SREBP-1 Has a Prognostic Role and Contributes to Invasion and Metastasis in Human Hepatocellular Carcinoma

Abstract
:1. Introduction
2. Results
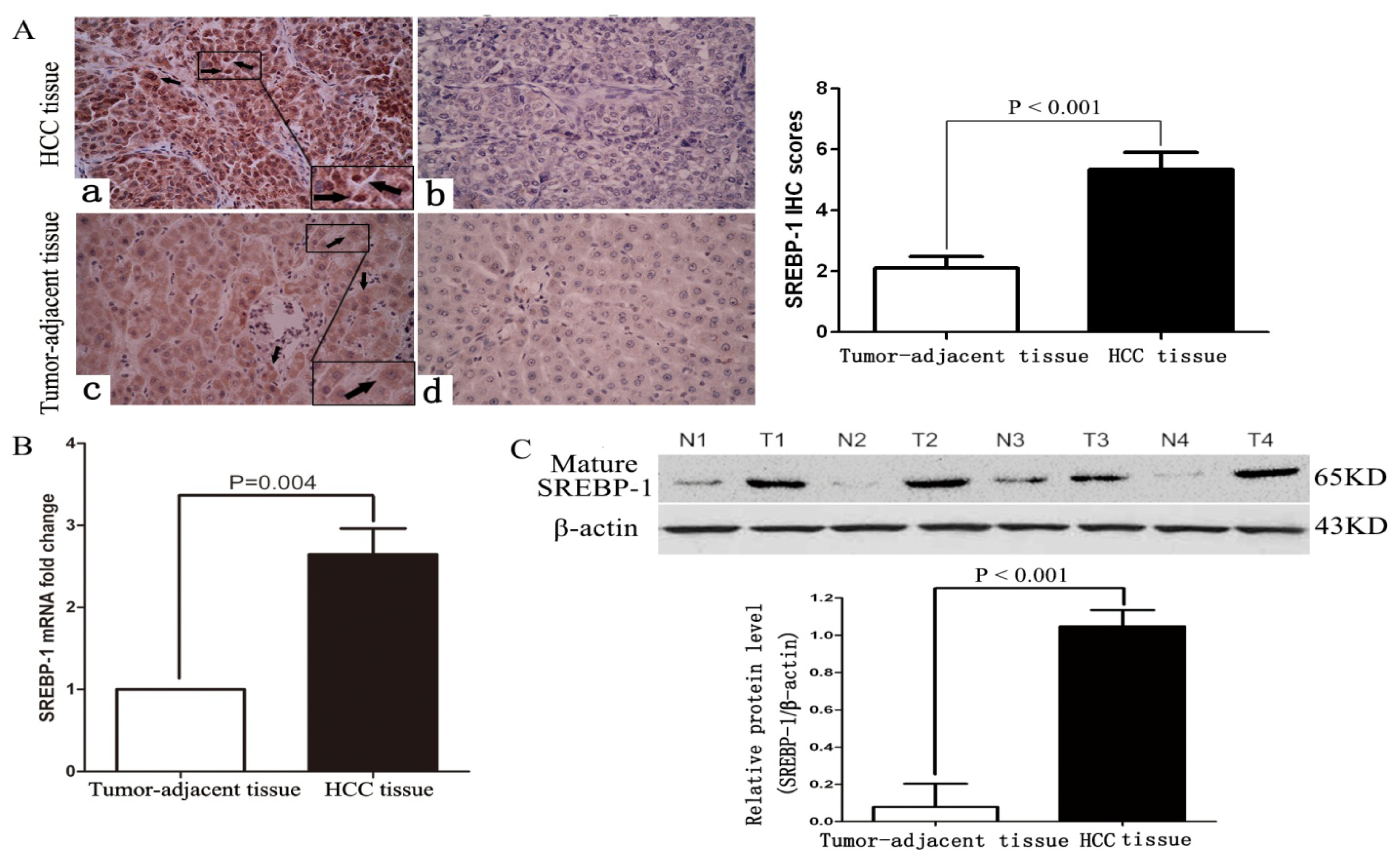
2.1. Clinical Significance of Elevated SREBP-1 Expression in HCC Tissues
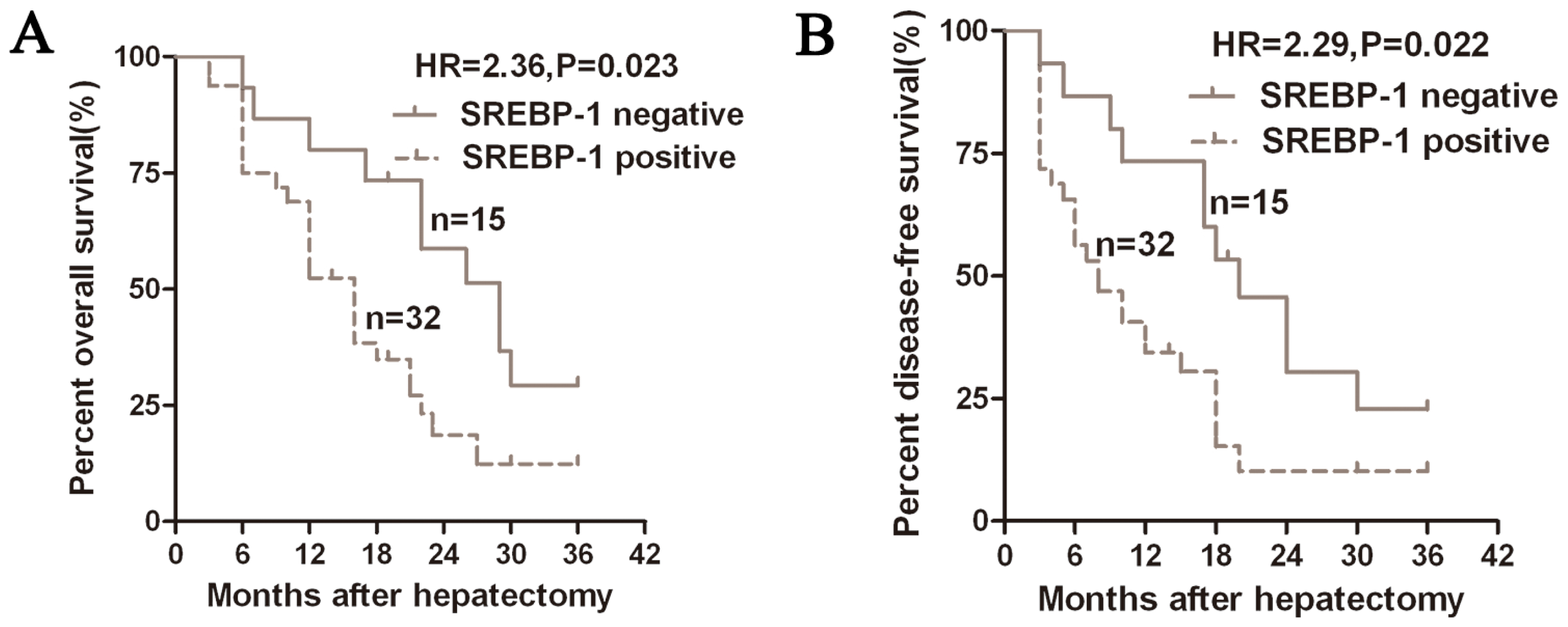
2.2. Positive Expression of SREBP-1 Correlates with a Worse 3-Year Survival of HCC Patients
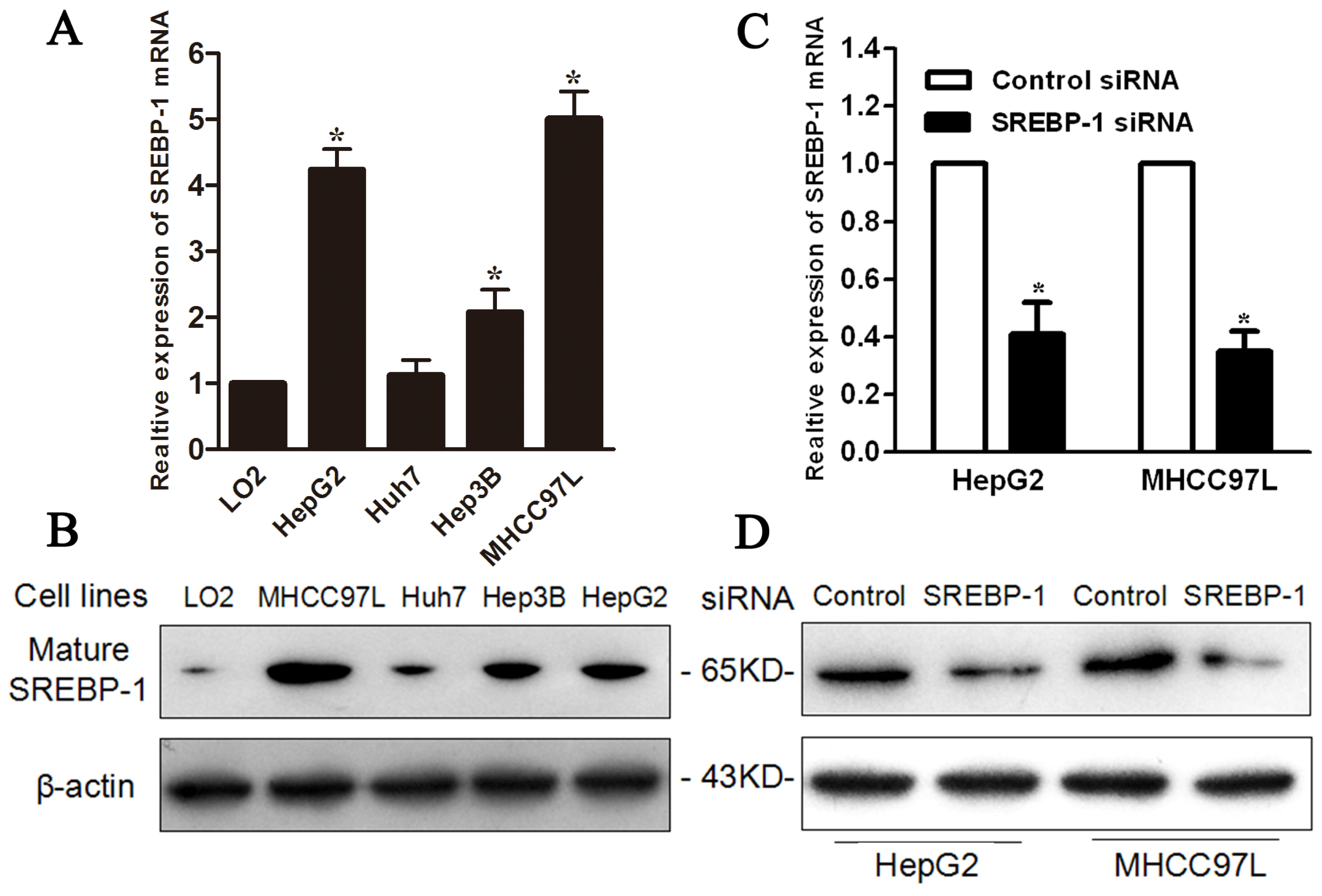
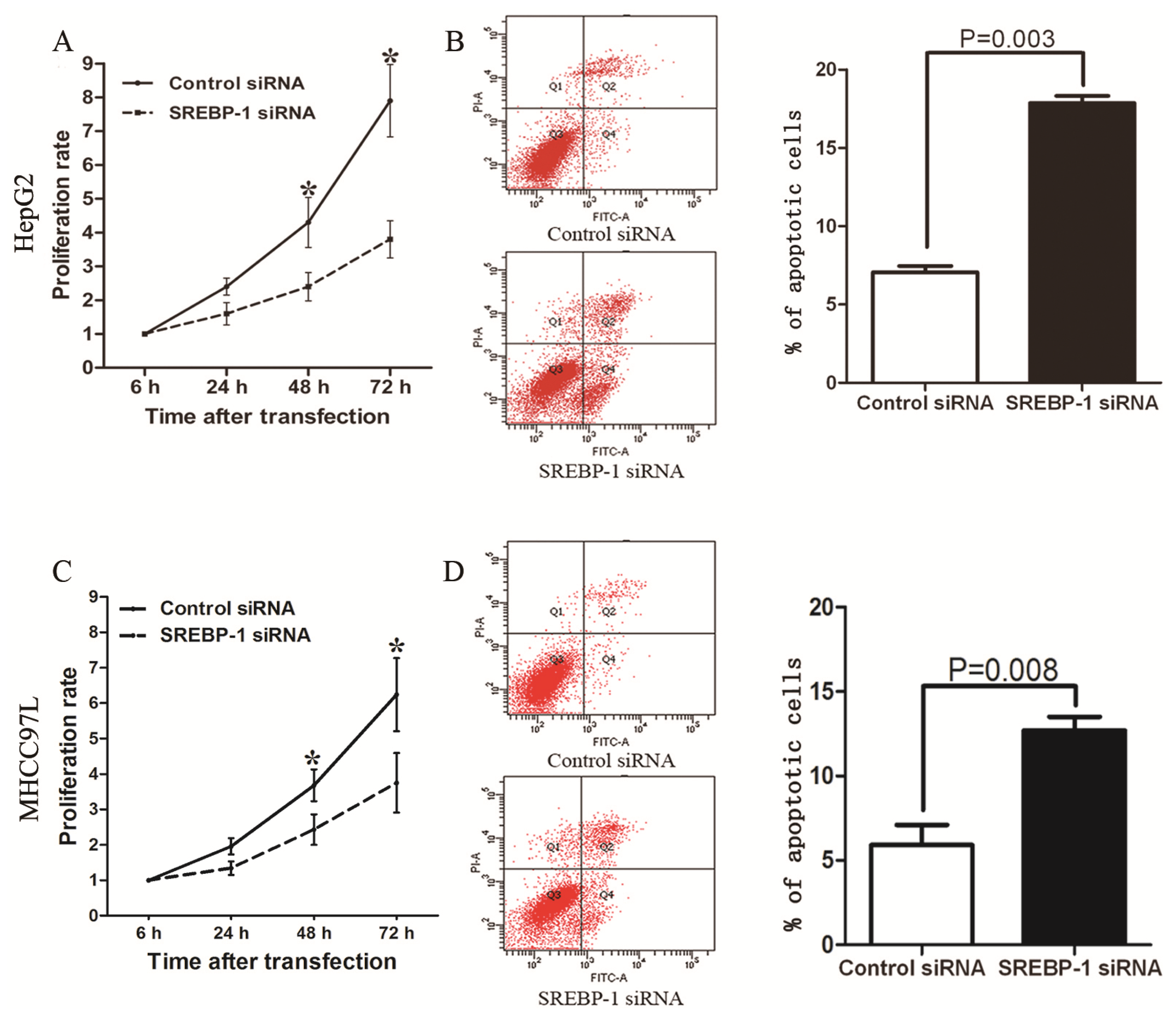
2.3. SREBP-1 Knockdown Inhibits Cell Proliferation and Induces Apoptosis in HepG2 and MHCC97L Cells
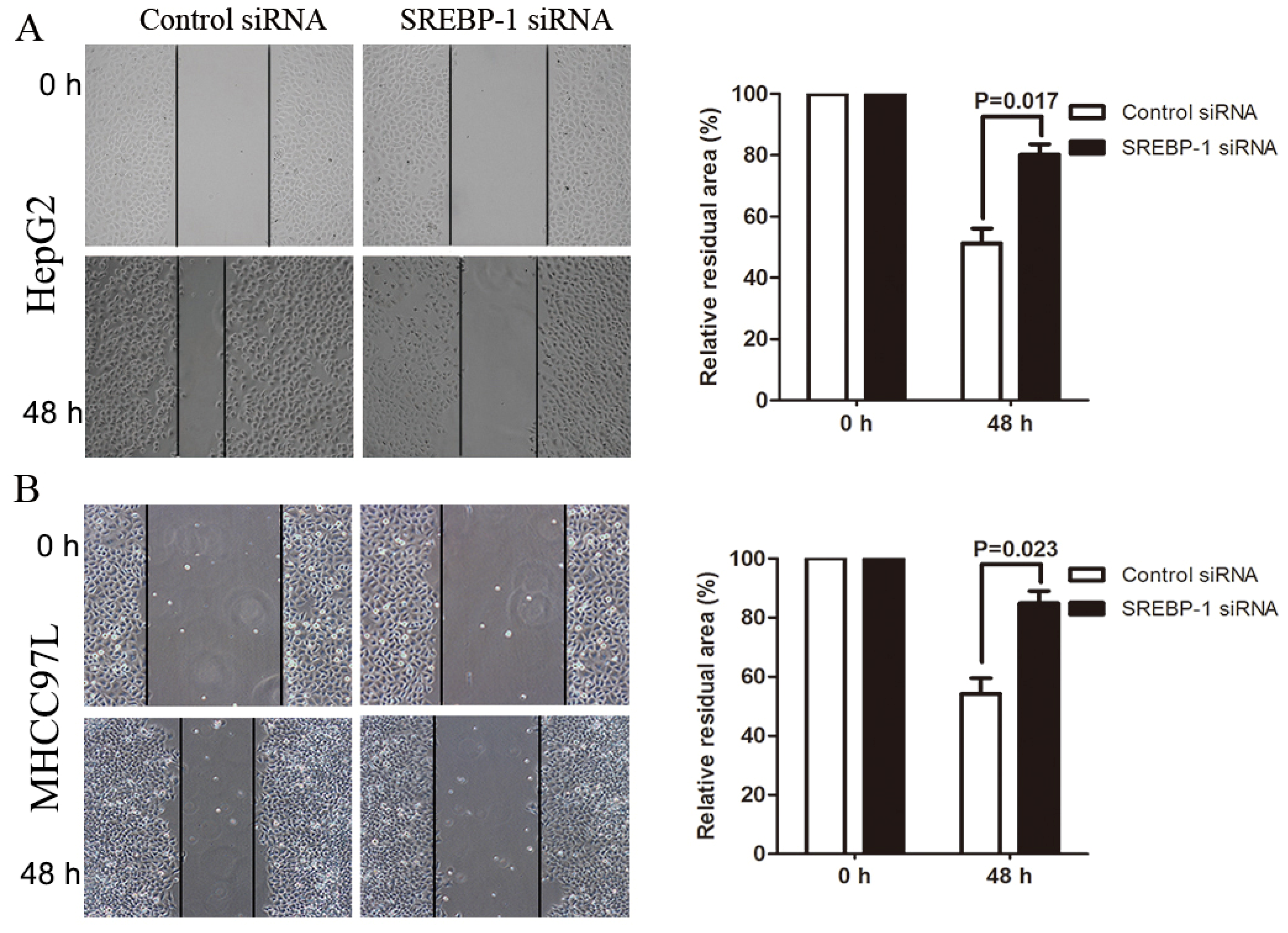
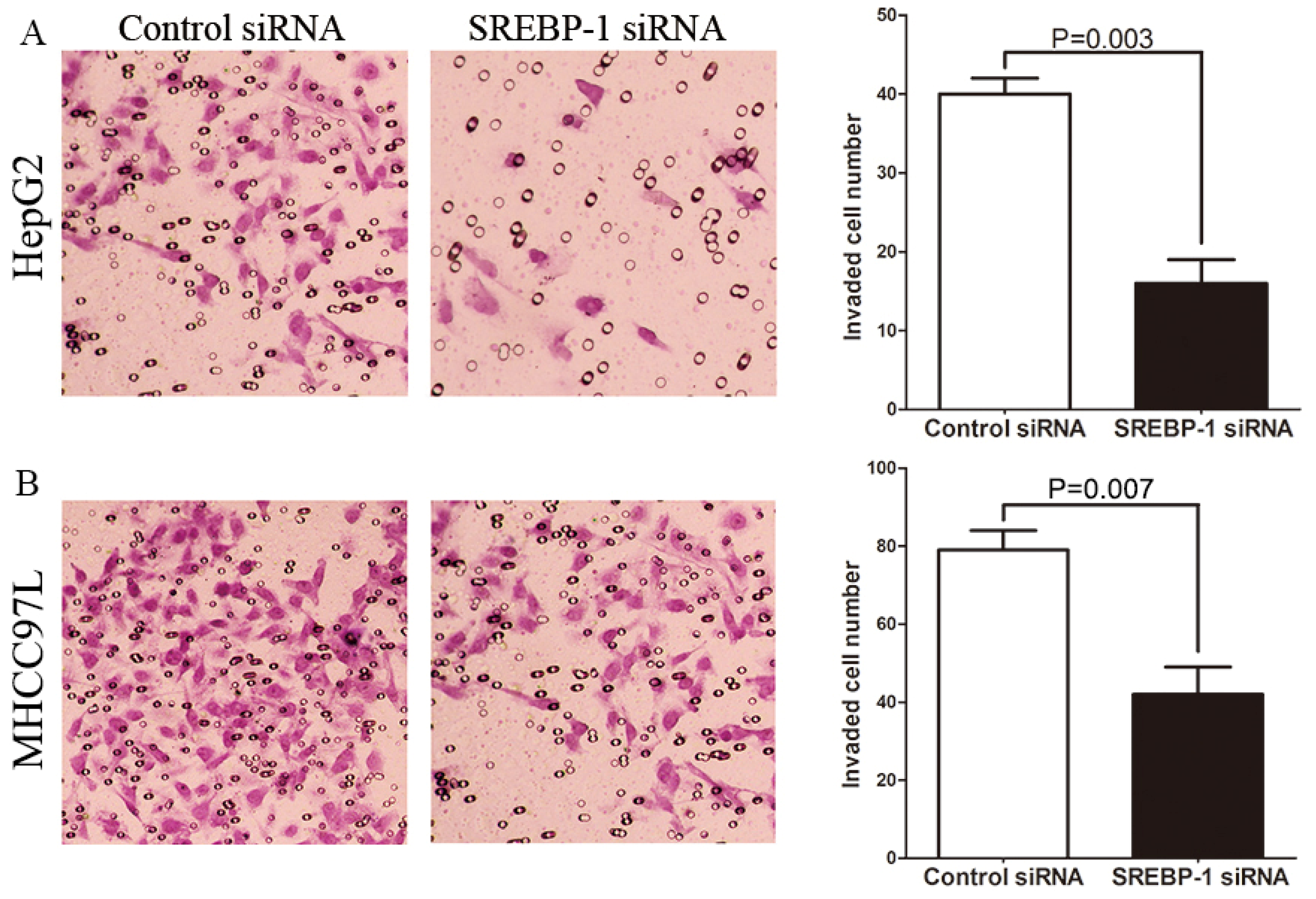
2.4. SREBP-1 Knockdown Suppresses Cell Migration and Invasion in HepG2 and MHCC97L Cells
3. Discussion
4. Experimental Section
4.1. Ethical Review
4.2. Clinical Samples and Cell Lines
4.3. Immunohistochemical Analysis
4.4. siRNA Transfection
4.5. Quantitative Real-Time Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
4.6. Immunoblot Analysis
4.7. MTT Assay
4.8. Flow Cytometry
4.9. Wound Healing Assay
4.10. Transwell Assay
4.11. Statistical Analysis
5. Conclusions
Acknowledgments
Conflicts of Interest
- Author ContributionsConceived and designed the experiments: C.L.; W.Y.; J.L.Z.; X.Z.; Y.M.Y.; K.S.T.; Q.G.L.Performed the experiments: C.L.; W.Y.; J.L.Z.; X.Z.Analyzed the data: C.L.; W.Y.; K.S.T.Contributed reagents/materials/analysis tools: X.Z.; Y.M.Y.; K.S.T.; Q.G.L.Wrote the paper: C.L.; W.Y.; J.L.Z.; X.Z.; Y.M.Y.; K.S.T.; Q.G.L.
References
- El-Serag, H.B.; Rudolph, K.L. Hepatocellular carcinoma: Epidemiology and molecular carcinogenesis. Gastroenterology 2007, 132, 2557–2576. [Google Scholar]
- Tu, K.; Zheng, X.; Zan, X.; Han, S.; Yao, Y.; Liu, Q. Evaluation of Fbxw7 expression and its correlation with the expression of cMyc, cyclin E and p53 in human hepatocellular carcinoma. Hepatol. Res 2012, 42, 904–910. [Google Scholar]
- Tu, K.; Zheng, X.; Zhou, Z.; Zan, X.; Han, S.; Yao, Y.; Liu, Q. Recombinant human adenovirus-p53 injection induced apoptosis in hepatocellular carcinoma cell lines mediated by p53-Fbxw7 pathway, which controls c-Myc and cyclin E. PLoS One 2013, 8, e68574. [Google Scholar]
- Ertle, J.; Dechêne, A.; Sowa, J.P.; Penndorf, V.; Herzer, K.; Kaiser, G.; Schlaak, J.F.; Gerken, G.; Syn, W.K.; Canbay, A. Non-alcoholic fatty liver disease progresses to hepatocellular carcinoma in the absence of apparent cirrhosis. Int. J. Cancer 2011, 128, 2436–2443. [Google Scholar]
- Nakakuki, M.; Shimano, H.; Inoue, N.; Tamura, M.; Matsuzaka, T.; Nakagawa, Y.; Yahagi, N.; Toyoshima, H.; Sato, R.; Yamada, N. A transcription factor of lipid synthesis, sterol regulatory element-binding protein (SREBP)-1a causes G (1) cell-cycle arrest after accumulation of cyclin-dependent kinase (cdk) inhibitors. FEBS J 2007, 274, 4440–4452. [Google Scholar]
- Taskinen, M.R. Diabetic dyslipidaemia: From basic research to clinical practice. Diabetologia 2003, 46, 733–749. [Google Scholar]
- Ziamajidi, N.; Khaghani, S.; Hassanzadeh, G.; Vardasbi, S.; Ahmadian, S.; Nowrouzi, A.; Ghaffari, S.M.; Abdirad, A. Amelioration by chicory seed extract of diabetes-and oleic acid-induced non-alcoholic fatty liver disease (NAFLD)/non-alcoholic steatohepatitis (NASH) via modulation of PPARα and SREBP-1. Food Chem. Toxicol 2013, 58, 198–209. [Google Scholar]
- Ruiz, R.; Jideonwo, V.; Ahn, M.; Surendran, S.; Tagliabracci, V.S.; Hou, Y.; Gamble, A.; Kerner, J.; Irimia-Dominguez, J.M.; Puchowicz, M.A.; et al. Sterol regulatory element binding Protein-1 (SREBP-1) is required to regulate glycogen synthesis and gluconeogenic gene expression in mouse liver. J. Biol. Chem 2014, 289, 5510–5517. [Google Scholar]
- Nie, L.Y.; Lu, Q.T.; Li, W.H.; Yang, N.; Dongol, S.; Zhang, X.; Jiang, J. Sterol regulatory element-binding protein 1 is required for ovarian tumor growth. Oncol. Rep 2013, 30, 1346–1354. [Google Scholar]
- Du, X.; Wang, Q.R.; Chan, E.; Merchant, M.; Liu, J.; French, D.; Ashkenazi, A.; Qing, J. FGFR3 stimulates stearoyl CoA desaturase 1 activity to promote bladder tumor growth. Cancer Res 2012, 72, 5843–5855. [Google Scholar]
- Li, W.; Tai, Y.; Zhou, J.; Gu, W.; Bai, Z.; Zhou, T.; Zhong, Z.; McCue, P.A.; Sang, N.; Ji, J.Y.; et al. Repression of endometrial tumor growth by targeting SREBP1 and lipogenesis. Cell Cycle 2012, 11, 2348–2358. [Google Scholar]
- Huang, W.C.; Li, X.; Liu, J.; Chung, L.W. Activation of androgen receptor, lipogenesis, and oxidative stress converged by SREBP-1 is responsible for regulating growth and progression of prostate cancer cells. Mol. Cancer Res 2012, 10, 133–142. [Google Scholar]
- Miyachi, K.; Sawada, Y.; Shida, Y.; Sugawara, A.; Hisatomi, H. Lipogenic gene expression profile in patients with gastric cancer. Mol. Clin. Oncol 2013, 1, 825–827. [Google Scholar]
- Mueller, K.M.; Kornfeld, J.W.; Friedbichler, K.; Blaas, L.; Egger, G.; Esterbauer, H.; Hasselblatt, P.; Schlederer, M.; Haindl, S.; Wagner, K.U.; et al. Impairment of hepatic growth hormone and glucocorticoid receptor signaling causes steatosis and hepatocellular carcinoma in mice. Hepatology 2011, 54, 1398–1409. [Google Scholar]
- Maejima, T.; Sugano, T.; Yamazaki, H.; Yoshinaka, Y.; Doi, T.; Tanabe, S.; Nishimaki-Mogami, T. Pitavastatin increases ABCA1 expression by dual mechanisms: SREBP2-driven transcriptional activation and PPARα-dependent protein stabilization but without activating LXR in rat hepatoma McARH7777 cells. J. Pharmacol. Sci 2011, 116, 107–115. [Google Scholar]
- Xu, L.; Hui, L.; Wang, S.; Gong, J.; Jin, Y.; Wang, Y.; Ji, Y.; Wu, X.; Han, Z.; Hu, G. Expression profiling suggested a regulatory role of liver-enriched transcription factors in human hepatocellular carcinoma. Cancer Res 2001, 61, 3176–3181. [Google Scholar]
- Tu, K.; Zheng, X.; Yin, G.; Zan, X.; Yao, Y.; Liu, Q. Evaluation of Fbxw7 expression and its correlation with expression of SREBP-1 in a mouse model of NAFLD. Mol. Med. Rep 2012, 6, 525–530. [Google Scholar]
- Yamashita, T.; Honda, M.; Takatori, H.; Nishino, R.; Minato, H.; Takamura, H.; Ohta, T.; Kaneko, S. Activation of lipogenic pathway correlates with cell proliferation and poor prognosis in hepatocellular carcinoma. J. Hepatol 2009, 50, 100–110. [Google Scholar]
- Kim, Y.M.; Shin, H.T.; Seo, Y.H.; Byun, H.O.; Yoon, S.H.; Lee, I.K.; Hyun, D.H.; Chung, H.Y.; Yoon, G. Sterol regulatory element-binding protein (SREBP)-1-mediated lipogenesis is involved in cell senescence. J. Biol. Chem 2010, 285, 29069–29077. [Google Scholar]
- Naito, M.; Bomsztyk, K.; Zager, R.A. Renal ischemia-induced cholesterol loading transcription factor recruitment and chromatin remodeling along the HMG-CoA reductase gene. Am. J. Pathol 2009, 174, 54–62. [Google Scholar]
- Meena, A.S.; Sharma, A.; Kumari, R.; Mohammad, N.; Vikram Singh, S.; Kumar Bhat, M. Inherent and acquired resistance to paclitaxel in hepatocellular carcinoma: molecular events involved. PLoS One 2013, 8, e61524. [Google Scholar]
- Calvisi, D.F.; Wang, C.; Ho, C.; Ladu, S.; Lee, S.A.; Mattu, S.; Destefanis, G.; Delogu, S.; Zimmermann, A.; Ericsson, J.; et al. Increased lipogenesis, induced by AKT-mTORC1-RPS6 signaling, promotes development of human hepatocellular carcinoma. Gastroenterology 2011, 140, 1071–1083. [Google Scholar]
- Cao, Z.; Fan-Minogue, H.; Bellovin, D.I.; Yevtodiyenko, A.; Arzeno, J.; Yang, Q.; Gambhir, S.S.; Felsher, D.W. MYC phosphorylation, activation, and tumorigenic potential in hepatocellular carcinoma are regulated by HMG-CoA reductase. Cancer Res 2011, 71, 2286–2297. [Google Scholar]
- Gliemroth, J.; Zulewski, H.; Arnold, H.; Terzis, A.J. Migration, proliferation, and invasion of human glioma cells following treatment with simvastatin. Neurosurg. Rev 2003, 26, 117–124. [Google Scholar]
- Fromigue, O.; Hamidouche, Z.; Vaudin, P.; Lecanda, F.; Patino, A.; Barbry, P.; Mari, B.; Marie, P.J. CYR61 downregulation reduces osteosarcoma cell invasion, migration, and metastasis. J. Bone Miner. Res 2011, 26, 1533–1542. [Google Scholar]
- Liu, Z.L.; Zhou, Y.; Luo, Q.F.; Hu, M.; Wang, G.; Huang, S.H.; Shu, Y. Inhibition of fatty acid synthase supresses osteosarcoma cell invasion and migration. Indian J. Pathol. Microbiol 2012, 55, 163–169. [Google Scholar]
- Yoshii, Y.; Furukawa, T.; Oyama, N.; Hasegawa, Y.; Kiyono, Y.; Nishii, R.; Waki, A.; Tsuji, A.B.; Sogawa, C.; Wakizaka, H.; et al. Fatty acid synthase is a key target in multiple essential tumor functions of prostate cancer:uptake of radiolabeled acetate as a predictor of the targeted therapy outcome. PLoS One 2013, 8, e64570. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinicopathologic | Features | Total No. of Patients, n = 47 | No. of Patients | p | r | |
|---|---|---|---|---|---|---|
| SREBP-1 Negative | SREBP-1 Positive | |||||
| Age (y) | <50 | 13 | 3 | 11 | 0.508 | –0.147 |
| ≥50 | 34 | 12 | 21 | |||
| Sex | Male | 30 | 11 | 19 | 0.353 | 0.135 |
| Female | 17 | 4 | 13 | |||
| HBV | Absent | 8 | 3 | 5 | 0.710 | 0.054 |
| Present | 39 | 12 | 27 | |||
| Serum AFP level (ng/mL) | <400 | 22 | 8 | 14 | 0.539 | 0.090 |
| ≥400 | 25 | 7 | 18 | |||
| Tumor size (cm) | <5 | 15 | 9 | 6 | 0.005 * | 0.413 |
| ≥5 | 32 | 6 | 26 | |||
| Cirrhosis | Absent | 9 | 5 | 4 | 0.196 | 0.247 |
| Present | 38 | 10 | 28 | |||
| Venous infiltration | Absent | 25 | 11 | 14 | 0.058 | 0.276 |
| Present | 22 | 4 | 18 | |||
| Number of nodules | 1 | 27 | 11 | 16 | 0.123 | 0.220 |
| ≥2 | 20 | 4 | 16 | |||
| Edmondson-Steiner grading | I + II | 18 | 10 | 8 | 0.006 * | 0.400 |
| III + IV | 29 | 5 | 24 | |||
| TNM tumor stage | I + II | 28 | 13 | 15 | 0.010 * | 0.378 |
| III + IV | 19 | 2 | 17 | |||
| Variables | Overall Survival | Disease-Free Survival | ||||
|---|---|---|---|---|---|---|
| HR | 95% CI | p | HR | 95% CI | p | |
| Edmondson-Steiner grade | 1.695 | 0.784–3.663 | 0.043 * | 1.646 | 0.579–4.680 | 0.035 * |
| TNM stage | 4.207 | 1.547–11.437 | 0.005 * | 7.363 | 2.134–25.400 | 0.002 * |
| SREBP-1 expression in HCC | 2.976 | 1.109–7.987 | 0.030 * | 2.327 | 1.093–4.955 | 0.029 * |
| Primers and siRNA | 5′-3′ | |
|---|---|---|
| SREBP-1 primer | F | CAGTCCAGCCTTTGAGGATA |
| R | CAAAGGATTGCAGGTCAGAC | |
| GAPDH primer | F | CAAGCTCATTTCCTGGTATGAC |
| R | CAGTGAGGGTCTCTCTCTTCCT | |
| SREBP-1 siRNA | Sense | GGAAGAGUCAGUGCCACUGTT |
| Anti-sense | CAGUGGCACUGACUCUUCCTT | |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, C.; Yang, W.; Zhang, J.; Zheng, X.; Yao, Y.; Tu, K.; Liu, Q. SREBP-1 Has a Prognostic Role and Contributes to Invasion and Metastasis in Human Hepatocellular Carcinoma. Int. J. Mol. Sci. 2014, 15, 7124-7138. https://doi.org/10.3390/ijms15057124
Li C, Yang W, Zhang J, Zheng X, Yao Y, Tu K, Liu Q. SREBP-1 Has a Prognostic Role and Contributes to Invasion and Metastasis in Human Hepatocellular Carcinoma. International Journal of Molecular Sciences. 2014; 15(5):7124-7138. https://doi.org/10.3390/ijms15057124
Chicago/Turabian StyleLi, Chao, Wei Yang, Junli Zhang, Xin Zheng, Yingmin Yao, Kangsheng Tu, and Qingguang Liu. 2014. "SREBP-1 Has a Prognostic Role and Contributes to Invasion and Metastasis in Human Hepatocellular Carcinoma" International Journal of Molecular Sciences 15, no. 5: 7124-7138. https://doi.org/10.3390/ijms15057124
APA StyleLi, C., Yang, W., Zhang, J., Zheng, X., Yao, Y., Tu, K., & Liu, Q. (2014). SREBP-1 Has a Prognostic Role and Contributes to Invasion and Metastasis in Human Hepatocellular Carcinoma. International Journal of Molecular Sciences, 15(5), 7124-7138. https://doi.org/10.3390/ijms15057124