Identification of Genes Related to Paulownia Witches’ Broom by AFLP and MSAP

Abstract
:1. Introduction
2. Results and Discussion
2.1. Morphological Changes of Paulownia Witches’ Broom (PaWB) Plantlets Treated with Methyl Methane Sulfonate (MMS)


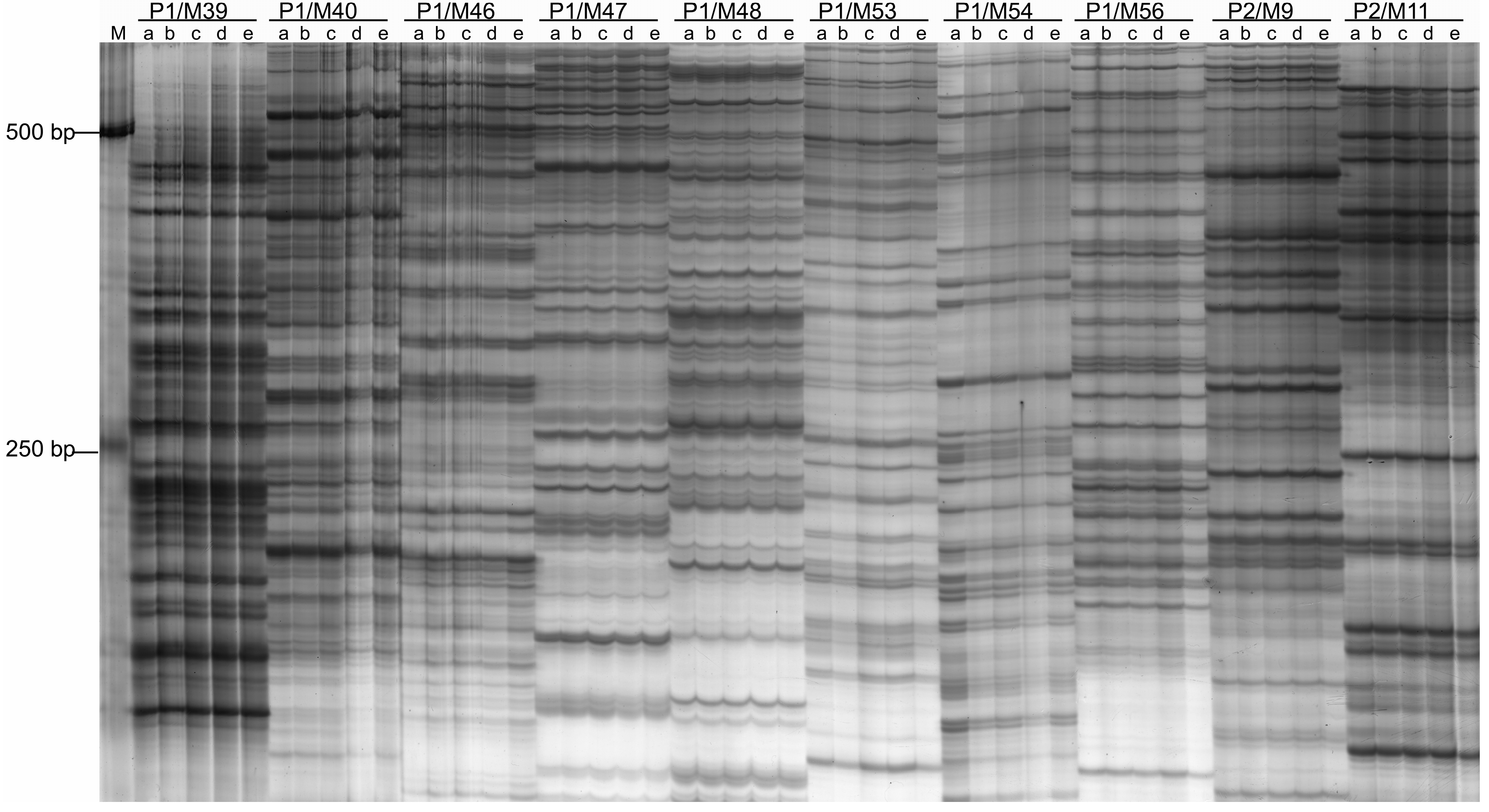
2.2. DNA Sequence Polymorphism Revealed by Amplified Fragment Length Polymorphism (AFLP)

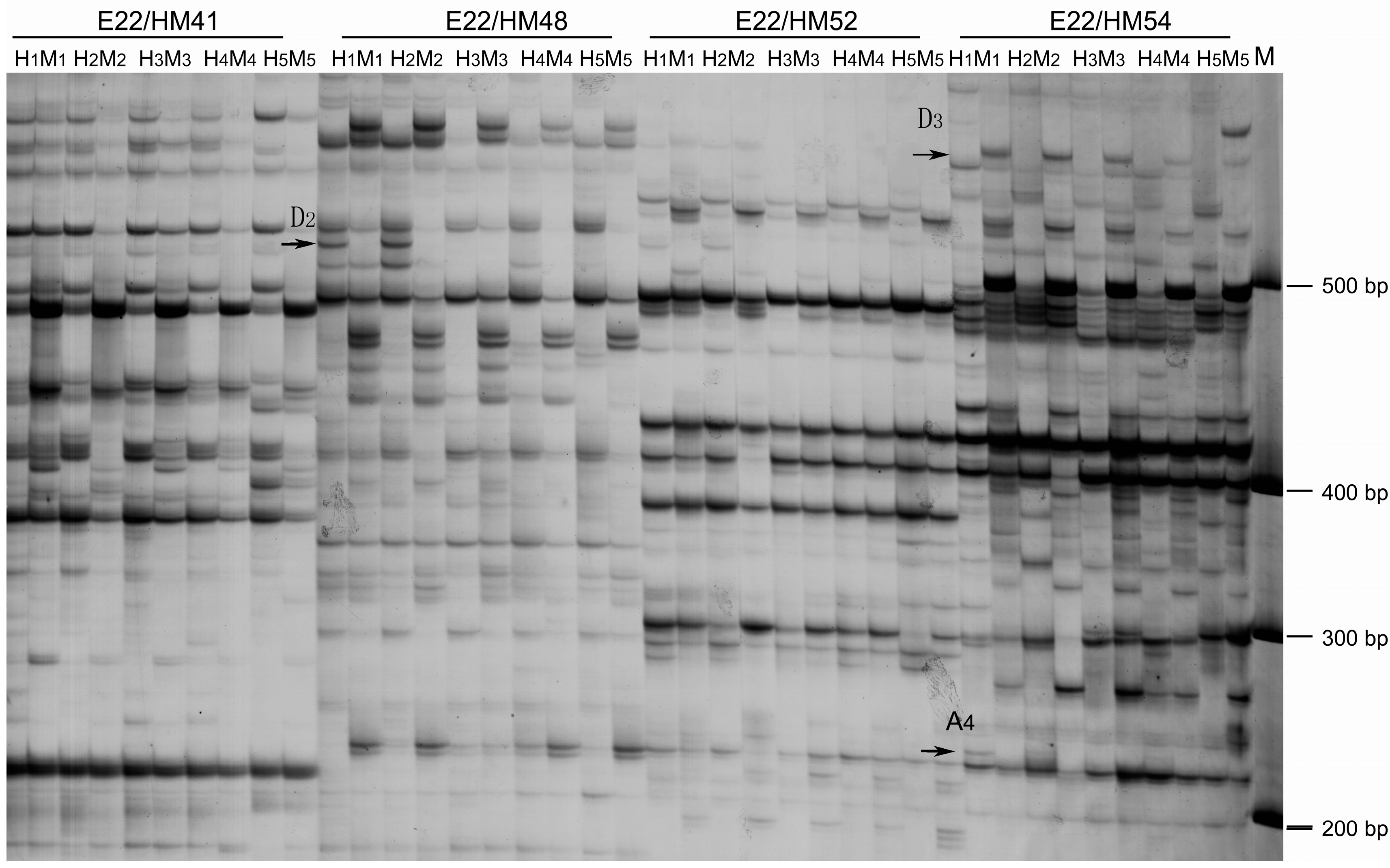
2.3. DNA Methylation Changes Revealed by Methylation-Sensitive Amplification Polymorphism (MSAP)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MMS Concentration/(mg·L–1) | Total Amplified Bands | Bands of Type I | Bands of Type II | Bands of Type III | Total Methylated Bands | Methylation Level/% |
|---|---|---|---|---|---|---|
| 0 | 2081 | 1495 | 199 | 387 | 586 | 28.15 a |
| 15 | 2173 | 1481 | 221 | 471 | 692 | 31.85 b |
| 30 | 2357 | 1559 | 267 | 531 | 798 | 33.86 c |
| 45 | 2215 | 1435 | 253 | 507 | 780 | 35.21 d |
| HP | 2193 | 1381 | 269 | 543 | 812 | 37.06 e |

| Digestion | Changes of DNA Methylation Pattern | Number of Differences Bands | Pattern | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| H1 | M1 | Hx | Mx | PaWB | Treatments/HP | 0–15 | 0–30 | 0–45 | 0–HP | |
| 1 | 1 | 0 | 1 | CCGG GGCC | CCGG GGCC | 73 | 75 | 53 | 39 | A1 |
| 1 | 1 | 1 | 0 | CCGG GGCC | CCGG CCGG GGCC GGCC | 65 | 83 | 69 | 67 | A2 |
| 0 | 1 | 0 | 0 | CCGG GGCC | CCGG GGCC | 99 | 87 | 109 | 171 | A3 |
| 1 | 0 | 0 | 0 | CCGG CCGG GGCC GGCC | CCGG GGCC | 105 | 119 | 169 | 159 | A4 |
| 0 | 1 | 1 | 1 | CCGG GGCC | CCGG GGCC | 81 | 63 | 67 | 75 | B1 |
| 1 | 0 | 1 | 1 | CCGG GGCC | CCGG CCGG GGCC GGCC | 39 | 55 | 61 | 53 | B2 |
| 0 | 0 | 0 | 1 | CCGG GGCC | CCGG GGCC | 127 | 115 | 139 | 187 | B3 |
| 0 | 0 | 1 | 1 | CCGG GGCC | CCGG GGCC | 31 | 57 | 45 | 109 | B4 |
| 0 | 1 | 1 | 0 | CCGG GGCC | CCGG CCGG GGCC GGCC | 23 | 9 | 17 | 27 | C |
| 1 | 1 | 1 | 1 | CCGG GGCC | CCGG GGCC | 1145 | 1007 | 975 | 885 | D1 |
| 1 | 0 | 1 | 0 | CCGG CCGG GGCC GGCC | CCGG CCGG GGCC GGCC | 81 | 71 | 97 | 81 | D2 |
| 0 | 1 | 0 | 1 | CCGG GGCC | CCGG GGCC | 279 | 245 | 255 | 189 | D3 |
2.4. Sequencing the MSAP Fragments
| Combination | Total Methylated Bands | Type A | Type B | Type C | Type D | ||||
|---|---|---|---|---|---|---|---|---|---|
| Bands | Ratio/% | Bands | Ratio/% | Bands | Ratio/% | Bands | Ratio/% | ||
| 0–15 | 2148 | 342 | 15.92 | 278 | 12.94 | 23 | 1.07 | 1505 | 70.07 |
| 0–30 | 1986 | 364 | 18.33 | 290 | 14.60 | 9 | 0.45 | 1323 | 66.62 |
| 0–45 | 2056 | 400 | 19.46 | 312 | 15.18 | 17 | 0.83 | 1327 | 64.54 |
| 0–HP | 2042 | 436 | 21.35 | 424 | 20.76 | 27 | 1.32 | 1155 | 56.56 |
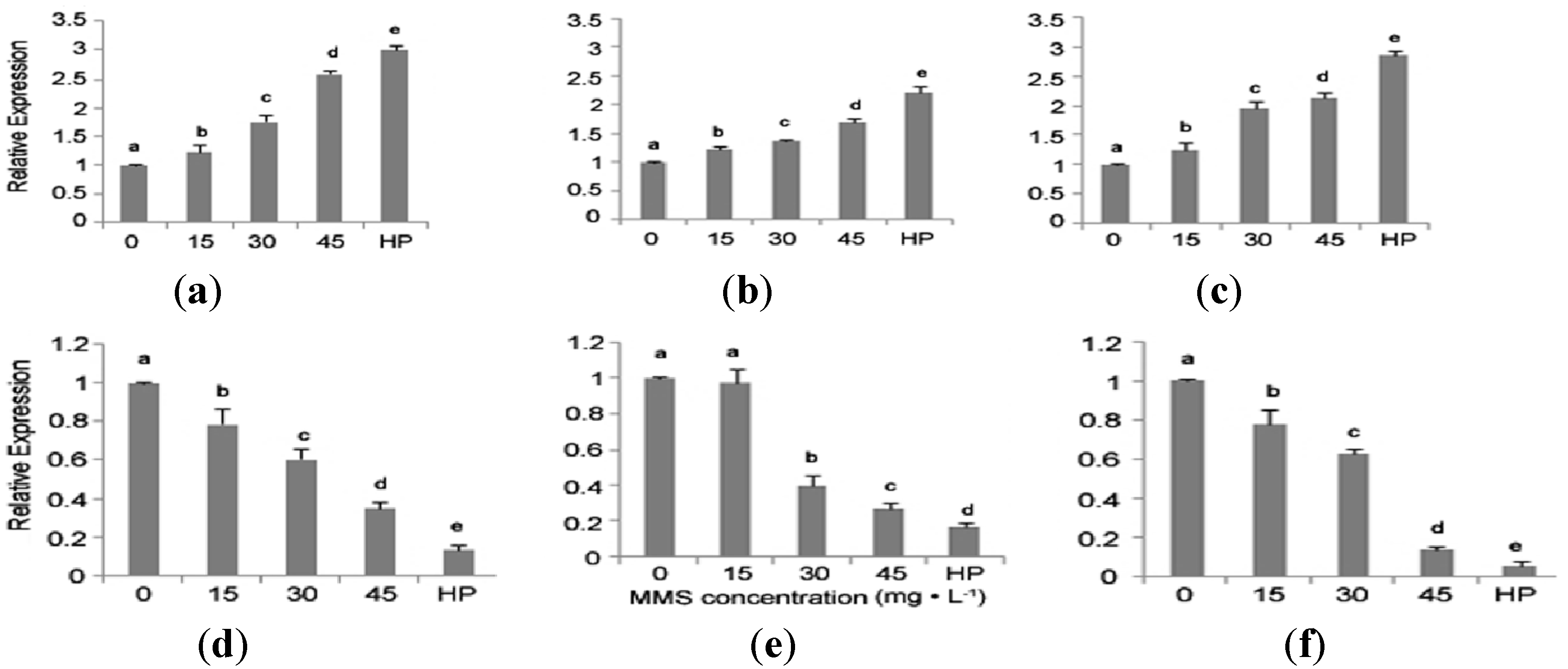
2.5. Quantitative Real-Time PCR (qRT-PCR) Analysis

2.6. Discussion
2.6.1. Relation between PaWB and DNA Methylation
2.6.2. DNA Methylation and Gene Expression Changes in PaWB
3. Experimental Section
3.1. Plantlets, MMS Treatments, and Morphological Changes
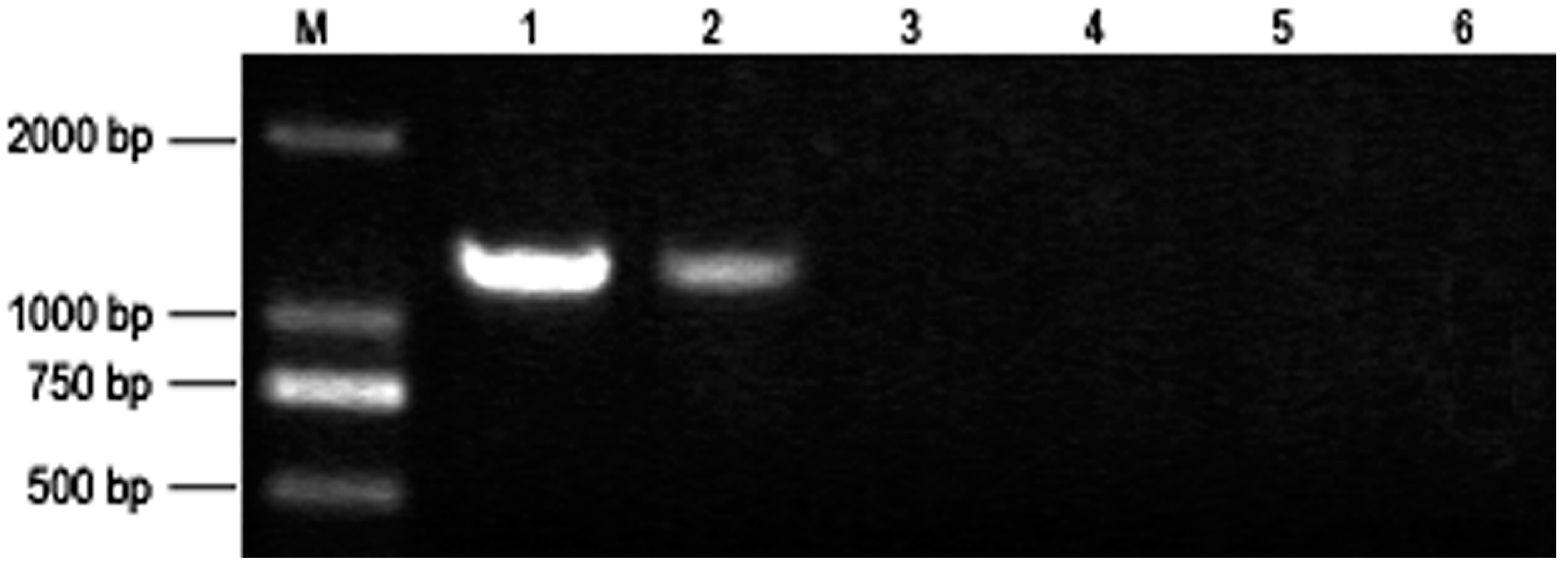
3.2. PaWB Phytoplasma Detection
3.3. AFLP and MSAP Analyses
3.4. Band Scoring and Data Analysis
3.5. Sequencing of the MSAP Fragment
3.6. Quantitative Real-Time PCR Analysis
4. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Saze, H.; Mittelsten Scheid, O.; Paszkowski, J. Maintenance of CpG methylation is essential for epigenetic inheritance during plant gametogenesis. Nat. Genet. 2003, 34, 65–69. [Google Scholar] [CrossRef]
- Cogoni, C.; Irelan, J.T.; Schumacher, M.; Schmidhauser, T.J.; Selker, E.U.; Macino, G. Transgene silencing of the al-1 gene in vegetative cells of neurospora is mediated by a cytoplasmic effector and does not depend on DNA–DNA interactions or DNA methylation. EMBO J. 1996, 15, 3153–3163. [Google Scholar]
- Tycko, B. DNA methylation in genomic imprinting. Mutat. Res.Rev. Mutat. 1997, 386, 131–140. [Google Scholar] [CrossRef]
- Zheng, X.; Chen, L.; Li, M.; Lou, Q.; Xia, H.; Wang, P.; Li, T.; Liu, H.; Luo, L. Transgenerational variations in DNA methylation induced by drought stress in two rice varieties with distinguished difference to drought resistance. PLoS One 2013, 8, e80253. [Google Scholar] [CrossRef]
- Mason, G.; Noris, E.; Lanteri, S.; Acquadro, A.; Accotto, G.P.; Portis, E. Potentiality of methylation-sensitive amplification polymorphism (MSAP) in identifying genes involved in tomato response to tomato yellow leaf curl sardinia virus. Plant Mol. Biol. Rep. 2008, 26, 156–173. [Google Scholar] [CrossRef]
- Da, K.; Nowak, J.; Flinn, B. Potato cytosine methylation and gene expression changes induced by a beneficial bacterial endophyte, Burkholderia Phytofirmans strain PsJN. Plant Physiol. Biochem. 2012, 50, 24–34. [Google Scholar] [CrossRef]
- Olson, J.R.; Carpenter, S.B. Specific gravity, fiber length, and extractive content of young paulownia. Wood Fiber Sci. 1985, 17, 428–438. [Google Scholar]
- López, F.; Pérez, A.; Zamudio, M.A.; de Alva, H.E.; García, J.C. Paulownia as raw material for solid biofuel and cellulose pulp. Biomass Bioenerg. 2012, 45, 77–86. [Google Scholar] [CrossRef]
- Krikorian, A.D. Paulownia in China: Cultivation and utilization. Econ. Bot. 1988, 42, 451. [Google Scholar] [CrossRef]
- Stanković, D.; Igić, R.; Šijačić-Nikolić, M.; Vilotić, D.; Pajević, S. Contents of the heavy metals nickel and lead in leaves of Paulownia elongata SY Hu and Paulownia fortunei Hems. Arch. Biol. Sci. 2009, 61, 827–834. (In Serbia) [Google Scholar] [CrossRef]
- Du, T.; Wang, Y.; Hu, Q.X.; Chen, J.; Liu, S.; Huang, W.J.; Lin, M.L. Transgenic paulownia expressing shiva-1 gene has increased resistance to paulownia witches’ broom disease. J. Integr. Plant Biol. 2005, 47, 1500–1506. [Google Scholar] [CrossRef]
- Yue, H.N.; Wu, Y.F.; Shi, Y.Z.; Wu, K.K.; Li, Y.R. First report of paulownia witches’-broom phytoplasma in China. Plant Dis. 2008, 92, 1134–1134. [Google Scholar]
- Mou, H.Q.; Lu, J.; Zhu, S.F.; Lin, C.L.; Tian, G.Z.; Xu, X.; Zhao, W.J. Transcriptomic analysis of paulownia infected by paulownia witches’-broom phytoplasma. PLoS One 2013, 8, e77217. [Google Scholar] [CrossRef]
- Liu, R.; Dong, Y.; Fan, G.; Zhao, Z.; Deng, M.; Cao, X.; Niu, S. Discovery of genes related to witches’ broom disease in Paulownia tomentosa × Paulownia fortunei by a de novo assembled transcriptome. PLoS One 2013, 8, e80238. [Google Scholar] [CrossRef]
- Cao, X.B.; Fan, G.Q.; Zhai, X.Q. Morphological changes of the witches’ broom seedlings of Paulownia tomentosa treated with methyl methane sulphonate and SSR analysis. Acta Phytopathol. Sin. 2012, 42, 214–218. (In Chinese) [Google Scholar]
- Zhai, X.Q.; Cao, X.B.; Fan, G.Q. Growth of paulownia witches’ broom seedlings treated with methyl methane sulphonate and SSR analysis. Sci. Silv. Sin. 2010, 46, 176–181. (In Chinese) [Google Scholar]
- Bird, A. DNA methylation patterns and epigenetic memory. Gene. Dev. 2002, 16, 6–21. [Google Scholar] [CrossRef]
- Li, M.; Zhai, X.Q.; Fan, G.Q.; Zhang, B.L.; Liu, F. Effect of oxytetracycline on the morphology of seedlings with witches’ broom and DNA methylation level of Paulownia tomentosa × Paulownia fortunei. Sci. Silv. Sin. 2008, 44, 152–156. (In Chinese) [Google Scholar]
- Xiong, L.Z.; Xu, C.G.; Saghai Maroof, M.A.; Zhang, Q. Patterns of cytosine methylation in an elite rice hybrid and its parental lines, detected by a methylation-sensitive amplification polymorphism technique. Mol. Gen. Genet. 1999, 261, 439–446. [Google Scholar] [CrossRef]
- Richards, E.J. DNA methylation and plant development. Trends Genet. 1997, 13, 319–323. [Google Scholar] [CrossRef]
- Finnegan, E.J.; Peacock, W.J.; Dennis, E.S. Reduced DNA methylation in Arabidopsis thaliana results in abnormal plant development. Proc. Natl. Acad. Sci. USA 1996, 93, 8449–8454. [Google Scholar]
- Ahmad, J.; Garcion, C.; Teyssier, E.; Hernould, M.; Gallusci, P.; Pracros, P.; Renaudin, J.; Eveillard, S. Effects of stolbur phytoplasma infection on DNA methylation processes in tomato plants. Plant Pathol. 2013, 62, 205–216. [Google Scholar] [CrossRef]
- Nambara, E.; Marion-Poll, A. Abscisic acid biosynthesis and catabolism. Annu. Rev. Plant. Biol. 2005, 56, 165–185. [Google Scholar] [CrossRef]
- Arney, S.E.; Mitchell, D.L. The effect of abscisic acid on stem elongation and correlative inhibition. New Phytol. 1969, 68, 1001–1015. [Google Scholar] [CrossRef]
- Raghavendra, A.S.; Gonugunta, V.K.; Christmann, A.; Grill, E. ABA perception and signalling. Trends Plant Sci. 2010, 15, 395–401. [Google Scholar] [CrossRef]
- Park, M.Y.; Kim, S.A.; Lee, S.J.; Kim, S.Y. ATHB17 is a positive regulator of abscisic acid response during early seedling growth. Mol. Cell 2013, 35, 125–133. [Google Scholar] [CrossRef]
- Kayyali, U.S.; Donaldson, C.; Huang, H.; Abdelnour, R.; Hassoun, P.M. Phosphorylation of xanthine dehydrogenase/oxidase in hypoxia. J. Biol. Chem. 2001, 276, 14359–14365. [Google Scholar]
- Ji, X.; Gai, Y.; Zheng, C.; Mu, Z. Comparative proteomic analysis provides new insights into mulberry dwarf responses in mulberry (Morus alba L.). Proteomics 2009, 9, 5328–5339. [Google Scholar] [CrossRef]
- Kwak, J.M.; Mori, I.C.; Pei, Z.M.; Leonhardt, N.; Torres, M.A.; Dangl, J.L.; Bloom, R.E.; Bodde, S.; Jones, J.D.; Schroeder, J.I. NADPH oxidase AtrbohD and AtrbohF genes function in ROS-dependent ABA signaling in Arabidopsis. EMBO J. 2003, 22, 2623–2633. [Google Scholar] [CrossRef]
- Kim, S.H.; Hong, J.K.; Lee, S.C.; Sohn, K.H.; Jung, H.W.; Hwang, B.K. CAZFP1, Cys2/His2-type zinc-finger transcription factor gene functions as a pathogen-induced early-defense gene in Capsicum annuum. Plant Mol. Biol. 2004, 55, 883–904. [Google Scholar] [CrossRef]
- Nakai, Y.; Nakahira, Y.; Sumida, H.; Takebayashi, K.; Nagasawa, Y.; Yamasaki, K.; Akiyama, M.; Ohme-Takagi, M.; Fujiwara, S.; Shiina, T.; et al. Vascular plant one-zinc-finger protein 1/2 transcription factors regulate abiotic and biotic stress responses in Arabidopsis. Plant J. 2013, 73, 761–775. [Google Scholar] [CrossRef]
- Park, H.C.; Kim, M.L.; Kim, H.S.; Park, J.H.; Jung, M.S.; Shen, M.; Kang, C.H.; Kim, M.C.; Lee, S.Y.; Cho, M.J.; et al. Specificity of DNA sequences recognized by the zinc-finger homeodomain protein, GmZF-HD1 in soybean. Phytochemistry 2010, 71, 1832–1838. [Google Scholar] [CrossRef]
- Hogenhout, S.A.; Oshima, K.; Ammar El, D.; Kakizawa, S.; Kingdom, H.N.; Namba, S. Phytoplasmas: Bacteria that manipulate plants and insects. Mol. Plant Pathol. 2008, 9, 403–423. [Google Scholar] [CrossRef]
- Kube, M.; Mitrovic, J.; Duduk, B.; Rabus, R.; Seemüller, E. Current view on phytoplasma genomes and encoded metabolism. Sci. World J. 2012, 2012, 25. [Google Scholar]
- Curnow, A.W.; Tumbula, D.L.; Pelaschier, J.T.; Min, B.; Soll, D. Glutamyl-tRNAGln amidotransferase in Deinococcus radiodurans may be confined to asparagine biosynthesis. Proc. Natl. Acad. Sci. USA 1998, 95, 12838–12843. [Google Scholar] [CrossRef]
- Melnick, R.L.; Marelli, J.P.; Sicher, R.C.; Strem, M.D.; Bailey, B.A. The interaction of Theobroma cacao and Moniliophthora perniciosa, the causal agent of witches' broom disease, during parthenocarpy. Tree Genet. Genomes 2012, 8, 1261–1279. [Google Scholar] [CrossRef]
- Talbot, N.J.; McCafferty, H.R.K.; Ma, M.; Moore, K.; Hamer, J.E. Nitrogen starvation of the rice blast fungus Magnaporthe grisea may act as an environmental cue for disease symptom expression. Physiol. Mol. Plant Pathol. 1997, 50, 179–195. [Google Scholar] [CrossRef]
- Snoeijers, S.S.; Pérez-García, A.; Joosten, M.H.; de Wit, P.J. The effect of nitrogen on disease development and gene expression in bacterial and fungal plant pathogens. Eur. J. Plant. Pathol. 2000, 106, 493–506. [Google Scholar] [CrossRef]
- Dubiel, W.; Ferrell, K.; Rechsteiner, M. Subunits of the regulatory complex of the 26S protease. Mol. Biol. Rep. 1995, 21, 27–34. [Google Scholar] [CrossRef]
- Ferrell, K.; Wilkinson, C.R.; Dubiel, W.; Gordon, C. Regulatory subunit interactions of the 26S proteasome, a complex problem. Trends Biochem. Sci. 2000, 25, 83–88. [Google Scholar] [CrossRef]
- Block, A.; Li, G.; Fu, Z.Q.; Alfano, J. Phytopathogen type III effector weaponry and their plant targets. Curr. Opin. Plant Biol. 2008, 11, 1–8. [Google Scholar] [CrossRef]
- Dielen, A.S.; Badaoui, S.; Candresse, T.; German-Retana, S. The ubiquitin/26S proteasome system in plant-pathogen interactions: A never-ending hide-and-seek game. Mol. Plant Pathol. 2010, 11, 293–308. [Google Scholar] [CrossRef]
- Ellis, C.; Turner, J.G.; Devoto, A. Protein complexes mediate signalling in plant responses to hormones, light, sucrose and pathogens. Plant Mol. Biol. 2002, 50, 971–980. [Google Scholar] [CrossRef]
- Manzano, C.; Abraham, Z.; Lopez-Torrejon, G.; del Pozo, J.C. Identification of ubiquitinated proteins in Arabidopsis. Plant Mol. Biol. 2008, 68, 145–158. [Google Scholar] [CrossRef]
- Bostick, M.; Lochhead, S.R.; Honda, A.; Palmer, S.; Callis, J. Related to ubiquitin 1 and 2 are redundant and essential and regulate vegetative growth, auxin signaling, and ethylene production in Arabidopsis. Plant Cell 2004, 16, 2418–2432. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plantarum 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Fan, G.Q.; Zhang, S.; Zhai, X.Q.; Liu, F.; Dong, Z.Q. Effects of antibiotics on the paulownia witches’ broom phytoplasmas and pathogenic protein related to witches' broom symptom. Sci. Silv. Sin. 2007, 43, 138–142. (In Chinese) [Google Scholar]
- Zhang, Y.Z.; Cao, X.B.; Zhai, X.Q.; Fan, G.Q. Study on DNA extraction of AFLP reaction system for Paulownia plants. J. Henan Agric. Univ. 2009, 43, 610–614. (In Chinese) [Google Scholar]
- Lee, I.M.; Hammond, R.W.; Davis, R.E.; Gundersen, D.E. Universal amplification and analysis of pathogen 16s rDNA for classification and identification of mycoplasmalike organisms. Phytopathology 1993, 83, 834–842. [Google Scholar] [CrossRef]
- Cao, X.B.; He, J.; Zhai, X.Q.; Fan, G.Q. Establishment of Paulownia AFLP reaction system and its primer selection. J. Henan Agric. Univ. 2010, 44, 145–150. (In Chinese) [Google Scholar]
- Cao, X.B.; Zhao, G.L.; Fan, G.Q. Establishment of Paulownia MSAP reaction systems and primers screening. J. Henan Agric. Univ. 2012, 46, 535–541. (In Chinese) [Google Scholar]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; Mcginnis, S.; Madden, T.L. NCBI BL-AST: A better web interface. Nucleic Acids Res. 2008, 36, W5–W9. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cao, X.; Fan, G.; Deng, M.; Zhao, Z.; Dong, Y. Identification of Genes Related to Paulownia Witches’ Broom by AFLP and MSAP. Int. J. Mol. Sci. 2014, 15, 14669-14683. https://doi.org/10.3390/ijms150814669
Cao X, Fan G, Deng M, Zhao Z, Dong Y. Identification of Genes Related to Paulownia Witches’ Broom by AFLP and MSAP. International Journal of Molecular Sciences. 2014; 15(8):14669-14683. https://doi.org/10.3390/ijms150814669
Chicago/Turabian StyleCao, Xibing, Guoqiang Fan, Minjie Deng, Zhenli Zhao, and Yanpeng Dong. 2014. "Identification of Genes Related to Paulownia Witches’ Broom by AFLP and MSAP" International Journal of Molecular Sciences 15, no. 8: 14669-14683. https://doi.org/10.3390/ijms150814669
APA StyleCao, X., Fan, G., Deng, M., Zhao, Z., & Dong, Y. (2014). Identification of Genes Related to Paulownia Witches’ Broom by AFLP and MSAP. International Journal of Molecular Sciences, 15(8), 14669-14683. https://doi.org/10.3390/ijms150814669