Molecular Actions of Ovarian Cancer G Protein-Coupled Receptor 1 Caused by Extracellular Acidification in Bone

{kind=link}
{kind=link}
Abstract
:1. Introduction
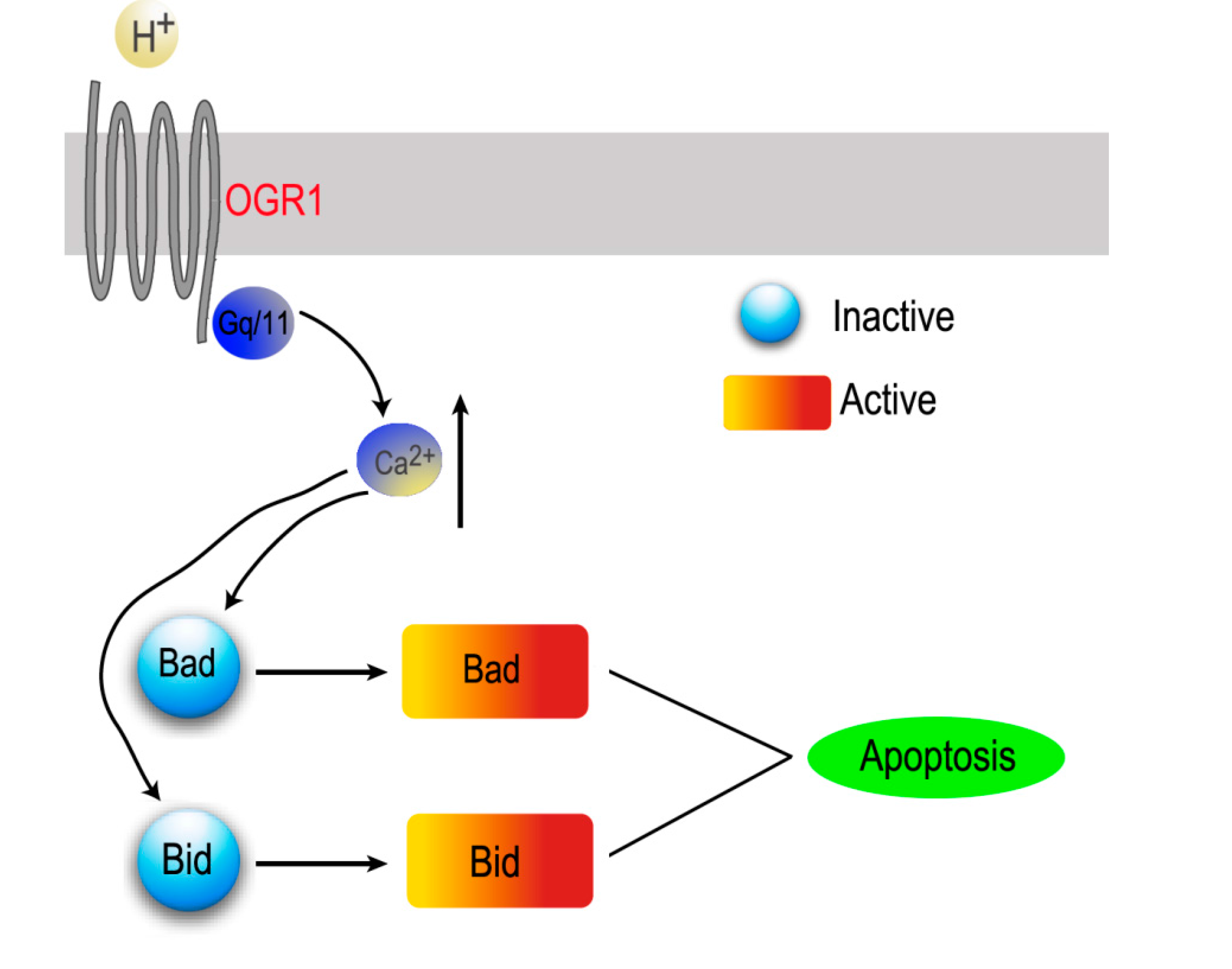
2. Proton-Sensing GPCRs

3. OGR1 and Bone
3.1. Causes of Acidosis
3.2. OGR1 and Osteoclasts
3.3. OGR1 and Osteoblasts
3.4. OGR1 and Chondrocytes

4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sgariglia, F.; Candela, M.E.; Huegel, J.; Jacenko, O.; Koyama, E.; Yamaguchi, Y.; Pacifici, M. Enomoto-Iwamoto, M. Epiphyseal abnormalities, trabecular bone loss and articular chondrocyte hypertrophy develop in the long bones of postnatal Ext1-deficient mice. Bone 2013, 57, 220–231. [Google Scholar]
- Ortega, N.; Behonick, D.; Stickens, D.; Werb, Z. How proteases regulate bone morphogenesis. Ann. N. Y. Acad. Sci. 2003, 995, 109–116. [Google Scholar]
- Li, X.; Wu, F.R.; Xu, R.S.; Hu, W.; Jiang, D.L.; Ji, C.; Chen, F.H.; Yuan, F.L. Acid-sensing ion channel 1α-mediated calcium influx regulates apoptosis of endplate chondrocytes in intervertebral discs. Expert Opin. Ther. Targets 2014, 18, 1–14. [Google Scholar]
- Saito, T.; Kawaguchi, H. HIF-2α as a possible therapeutic target of osteoarthritis. Osteoarthr. Cartil. 2010, 18, 1552–1556. [Google Scholar]
- Su, Y.S.; Ren, D.; Wang, P.C. Comparison of biomechanical properties of single- and two-segment fusion for Denis type B spinal fractures. Orthop. Surg. 2013, 5, 266–273. [Google Scholar]
- Kato, K.; Morita, I. Promotion of osteoclast differentiation and activation in spite of impeded osteoblast-lineage differentiation under acidosis: Effects of acidosis on bone metabolism. Biosci. Trends 2013, 7, 33–41. [Google Scholar]
- Spector, J.A.; Mehrara, B.J.; Greenwald, J.A.; Saadeh, P.B.; Steinbrech, D.S.; Bouletreau, P.J.; Smith, L.P.; Longaker, M.T. Osteoblast expression of vascular endothelial growth factor is modulated by the extracellular microenvironment. Am. J. Physiol. Cell Physiol. 2001, 280, C72–C80. [Google Scholar]
- Takeuchi, S.; Hirukawa, K.; Togari, A. Acidosis inhibits mineralization in human osteoblasts. Calcif. Tissue Int. 2013, 93, 233–240. [Google Scholar]
- Santhanagopal, A.; Dixon, S.J. Insulin-like growth factor I rapidly enhances acid efflux from osteoblastic cells. Am. J. Physiol. 1999, 277, E423–E432. [Google Scholar]
- Felka, T.; Schafer, R.; Schewe, B.; Benz, K.; Aicher, W.K. Hypoxia reduces the inhibitory effect of IL-1β on chondrogenic differentiation of FCS-free expanded MSC. Osteoarthr. Cartil. 2009, 17, 1368–1376. [Google Scholar]
- Solinski, H.J.; Zierler, S.; Gudermann, T.; Breit, A. Human sensory neuron-specific Mas-related G protein-coupled receptors-X1 sensitize and directly activate transient receptor potential cation channel V1 via distinct signaling pathways. J. Biol. Chem. 2012, 287, 40956–40971. [Google Scholar]
- Li, X.; Xu, R.S.; Jiang, D.L.; He, X.L.; Jin, C.; Lu, W.G.; Su, Q.; Yuan, F.L. Acid-sensing ion channel 1α is involved in acid-induced osteoclastogenesis by regulating activation of the transcription factor NFATc1. FEBS Lett. 2013, 587, 3236–3242. [Google Scholar]
- Yuan, F.L.; Wang, H.R.; Zhao, M.D.; Yuan, W.; Cao, L.; Duan, P.G.; Jiang, Y.Q.; Li, X.L.; Dong, J. Ovarian cancer G protein-coupled receptor 1 is involved in acid-induced apoptosis of endplate chondrocytes in intervertebral discs. J. Bone Miner. Res. 2014, 29, 67–77. [Google Scholar]
- Damaghi, M.; Wojtkowiak, J.W.; Gillies, R.J. pH sensing and regulation in cancer. Front. Physiol. 2013, 4, 370. [Google Scholar]
- Frick, K.K.; Krieger, N.S.; Nehrke, K.; Bushinsky, D.A. Metabolic acidosis increases intracellular calcium in bone cells through activation of the proton receptor OGR1. J. Bone Miner. Res. 2009, 24, 305–313. [Google Scholar]
- Yang, M.; Mailhot, G.; Birnbaum, M.J.; MacKay, C.A.; Mason-Savas, A.; Odgren, P.R. Expression of and role for ovarian cancer G-protein-coupled receptor 1 (OGR1) during osteoclastogenesis. J. Biol. Chem. 2006, 281, 23598–23605. [Google Scholar]
- Ludwig, M.G.; Vanek, M.; Guerini, D.; Gasser, J.A.; Jones, C.E.; Junker, U.; Hofstetter, H.; Wolf, R.M.; Seuwen, K. Proton-sensing G-protein-coupled receptors. Nature 2003, 425, 93–98. [Google Scholar]
- Meyer zu Heringdorf, D.; Jakobs, K.H. Lysophospholipid receptors: Signalling, pharmacology and regulation by lysophospholipid metabolism. Biochim. Biophys. Acta 2007, 1768, 923–940. [Google Scholar]
- Radu, C.G.; Nijagal, A.; McLaughlin, J.; Wang, L.; Witte, O.N. Differential proton sensitivity of related G protein-coupled receptors T cell death-associated gene 8 and G2A expressed in immune cells. Proc. Natl. Acad. Sci. USA 2005, 102, 1632–1637. [Google Scholar]
- Justus, C.R.; Dong, L.; Yang, L.V. Acidic tumor microenvironment and pH-sensing G protein-coupled receptors. Front. Physiol. 2013, 4, 354. [Google Scholar]
- Murakami, N.; Yokomizo, T.; Okuno, T.; Shimizu, T. G2A is a proton-sensing G-protein-coupled receptor antagonized by lysophosphatidylcholine. J. Biol. Chem. 2004, 279, 42484–42491. [Google Scholar]
- Wang, J.Q.; Kon, J.; Mogi, C.; Tobo, M.; Damirin, A.; Sato, K.; Komachi, M.; Malchinkhuu, E.; Murata, N.; Kimura, T.; et al. TDAG8 is a proton-sensing and psychosine-sensitive G-protein-coupled receptor. J. Biol. Chem. 2004, 279, 45626–45633. [Google Scholar]
- Xu, Y.; Casey, G. Identification of human OGR1, a novel G protein-coupled receptor that maps to chromosome 14. Genomics 1996, 35, 397–402. [Google Scholar]
- Okajima, F. Regulation of inflammation by extracellular acidification and proton-sensing GPCRs. Cell Signal. 2013, 25, 2263–2271. [Google Scholar]
- Im, D.S. Two ligands for a GPCR, proton vs. lysolipid. Acta Pharmacol. Sin. 2005, 26, 1435–1441. [Google Scholar]
- Arnett, T.R. Acidosis, hypoxia and bone. Arch. Biochem. Biophys. 2010, 503, 103–109. [Google Scholar]
- Arnett, T. Regulation of bone cell function by acid-base balance. Proc. Nutr. Soc. 2003, 62, 511–520. [Google Scholar]
- Pereverzev, A.; Komarova, S.V.; Korcok, J.; Armstrong, S.; Tremblay, G.B.; Dixon, S.J.; Sims, S.M. Extracellular acidification enhances osteoclast survival through an NFAT-independent, protein kinase C-dependent pathway. Bone 2008, 42, 150–161. [Google Scholar]
- Komarova, S.V.; Pereverzev, A.; Shum, J.W.; Sims, S.M.; Dixon, S.J. Convergent signaling by acidosis and receptor activator of NF-κB ligand (RANKL) on the calcium/calcineurin/NFAT pathway in osteoclasts. Proc. Natl. Acad. Sci. USA 2005, 102, 2643–2648. [Google Scholar]
- Iwai, K.; Koike, M.; Ohshima, S.; Miyatake, K.; Uchiyama, Y.; Saeki, Y.; Ishii, M. RGS18 acts as a negative regulator of osteoclastogenesis by modulating the acid-sensing OGR1/NFAT signaling pathway. J. Bone Miner. Res. 2007, 22, 1612–1620. [Google Scholar]
- Li, H.; Wang, D.; Singh, L.S.; Berk, M.; Tan, H.; Zhao, Z.; Steinmetz, R.; Kirmani, K.; Wei, G.; Xu, Y. Abnormalities in osteoclastogenesis and decreased tumorigenesis in mice deficient for ovarian cancer G protein-coupled receptor 1. PLoS One 2009, 4, e5705. [Google Scholar]
- Tomura, H.; Wang, J.Q.; Liu, J.P.; Komachi, M.; Damirin, A.; Mogi, C.; Tobo, M.; Nochi, H.; Tamoto, K.; Im, D.S.; et al. Cyclooxygenase-2 expression and prostaglandin E2 production in response to acidic pH through OGR1 in a human osteoblastic cell line. J. Bone Miner. Res. 2008, 23, 1129–1139. [Google Scholar]
- Frick, K.K.; Bushinsky, D.A. Effect of metabolic and respiratory acidosis on intracellular calcium in osteoblasts. Am. J. Physiol. Renal Physiol. 2010, 299, F418–F425. [Google Scholar]
- Hikiji, H.; Endo, D.; Horie, K.; Harayama, T.; Akahoshi, N.; Igarashi, H.; Kihara, Y.; Yanagida, K.; Takeda, J.; Koji, T.; et al. TDAG8 activation inhibits osteoclastic bone resorption. FASEB J. 2014, 28, 871–879. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, F.-L.; Zhao, M.-D.; Jiang, L.-B.; Wang, H.-R.; Cao, L.; Zhou, X.-G.; Li, X.-L.; Dong, J. Molecular Actions of Ovarian Cancer G Protein-Coupled Receptor 1 Caused by Extracellular Acidification in Bone. Int. J. Mol. Sci. 2014, 15, 22365-22373. https://doi.org/10.3390/ijms151222365
Yuan F-L, Zhao M-D, Jiang L-B, Wang H-R, Cao L, Zhou X-G, Li X-L, Dong J. Molecular Actions of Ovarian Cancer G Protein-Coupled Receptor 1 Caused by Extracellular Acidification in Bone. International Journal of Molecular Sciences. 2014; 15(12):22365-22373. https://doi.org/10.3390/ijms151222365
Chicago/Turabian StyleYuan, Feng-Lai, Ming-Dong Zhao, Li-Bo Jiang, Hui-Ren Wang, Lu Cao, Xiao-Gang Zhou, Xi-Lei Li, and Jian Dong. 2014. "Molecular Actions of Ovarian Cancer G Protein-Coupled Receptor 1 Caused by Extracellular Acidification in Bone" International Journal of Molecular Sciences 15, no. 12: 22365-22373. https://doi.org/10.3390/ijms151222365
APA StyleYuan, F. -L., Zhao, M. -D., Jiang, L. -B., Wang, H. -R., Cao, L., Zhou, X. -G., Li, X. -L., & Dong, J. (2014). Molecular Actions of Ovarian Cancer G Protein-Coupled Receptor 1 Caused by Extracellular Acidification in Bone. International Journal of Molecular Sciences, 15(12), 22365-22373. https://doi.org/10.3390/ijms151222365