Analysis of Human TAAR8 and Murine Taar8b Mediated Signaling Pathways and Expression Profile

 ,
,
Abstract
:1. Introduction
2. Results
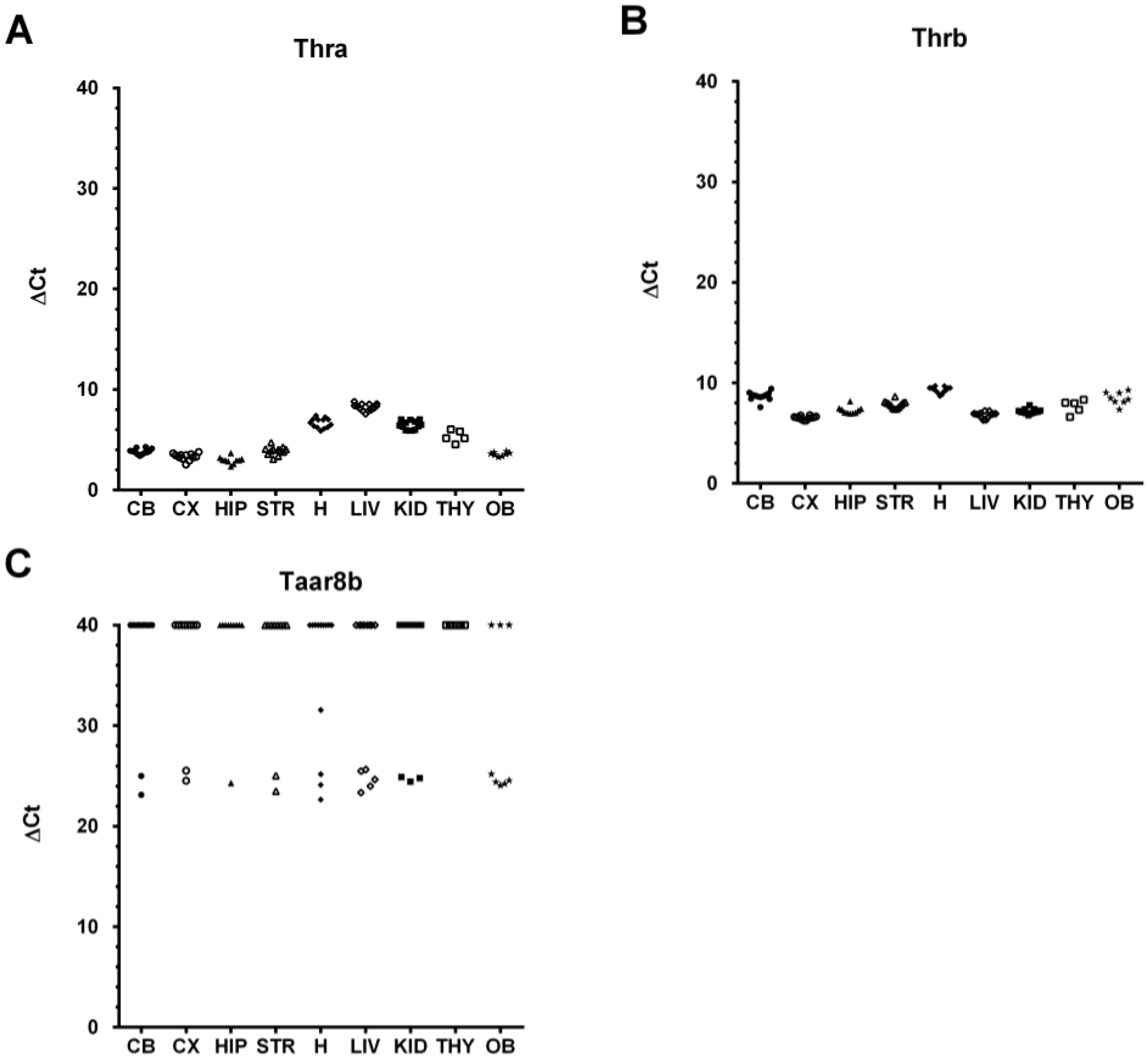
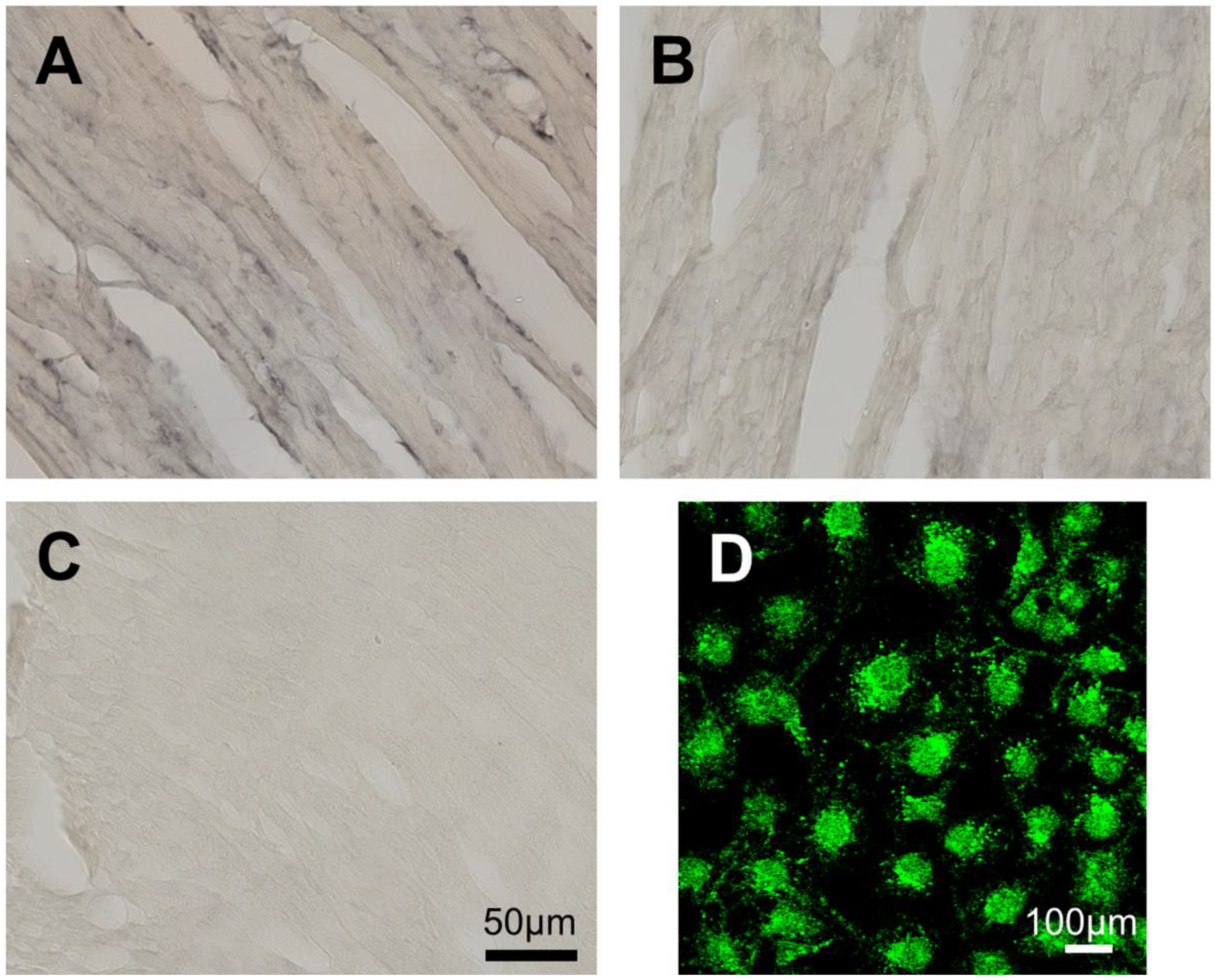
2.1. The Murine Taar8b Is at Most Marginally Expressed in Several Mouse Tissues


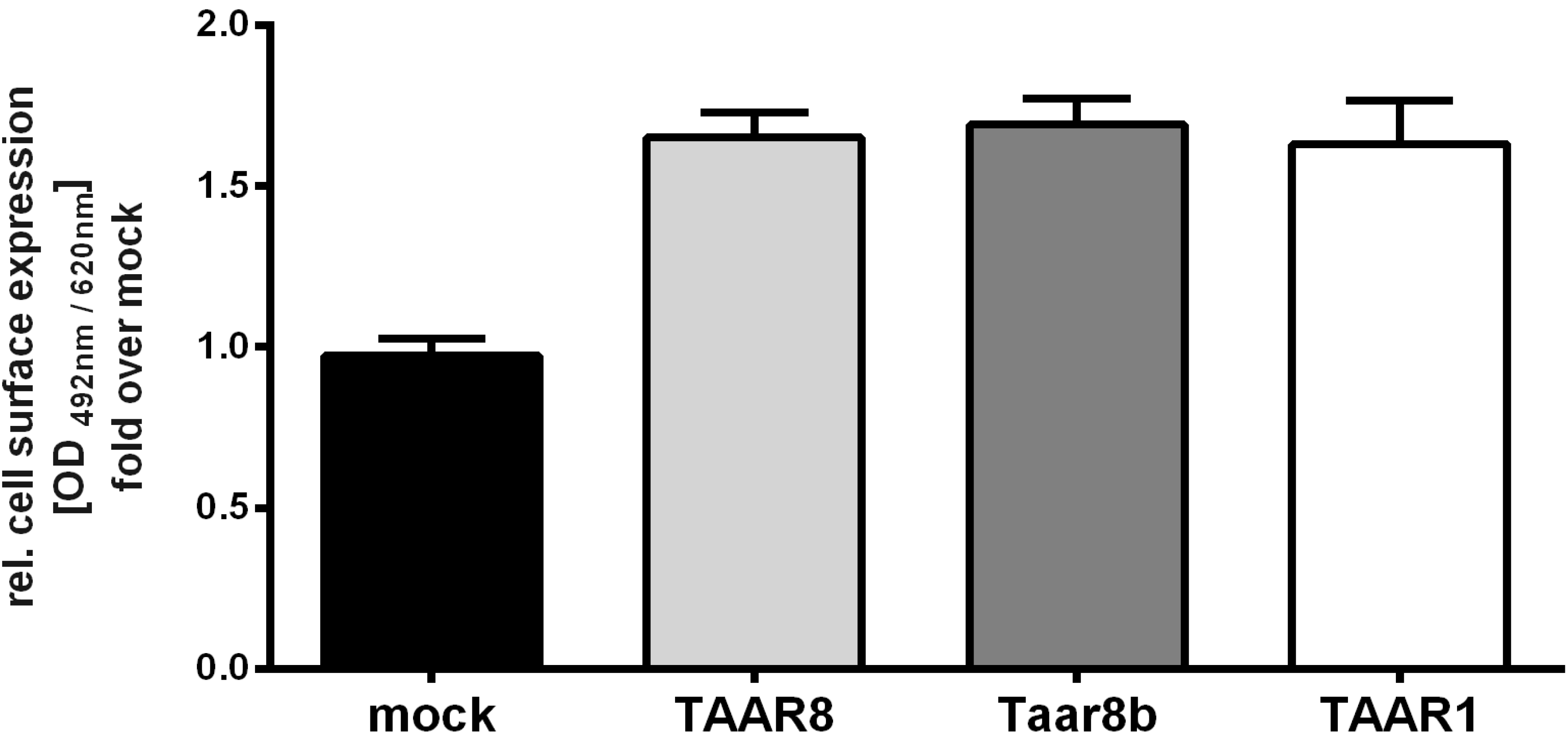
2.2. Cell Surface Expression of TAAR8

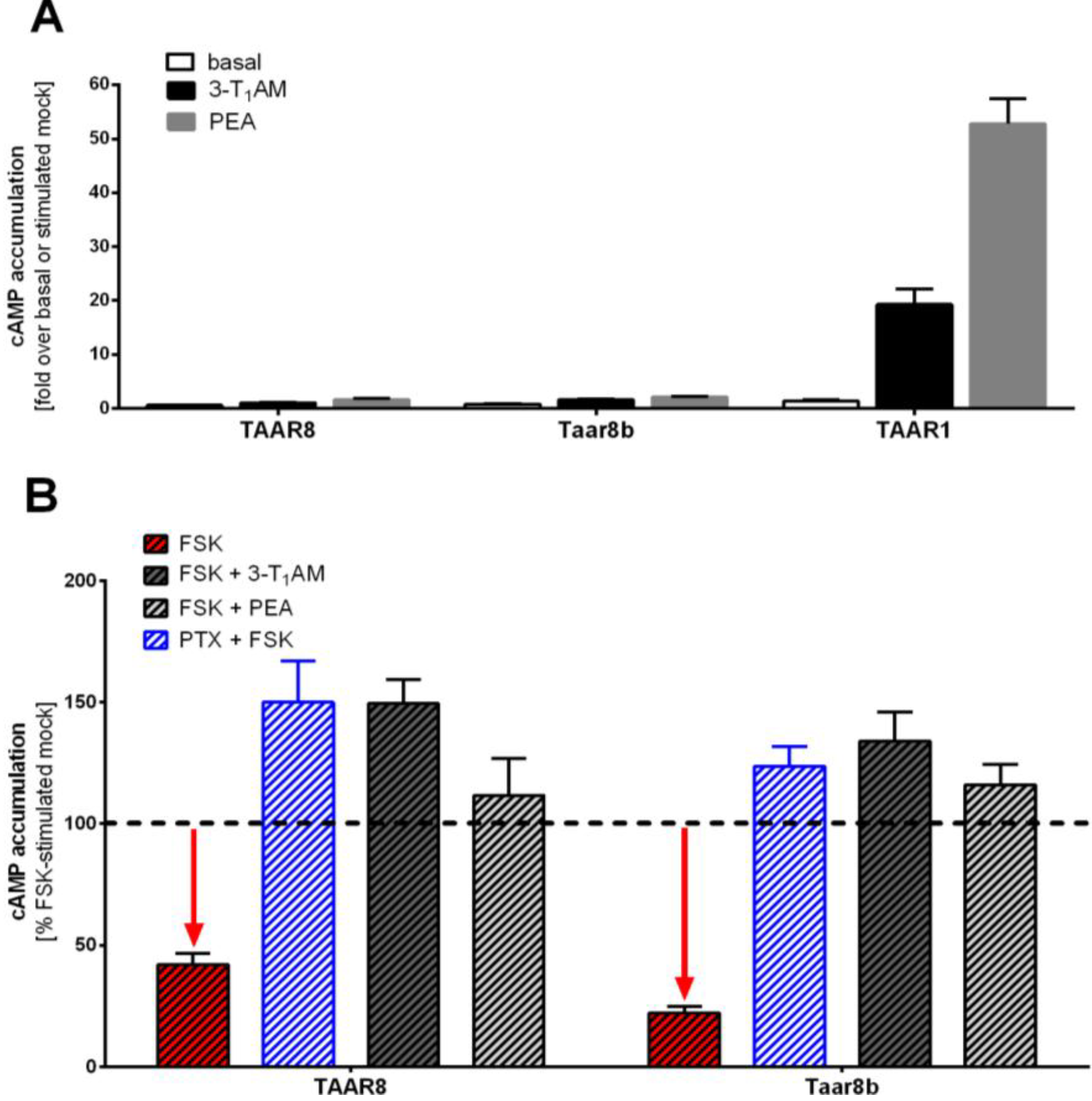
2.3. Identification of Basal Gi/o Mediated Signaling

2.4. G12/13 or Gq/11 Signaling Pathways

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TAAR Subtype | Second Messenger Signaling | Ligand | Basal Activity (Fold Over Basal Mock) | Ligand Stimulated (Fold Over Stimulated Mock) |
|---|---|---|---|---|
| TAAR8 | IP3–luc | 3-T1AM | 1.6 ± 0.1 | 1.1 ± 0.2 |
| PEA | 1.1 ± 0.2 | |||
| RhoA–luc | 3-T1AM | 1.2 ± 0.1 | 1.5 ± 0.3 | |
| PEA | 1.2 ± 0.1 | |||
| Taar8b | IP3–luc | 3-T1AM | 1.2 ± 0.1 | 0.9 ± 0.1 |
| PEA | 1.4 ± 0.2 | |||
| RhoA–luc | 3-T1AM | 1.0 ± 0.1 | 0.8 ± 0.1 | |
| PEA | 0.9 ± 0.1 |
3. Discussion
4. Experimental Section
4.1. Expression Analysis of Taar8b in the Heart by in Situ Hybridization
4.2. Expression Analysis of Taar8b by RT-qPCR
4.3. Cloning of TAAR8
4.4. Cell Culture and Transient Transfection
4.5. Determination of Cell Surface Expression
4.6. Determination of Gs and Gi/o Activation by cAMP Accumulation Assay
4.7. Determination of Gq/11 and G12/13 Activation by Reporter Gene Assay
4.8. Statistical Data Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Grandy, D.K. Trace amine-associated receptor 1-family archetype or iconoclast? Pharmacol. Ther. 2007, 116, 355–390. [Google Scholar]
- Zucchi, R.; Chiellini, G.; Scanlan, T.S.; Grandy, D.K. Trace amine-associated receptors and their ligands. Br. J. Pharmacol. 2006, 149, 967–978. [Google Scholar] [CrossRef] [PubMed]
- Lindemann, L.; Ebeling, M.; Kratochwil, N.A.; Bunzow, J.R.; Grandy, D.K.; Hoener, M.C. Trace amine-associated receptors form structurally and functionally distinct subfamilies of novel G protein-coupled receptors. Genomics 2005, 85, 372–385. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Saraiva, L.R.; Korsching, S.I. Positive Darwinian selection and the birth of an olfactory receptor clade in teleosts. Proc. Natl. Acad. Sci. USA 2009, 106, 4313–4318. [Google Scholar] [CrossRef] [PubMed]
- Tessarolo, J.A.; Tabesh, M.J.; Nesbitt, M.; Davidson, W.S. Genomic organization and evolution of the trace amine-associated receptor (TAAR) repertoire in Atlantic salmon (Salmo salar). G3 2014, 4, 1135–1141. [Google Scholar] [CrossRef] [PubMed]
- Staubert, C.; Boselt, I.; Bohnekamp, J.; Rompler, H.; Enard, W.; Schoneberg, T. Structural and functional evolution of the trace amine-associated receptors TAAR3, TAAR4 and TAAR5 in primates. PLoS One 2010, 5, e11133. [Google Scholar] [CrossRef] [PubMed]
- Liberles, S.D.; Buck, L.B. A second class of chemosensory receptors in the olfactory epithelium. Nature 2006, 442, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Ferrero, D.M.; Wacker, D.; Roque, M.A.; Baldwin, M.W.; Stevens, R.C.; Liberles, S.D. Agonists for 13 trace amine-associated receptors provide insight into the molecular basis of odor selectivity. ACS Chem. Biol. 2012, 7, 1184–1189. [Google Scholar] [CrossRef] [PubMed]
- Scanlan, T.S.; Suchland, K.L.; Hart, M.E.; Chiellini, G.; Huang, Y.; Kruzich, P.J.; Frascarelli, S.; Crossley, D.A.; Bunzow, J.R.; Ronca-Testoni, S.; et al. 3-Iodothyronamine is an endogenous and rapid-acting derivative of thyroid hormone. Nat. Med. 2004, 10, 638–642. [Google Scholar] [CrossRef] [PubMed]
- Chiellini, G.; Frascarelli, S.; Ghelardoni, S.; Carnicelli, V.; Tobias, S.C.; DeBarber, A.; Brogioni, S.; Ronca-Testoni, S.; Cerbai, E.; Grandy, D.K.; et al. Cardiac effects of 3-iodothyronamine: A new aminergic system modulating cardiac function. FASEB J. 2007, 21, 1597–608. [Google Scholar] [CrossRef] [PubMed]
- Vanti, W.B.; Nguyen, T.; Cheng, R.; Lynch, K.R.; George, S.R.; O’Dowd, B.F. Novel human G-protein-coupled receptors. Biochem. Biophys. Res. Commun. 2003, 305, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Panas, M.W.; Xie, Z.; Panas, H.N.; Hoener, M.C.; Vallender, E.J.; Miller, G.M. Trace amine associated receptor 1 signaling in activated lymphocytes. J. Neuroimmune Pharmacol. 2012, 7, 866–876. [Google Scholar] [CrossRef] [PubMed]
- D’Andrea, G.; Terrazzino, S.; Fortin, D.; Farruggio, A.; Rinaldi, L.; Leon, A. HPLC electrochemical detection of trace amines in human plasma and platelets and expression of mRNA transcripts of trace amine receptors in circulating leukocytes. Neurosci. Lett. 2003, 346, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Wasik, A.M.; Millan, M.J.; Scanlan, T.; Barnes, N.M.; Gordon, J. Evidence for functional trace amine associated receptor-1 in normal and malignant B cells. Leuk Res. 2012, 36, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Babusyte, A.; Kotthoff, M.; Fiedler, J.; Krautwurst, D. Biogenic amines activate blood leukocytes via trace amine-associated receptors TAAR1 and TAAR2. J. Leukoc. Biol. 2013, 93, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Borowsky, B.; Adham, N.; Jones, K.A.; Raddatz, R.; Artymyshyn, R.; Ogozalek, K.L.; Durkin, M.M.; Lakhlani, P.P.; Bonini, J.A.; Pathirana, S.; et al. Trace amines: Identification of a family of mammalian G protein-coupled receptors. Proc. Natl. Acad. Sci. USA 2001, 98, 8966–8971. [Google Scholar] [CrossRef] [PubMed]
- Liberles, S.D. Trace amine-associated receptors are olfactory receptors in vertebrates. Ann. N. Y. Acad. Sci. 2009, 1170, 168–172. [Google Scholar] [CrossRef] [PubMed]
- Carnicelli, V.; Santoro, A.; Sellari-Franceschini, S.; Berrettini, S.; Zucchi, R. Expression of trace amine-associated receptors in human nasal mucosa. Chemosens. Percept. 2010, 3, 99–107. [Google Scholar] [CrossRef]
- Johnson, M.A.; Tsai, L.; Roy, D.S.; Valenzuela, D.H.; Mosley, C.; Magklara, A.; Lomvardas, S.; Liberles, S.D.; Barnea, G. Neurons expressing trace amine-associated receptors project to discrete glomeruli and constitute an olfactory subsystem. Proc. Natl. Acad. Sci. USA 2012, 109, 13410–13415. [Google Scholar] [CrossRef] [PubMed]
- Pacifico, R.; Dewan, A.; Cawley, D.; Guo, C.; Bozza, T. An olfactory subsystem that mediates high-sensitivity detection of volatile amines. Cell Rep. 2012, 2, 76–88. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Pacifico, R.; Cawley, D.; Feinstein, P.; Bozza, T. Ultrasensitive detection of amines by a trace amine-associated receptor. J. Neurosci. 2013, 33, 3228–3239. [Google Scholar] [CrossRef] [PubMed]
- Frascarelli, S.; Ghelardoni, S.; Chiellini, G.; Galli, E.; Ronca, F.; Scanlan, T.S.; Zucchi, R. Cardioprotective effect of 3-iodothyronamine in perfused rat heart subjected to ischemia and reperfusion. Cardiovasc. Drugs Ther. 2011, 25, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Doyle, K.P.; Suchland, K.L.; Ciesielski, T.M.; Lessov, N.S.; Grandy, D.K.; Scanlan, T.S.; Stenzel-Poore, M.P. Novel thyroxine derivatives, thyronamine and 3-iodothyronamine, induce transient hypothermia and marked neuroprotection against stroke injury. Stroke 2007, 38, 2569–2576. [Google Scholar] [CrossRef] [PubMed]
- Panas, H.N.; Lynch, L.J.; Vallender, E.J.; Xie, Z.; Chen, G.L.; Lynn, S.K.; Scanlan, T.S.; Miller, G.M. Normal thermoregulatory responses to 3-iodothyronamine, trace amines and amphetamine-like psychostimulants in trace amine associated receptor 1 knockout mice. J. Neurosci. Res. 2010, 88, 1962–1969. [Google Scholar] [PubMed]
- Lindemann, L.; Meyer, C.A.; Jeanneau, K.; Bradaia, A.; Ozmen, L.; Bluethmann, H.; Bettler, B.; Wettstein, J.G.; Borroni, E.; Moreau, J.L.; et al. Trace amine-associated receptor 1 modulates dopaminergic activity. J. Pharmacol. Exp. Ther. 2008, 324, 948–956. [Google Scholar] [CrossRef] [PubMed]
- Wallrabenstein, I.; Kuklan, J.; Weber, L.; Zborala, S.; Werner, M.; Altmuller, J.; Becker, C.; Schmidt, A.; Hatt, H.; Hummel, T.; et al. Human trace amine-associated receptor TAAR5 can be activated by trimethylamine. PLoS One 2013, 8, e54950. [Google Scholar] [CrossRef] [PubMed]
- Chiellini, G.; Erba, P.; Carnicelli, V.; Manfredi, C.; Frascarelli, S.; Ghelardoni, S.; Mariani, G.; Zucchi, R. Distribution of exogenous [125I]-3-iodothyronamine in mouse in vivo: Relationship with trace amine-associated receptors. J. Endocrinol. 2012, 213, 223–230. [Google Scholar] [CrossRef] [PubMed]
- D’Andrea, G.; D’Arrigo, A.; Facchinetti, F.; del Giudice, E.; Colavito, D.; Bernardini, D.; Leon, A. Octopamine, unlike other trace amines, inhibits responses of astroglia-enriched cultures to lipopolysaccharide via a beta-adrenoreceptor-mediated mechanism. Neurosci. Lett. 2012, 517, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Regard, J.B.; Sato, I.T.; Coughlin, S.R. Anatomical profiling of G protein-coupled receptor expression. Cell 2008, 135, 561–571. [Google Scholar] [CrossRef] [PubMed]
- Ambion/Life Technologies. Carlsbad, CA, USA. Personal communication, 2014.
- Thiel, G.; Kaufmann, A.; Rossler, O.G. G-protein-coupled designer receptors—New chemical-genetic tools for signal transduction research. Biol. Chem. 2013, 394, 1615–1622. [Google Scholar] [CrossRef] [PubMed]
- Eason, M.G.; Kurose, H.; Holt, B.D.; Raymond, J.R.; Liggett, S.B. Simultaneous coupling of alpha 2-adrenergic receptors to two G-proteins with opposing effects. Subtype-selective coupling of alpha 2C10, alpha 2C4, and alpha 2C2 adrenergic receptors to Gi and Gs. J. Biol. Chem. 1992, 267, 15795–15801. [Google Scholar] [PubMed]
- Xiao, R.P.; Zhu, W.; Zheng, M.; Cao, C.; Zhang, Y.; Lakatta, E.G.; Han, Q. Subtype-specific alpha1- and beta-adrenoceptor signaling in the heart. Trends Pharmacol. Sci. 2006, 27, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.V.; Heilman, C.J.; Brann, M.R. Functional responses of cloned muscarinic receptors expressed in CHO-K1 cells. Mol. Pharmacol. 1991, 40, 242–247. [Google Scholar] [PubMed]
- Wess, J. Molecular basis of receptor/G-protein-coupling selectivity. Pharmacol. Ther. 1998, 80, 231–264. [Google Scholar] [CrossRef] [PubMed]
- Raymond, J.R. Multiple mechanisms of receptor-G protein signaling specificity. Am. J. Physiol. 1995, 269, F141–F158. [Google Scholar] [PubMed]
- Rozenfeld, R.; Devi, L.A. Exploring a role for heteromerization in GPCR signalling specificity. Biochem. J. 2011, 433, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Levoye, A.; Dam, J.; Ayoub, M.A.; Guillaume, J.L.; Jockers, R. Do orphan G-protein-coupled receptors have ligand-independent functions? New insights from receptor heterodimers. EMBO Rep. 2006, 7, 1094–1098. [Google Scholar] [CrossRef] [PubMed]
- Kuner, R.; Kohr, G.; Grunewald, S.; Eisenhardt, G.; Bach, A.; Kornau, H.C. Role of heteromer formation in GABAB receptor function. Science 1999, 283, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Ng, G.Y.; Clark, J.; Coulombe, N.; Ethier, N.; Hebert, T.E.; Sullivan, R.; Kargman, S.; Chateauneuf, A.; Tsukamoto, N.; McDonald, T.; et al. Identification of a GABAB receptor subunit, gb2, required for functional GABAB receptor activity. J. Biol. Chem. 1999, 274, 7607–7610. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.A.; Borowsky, B.; Tamm, J.A.; Craig, D.A.; Durkin, M.M.; Dai, M.; Yao, W.J.; Johnson, M.; Gunwaldsen, C.; Huang, L.Y.; et al. GABA(B) receptors function as a heteromeric assembly of the subunits GABA(B)R1 and GABA(B)R2. Nature 1998, 396, 674–679. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, H.; Matsunami, H. Synergism of accessory factors in functional expression of mammalian odorant receptors. J. Biol. Chem. 2007, 282, 15284–15293. [Google Scholar] [CrossRef] [PubMed]
- Kleinau, G.; Pratzka, J.; Nurnberg, D.; Gruters, A.; Fuhrer-Sakel, D.; Krude, H.; Kohrle, J.; Schoneberg, T.; Biebermann, H. Differential modulation of beta-adrenergic receptor signaling by trace amine-associated receptor 1 agonists. PLoS One 2011, 6, e27073. [Google Scholar] [CrossRef] [PubMed]
- Piechowski, C.L.; Rediger, A.; Lagemann, C.; Muhlhaus, J.; Muller, A.; Pratzka, J.; Tarnow, P.; Gruters, A.; Krude, H.; Kleinau, G.; et al. Inhibition of melanocortin-4 receptor dimerization by substitutions in intracellular loop 2. J. Mol. Endocrinol. 2013, 51, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Hamblin, M.W.; Metcalf, M.A.; McGuffin, R.W.; Karpells, S. Molecular cloning and functional characterization of a human 5-HT1B serotonin receptor: A homologue of the rat 5-HT1B receptor with 5-HT1D-like pharmacological specificity. Biochem. Biophys. Res. Commun. 1992, 184, 752–759. [Google Scholar] [CrossRef] [PubMed]
- Laugwitz, K.L.; Allgeier, A.; Offermanns, S.; Spicher, K.; van Sande, J.; Dumont, J.E.; Schultz, G. The human thyrotropin receptor: A heptahelical receptor capable of stimulating members of all four G protein families. Proc. Natl. Acad. Sci. USA 1996, 93, 116–120. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mühlhaus, J.; Dinter, J.; Nürnberg, D.; Rehders, M.; Depke, M.; Golchert, J.; Homuth, G.; Yi, C.-X.; Morin, S.; Köhrle, J.; et al. Analysis of Human TAAR8 and Murine Taar8b Mediated Signaling Pathways and Expression Profile. Int. J. Mol. Sci. 2014, 15, 20638-20655. https://doi.org/10.3390/ijms151120638
Mühlhaus J, Dinter J, Nürnberg D, Rehders M, Depke M, Golchert J, Homuth G, Yi C-X, Morin S, Köhrle J, et al. Analysis of Human TAAR8 and Murine Taar8b Mediated Signaling Pathways and Expression Profile. International Journal of Molecular Sciences. 2014; 15(11):20638-20655. https://doi.org/10.3390/ijms151120638
Chicago/Turabian StyleMühlhaus, Jessica, Juliane Dinter, Daniela Nürnberg, Maren Rehders, Maren Depke, Janine Golchert, Georg Homuth, Chun-Xia Yi, Silke Morin, Josef Köhrle, and et al. 2014. "Analysis of Human TAAR8 and Murine Taar8b Mediated Signaling Pathways and Expression Profile" International Journal of Molecular Sciences 15, no. 11: 20638-20655. https://doi.org/10.3390/ijms151120638
APA StyleMühlhaus, J., Dinter, J., Nürnberg, D., Rehders, M., Depke, M., Golchert, J., Homuth, G., Yi, C. -X., Morin, S., Köhrle, J., Brix, K., Tschöp, M., Kleinau, G., & Biebermann, H. (2014). Analysis of Human TAAR8 and Murine Taar8b Mediated Signaling Pathways and Expression Profile. International Journal of Molecular Sciences, 15(11), 20638-20655. https://doi.org/10.3390/ijms151120638