor. Sci. China Life Sci. 2011, 54, 379–385. [Google Scholar]
- Wu, T.Y.; Chen, H.A.; Li, F.Y.; Lin, C.T.; Wu, C.M.; Hsieh, F.C.; Tzen, J.T.; Hsieh, S.K.; Ko, J.L.; Jinn, T.R. High-level expression, purification and production of the fungal immunomodulatory protein-gts in baculovirus-infected insect larva. Appl. Biochem. Biotechnol. 2013, 169, 976–989. [Google Scholar] [CrossRef]
- Kong, X.; Zhang, J.; Han, X.; Zhang, P.; Dai, X.; Liu, J.; Zhang, X.; Lee, I.; Liu, S. High-yield production in Escherichia coli of fungal immunomodulatory protein isolated from Flammulina velutipes and its bioactivity assay in vivo. Int. J. Mol. Sci. 2013, 14, 2230–2241. [Google Scholar]
- Ko, J.L.; Lin, S.J.; Hsu, C.I.; Kao, C.L.; Lin, J.Y. Molecular cloning and expression of a fungal immunomodulatory protein, FIP-fve, from Flammulina velutipes. J. Formos Med. Assoc. 1997, 96, 517–524. [Google Scholar]
- Strong, D.M.; Ahmed, A.A.; Thurman, G.B.; Sell, K.W. In vitro stimulation of murine spleen cells using a micro culture system and a multiple automated sample harvester. J. Immunol. Methods 1973, 2, 279–291. [Google Scholar] [CrossRef]
- Peng, Y.R.; Li, Y.B.; Liu, X.D.; Zhang, J.F.; Duan, J.A. Antitumor activity of C-21 steroidal glycosides from Cynanchum auriculatum Royle ex Wight. Phytomedicine 2008, 15, 1016–1020. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Nie, Y.; Ding, Y.; Shi, L.; Tang, X. Recombinant Expression of a Novel Fungal Immunomodulatory Protein with Human Tumor Cell Antiproliferative Activity from Nectria haematococca. Int. J. Mol. Sci. 2014, 15, 17751-17764. https://doi.org/10.3390/ijms151017751
Li S, Nie Y, Ding Y, Shi L, Tang X. Recombinant Expression of a Novel Fungal Immunomodulatory Protein with Human Tumor Cell Antiproliferative Activity from Nectria haematococca. International Journal of Molecular Sciences. 2014; 15(10):17751-17764. https://doi.org/10.3390/ijms151017751
Chicago/Turabian StyleLi, Shuying, Ying Nie, Yang Ding, Lijun Shi, and Xuanming Tang. 2014. "Recombinant Expression of a Novel Fungal Immunomodulatory Protein with Human Tumor Cell Antiproliferative Activity from Nectria haematococca" International Journal of Molecular Sciences 15, no. 10: 17751-17764. https://doi.org/10.3390/ijms151017751
APA StyleLi, S., Nie, Y., Ding, Y., Shi, L., & Tang, X. (2014). Recombinant Expression of a Novel Fungal Immunomodulatory Protein with Human Tumor Cell Antiproliferative Activity from Nectria haematococca. International Journal of Molecular Sciences, 15(10), 17751-17764. https://doi.org/10.3390/ijms151017751