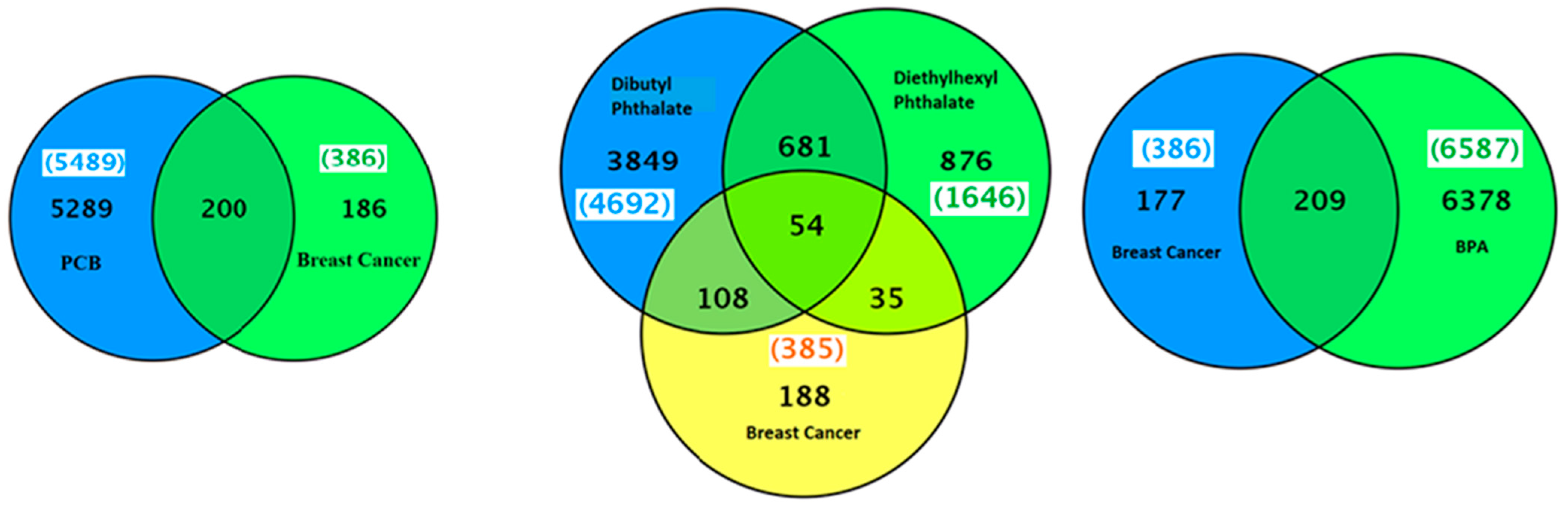
2.3. PCBs–Breast Cancer–Gene Association
The CTD search revealed that besides PCBs, the five most common PCB congeners studied for gene interaction were 3,4,3′,4′-tetrachlorobiphenyl (77), 2′,3,3′,4′,5-pentachloro-4-hydroxybiphenyl (4′-OH-PCB-86), 3,4,5,3′,4′-pentachlorobiphenyl (126), 2,3,3′,4,4′,5-hexachlorobiphenyl (153), and 2,2′,3,4,4′,5,5′-heptachlorobiphenyl (180) (
Table 2). There were 5289 genes related to PCB family of chemicals and 386 genes related to breast cancer (
Figure 3). The common genes between PCBs and breast cancer were 200. The top interacting genes with PCBs as a chemical class were
CYP1A1,
AHR,
CYP1A2,
AR,
CYP1A,
CYP1B1,
VCAM1,
MAPK1,
MAPK3, and
PTGS2. The top interacting genes with PCBs in breast neoplasms were
AR,
CYP1A1,
CYP1B1,
ESR1,
ESR2,
PTGS2, and
RAF1. Out of a total 200 genes interactions observed with individual PCBs, the interaction of genes
AR,
BAX,
CYP1A1,
CYP1B1,
KDR,
PARP1,
PTGS2, and
RAF1 was common with tetrachloride, pentachloride, and hexachloride biphenyls in beast neoplasms (
Table 2).
CYP1A1,
AHR,
AR,
CYP1A,
CYP1B1 and
PTGS2 genes are common in both PCB-gene and PCB-gene-breast cancer groups. Interactions among these genes are shown in
Figure S1. Enrichment pathway analysis revealed that these genes are part of: (1) pathways in cancer (KEGG: 05200); (2) signal transduction (REACT: 111102); (3) mTOR signaling pathway (KEGG: 04150); (4) focal adhesion (KEGG: 04510); (5) VEGF signaling pathway (KEGG: 04370); and (6) ErbB signaling pathway (
Table 3).
Figure 3.
A Venn diagram of list of genes common between breast neoplasms and PCBs, phthalates or bisphenol A.
Figure 3.
A Venn diagram of list of genes common between breast neoplasms and PCBs, phthalates or bisphenol A.
2.5. Association between Endometriosis and Exposure to PCB, Phthalates or BPA
We identified 11 epidemiologic studies related to PCB, phthalate, or BPA exposure and endometriosis. Eight of the studies assessed the relationship between PCB exposure and endometriosis [
38,
39,
40,
41,
42,
43,
44], two studies assessed the relationship between phthalate exposure and endometriosis [
29,
45], one study assessed the relationship between BPA exposure and endometriosis [
46], and one study assessed the relationship between phthalate and BPA exposure and endometriosis [
47]. Of these studies, eight were case-control studies, one was a cross-sectional study and two were cohort studies. In all of the studies, endometriosis cases were confirmed with a laparoscopic examination and/or biopsy and in nine of the eleven studies controls were also confirmed to be disease free through laparoscopic examination. Controls in the remaining two studies were randomly selected from a list of Group Health Enrollees that were known to not have endometriosis.
All eight of the studies that addressed the relationship between PCB exposure and endometriosis presented individual congener results as well as a measure of total PCBs, the sum of individual congeners. To summarize the main results between PCB exposure and endometriosis, lipid adjusted arithmetic means or geometric means of total PCB exposure were provided for four studies [
34,
38,
41,
46], median TEQ values (pg TEQ/g lipid) were provided in two studies [
40,
43], and median wet weight serum PCB concentrations were calculated in one study [
42]. Furthermore, all studies estimated the risk of endometriosis using adjusted logistic regression models with OR and 95% confidence intervals, with the majority of the studies using tertiles or quartiles to compare highest
versus lowest exposure categories.
Only three of the eight PCB case-control studies found associations between exposure to total PCBs and risk of endometriosis [
34,
36,
41]. Louis
et al. [
36] measured total PCBs (
n = 62), the sum of estrogenic PCBs (
n = 12), and the sum of anti-estrogenic PCBs (
n = 4) in a cohort study of 84 women undergoing laparoscopy (32 endometriosis cases, 52 controls). They found a significant increased risk of endometriosis for the sum of anti-estrogenic PCBs for women in the third tertile (OR = 3.77, 95% CI 1.12–12.68), however, the risk remained elevated but not significant when adjusted for all listed covariates. In a case-control study of 158 women (80 cases and 78 controls), Porpora
et al. [
42] found the GM of total PCBs to be significantly higher in cases than controls (301.3
vs. 203.0,
p < 0.01). The OR of endometriosis risk in the highest tertile of total PCBs compared with the lowest tertile, was 5.63 (95% CI 2.25–14.10). Significant increased risk of endometriosis was also found for PCB congeners 118, 138, 153, and 170. Heiler
et al. [
38] conducted a case-control study of 50 cases (25 with peritoneal endometriosis (PE) and 25 with deep endometriotic (DE) nodules) and 21 controls. Multiple dioxin-like PCBs were measured and expressed as toxic equivalent (TEQ) per gram of serum lipids. Dioxin-like PCB concentrations were higher in women with DE compared to controls {12.4 (10.3 − 14.9)
vs. 8.5 (6.9 − 10.5),
p = 0.026} but did not significantly differ for women with PE compared to controls {11.0 (9.1 − 13.3)
vs. 8.5 (6.9 − 10.5)} and for women with DE compared to women with PE (12.4
vs. 11.0).
Four of the PCB case-control studies failed to find significant associations between endometriosis and exposure to individual PCB congeners, total PCBs, or specific sub-groups [
38,
40,
42,
43]. Niskar
et al. [
40] conducted a case-control study with 60 confirmed endometriosis cases staged as I (minimal), II (mild), III (moderate), and IV (severe) and 30 controls. Mean lipid-adjusted PCB concentrations were not significantly different (179.98
vs. 217.33
vs. 194.76
vs. 193.37) between stage I–II cases, stage III cases, stage IV cases, and controls, respectively. In the largest case-control study (Trabert
et al. 2010 [
43]), total PCBs (
n = 20), estrogenic PCBs (
n = 6), and individual PCB congeners were measured in the serum from 251 cases and 538 controls, matched for age and reference year. Adjusted total and estrogenic PCBs in the highest quartiles were not associated with an increased risk of endometriosis (Total: OR = 1.2, 95% CI 0.6–2.3, Estrogenic: OR = 0.9, 95% CI 0.5–1.4). In two case-control studies measuring median TEQ values (pg TEQ/g lipid) Pauwels
et al. [
43] found no association between endometriosis and the median TEQ values (pg TEQ/g lipid) in cases and controls (29
vs. 27) and Tsukino
et al. [
44] found no difference in median TEQ values for endometriosis cases (stage II–IV) and controls (stage 0–I) (cPCBs: 3.40
vs. 3.59, PCBs: 4.61
vs. 5.14), respectively. The OR of endometriosis risk in the highest quartile of total PCBs compared with the lowest quartile was 0.41 (95% CI 0.14–1.27).
Like breast cancer, results of the association between PCB exposure and endometriosis in eight epidemiologic studies were inconsistent or conflicting; therefore, we extracted and summarized risk estimates of PCBs on endometriosis from four case control studies using meta-analytic methods. Combining four studies of exposure to PCBs produced a summary risk estimate of 1.91 (95% CI: 1.05–5.54) (
Table 4;
Figure 4). PCBs exposures were found to be significantly associated with development of endometriosis as a meta-analysis of four studies produced an increased risk of 1.91. However, there is not much confidence in the combined risk estimate of endometriosis with exposure to PCBs because of the lower estimate of CI being barely higher than 1 (1.05).
Figure 4.
Forest plot of epidemiological studies of the associations between exposure to PCBs and risk of endometriosis.
Figure 4.
Forest plot of epidemiological studies of the associations between exposure to PCBs and risk of endometriosis.
Table 4.
Epidemiological Studies of the Association between Exposure to PCBs and Risk of Endometriosis.
Table 4.
Epidemiological Studies of the Association between Exposure to PCBs and Risk of Endometriosis.
| Reference, Location | Study Design | Study Population | Measurement of Exposure | Outcomes | Results | Comments | Confounders |
|---|
| Heiler et al. [38], Belgium | Case-control study | 50 cases: (25 with PE, 25 with DE), 21 controls | Multiple PCBs from serum, 12 dioxin-like PCBs (pg TEQ/g lipids). | Mean serum PCB Range (pg TEQ/g lipids): Controls: 6.9–10.5; PE Cases: 9.1–13.3; DE Cases: 0.3–14.9; Logistic Regression (OR, 95% CI). | Significant risk with DE nodules (OR = 6.7; 95% CI, 1.4–31.2). | Controls did not present for infertility; normal pelvic exam. Cases confirmed with histological exam of lesions. | Adjusted for age, BMI, tobacco consumption, age at menarche, duration of OC use, family history, menstrual cycle regularity, # of children, breast-feeding duration. |
| Niskar et al. [40], USA | Case-control study | 60 cases, 30 controls/ 64 controls | Serum total PCBs (ng/g) (n = 36). | GM Total PCBs (ng/g lipid): Cases stage I–II (179.98), stage III (217.33), stage IV (194.76), Controls (193.37). Logistic Regression (OR, 95% CI). | No significant differences in GMs (p = 0.97). No significant associations (OR = 1.00, 95% CI 0.99–1.01). | Cases confirmed with laparoscopic examination and/or biopsy. 30 controls confirmed with laparoscopy, 27 with infertile partner and 7 with ovulation problems. | Adjusted for age, gravidity, education, income. |
| Pauwels et al. [41], Belgium | Prospective case-control study | 42 cases, 27 controls | Multiple PCBs from serum; Total PCBs, TEQ (pg TEQ/g lipid). | Median TEQ (pg TEQ/g lipid): Cases (29), Controls (27). Logistic Regression (OR, 95% CI). | No significant associations found (OR = 4.33, 95% CI 0.49–38.19). | Cases and controls infertile. Endometriosis confirmed with laparoscopic examination. | Age, BMI, alcohol consumption. |
| Porpora et al. [42], Italy | Case-control study | 80 cases, 78 controls | Multiple PCBs from serum, Total PCBs. | GM of Total PCBs (ng/g of fat): Cases: 301.3; Controls: 203.0; Logistic Regression (OR, 95% CI). | Total PCB concentrations significantly higher in cases (OR = 5.63, 95% CI 2.25–14.10); Significant increased risk for PCBs 118, 138, 153, and 170 for 2nd and 3rd tertiles when compared to the lowest tertile. | Cases and controls confirmed with laparoscopic examination. | Adjusted for age, BMI, smoking habits, weight modification. |
| Trabert et al. [43], USA | Case-control study | 251 cases, 538 controls; matched for age (5 year) and reference year | Multiple PCB congeners in serum (n = 20); Total PCBs, Estrogenic PCBs. | Logistic Regression (OR, 95% CI); Quartiles. | No significant associations found. | Cases: Group Health (GH) enrollees with endometriosis diagnosis, Controls: randomly selected from list of GH enrollees. | Adjusted for matching factors, serum lipids, income, alcohol consumption, DDE exposure. |
| Tsukino et al. [44], Japan | Case-control study | 139 women: Controls: Stage 0 & I, Cases: Stage II–IV; Stage 0 = 59 Stage I = 22 Stage II = 10 Stage III = 23 Stage IV = 25 | Multiple PCBs in serum; Total TEQ values of cPCBs and PCBs. | Median TEQ values (pg TEQ/g lipid); Logistic Regression (OR, 95% CI); Quartiles. | No significant associations found (OR = 1.2, 95% CI 0.6–2.3). | Cases and controls confirmed with laparoscopic examination. | Adjusted for menstrual regularity and average cycle days. |
Table 5 lists epidemiological studies of the association between EDCs-phthalate or BPA and endometriosis. No meta-analysis was performed on exposure to BPA or phthalates and endometriosis, because only two studies that met our criteria of selection examined the association between endometriosis and phthalates [
39,
44]; one study addressed the association between endometriosis and BPA [
45], and one study addressed the association between both BPA and phthalates and endometriosis [
29]. Besides these two studies, there are several other epidemiological studies that have examined the association between phthalate or BPA exposure and endometriosis [
10,
29,
37,
39,
46,
48,
49,
50],
Table 5. Kim
et al. [
39] measured plasma levels of mono (2-ethylhexyl) phthalate (MEHP) and di-(2-ethylhexyl) phthalate (DEHP) in 97 women with advanced-stage endometriosis and 169 control women. Mean plasma levels of MEHP and DEHP were found to be significantly higher in cases than controls (MEHP: 17.4
vs. 12.4,
p < 0.001, DEHP: 179.7
vs. 92.5,
p = 0.010). In a population-based case-control study conducted by Upson
et al. [
45] 8 urinary phthalate metabolites were measured in 92 surgically-confirmed endometriosis cases and 195 controls. A significant inverse association was found between urinary MEHP and risk of endometriosis (OR = 0.3, 95% CI 0.1–0.7). The ENDO study was designed to assess the relationship between exposure to environmental chemicals and endometriosis. Louis
et al. [
46] analyzed 14 phthalate metabolites and total BPA in urine from 495 women who underwent laparoscopy (operative cohort) and 131 women (population cohort) who underwent pelvic magnetic resonance imaging (MRI) for the assessment of endometriosis. In the operative cohort, GMs of phthalate metabolites were not found to be significantly higher in women with endometriosis, whereas, in the population cohort, GMs of six phthalate metabolites were found to be significantly higher for women with endometriosis and a two-fold or higher increase in ORs was observed for mono-
n-butyl phthalate (mBP), mono-(2-ethyl-5-carboxyphentyl) phthalate (mECPP), mono-[(2-carboxymethyl) hexyl] phthalate (mCMHP), mono (2-ethyl-5-hydroxyhexyl) phthalate (mEHHP), mono (2-ethyl-5-oxohexyl) phthalate (mEOHP), and mono (2-ethylhexyl) phthalate (mEHP). No significant associations were found for urinary BPA concentrations in either the operative cohort or the population cohort. In a hospital based cross-sectional study, conducted by Itoh
et al. [
51], urinary BPA concentrations were analyzed in 140 women who underwent laparoscopy. The severity of endometriosis was classified into five stages: 0 (
n = 60), I (
n = 21), II (
n = 10), III (
n = 24), and IV (
n = 25). Median creatinine adjusted urinary BPA concentrations did not significantly differ by endometriosis stage (0.74
vs. 0.93,
p = 0.24) for stages 0–I and stages II–IV, respectively.
Table 5.
Epidemiological studies of the association between EDCs-Phthalate or BPA and endometriosis.
Table 5.
Epidemiological studies of the association between EDCs-Phthalate or BPA and endometriosis.
| EDCs | Biological Samples | Study Population | Outcomes | References |
|---|
| Bisphenol A | Serum | 69 fertile women undergoing laparoscopy, Naples, Italy | Detected in cases | Cobellis et al. [47 |
| Bisphenol B | Serum | 69 fertile women undergoing laparoscopy, Naples, Italy | Detected in cases | Cobellis et al. [47] |
| Phthalate esters | Plasma | 220 South Indian women undergoing laparoscopy | Increased risk | Reddy et al. [37] |
| Serum | 108 South Indian women undergoing laparoscopy | Increased risk | Reddy et al. [50] |
| Diethylphthalate | Blood/perit | 59 fertile women undergoing laparoscopy | Higher in cases | Cobellis et al. [47] |
| Monoethylphthalate | Blood/peri-toneal fluid | 59 fertile women undergoing laparoscopy | No association | Cobellis et al. [47] |
| Monobutylphthalate | Urine | 1227 women from the NHANES study, United States | No association | Calafat et al. [10] |
| Urine | 109 women undergoing laparotomy, Taiwan | Increased in cases | Huang et al. [48] |
| Monobutylphthalate | Urine | 1227 women from the NHANES study, USA | No association | Calafat et al. [10] |
| Urine | 109 women undergoing laparotomy, Taiwan | Increased in cases | Huang et al. [48] |
Table 6.
Genes interacting with polychlorinated biphenyls in endometriosis.
Table 6.
Genes interacting with polychlorinated biphenyls in endometriosis.
| IUPAC Name (Congener Number) | Interacting Genes |
|---|
| Polychlorinated Biphenyls | 19 genes: AKR1C3 | ANKRD1 | AREG | ARNT | CYP19A1 | DUSP1 | ESR2 | FBN1 | FOS | GREB1 | IGFBP1 | KRAS | NR2C2 | NR3C1 | PAPPA | PTGER4 | STC2 | TGFB2 | THRA |
| 2,4,4ʹ-Trichlorobiphenyl (28) | 2 genes: ESR2 | NR3C1 |
| 3,4,3ʹ,4ʹ-Tetrachlorobiphenyl (77) | 11 genes: ARNT | DDX5 | ESR2 | FKBP5 | ITGB8 | KLF13 | MAOB | NR1D2 | PRLR | SULF2 | TXNIP |
| 2ʹ,3,3ʹ,4ʹ,5-Pentachloro-4-hydroxybiphenyl (4ʹ-OH-PCB-86; 4-hydroxy-2,2ʹ,3ʹ,4ʹ,5ʹ-pentachlorobiphenyl ) | 25 genes: ABCC9 | BRD8 | CD55 | CNR1 | ELAVL1 | ERRFI1 | FKBP5 | IFNGR1 | IGFBP1 | MED1 | MED4 | MTA1 | NCOA6 | NR2C1 | NR3C1 | NR3C2 | NR4A1 | NRP1 | PRLR | SLC16A6 | SPARCL1 | SST | TAGLN | THRA | TNC |
| 2,2ʹ,4,6,6ʹ-Pentachlorobiphenyl (104) | 2 genes: EGFR | FOS |
| 3,4,5,3ʹ,4ʹ-Pentachlorobiphenyl (126) | 36 genes: AREG | CD55 | CXCL14 | CYP19A1 | DUSP1 | ENPP1 | FBN1 | GPX3 | HBEGF | HSD17B1 | HSD17B2 | IGF1 | IGFBP1 | IGFBP6 | IL1R1 | IMPA2 | MAOA | MAOB | MED1 | NEDD4L | NR3C1 | OSR2 | PRLR | RASL11A | SEPP1 | SLC20A1 | SLC40A1 | SLC7A8 | SPARCL1 | SRD5A1 | SRD5A2 | SST | STC2 | TAGLN | TGFB2 | TXNIP |
| 2,2ʹ,3ʹ,4,4ʹ,5-Hexachlorobiphenyl (137) | 10 genes: FBLN1 | FOS | HBEGF | IGF1 | NEFM | PRL | SLC16A6 | SRD5A1 | SRD5A2 | SST |
| 2,3,6,2ʹ,3ʹ,6ʹ-Hexachlorobiphenyl (136) | 2 genes: AR | CYP2B1 |
| 2,4,5,2ʹ,4ʹ,5ʹ-Hexachlorobiphenyl (153) | 18 genes: CYP19A1 | DCSTAMP | EGFR | ESR2 | FOS | HSD17B1 | HSD17B2 | IFIT1 | IGF1 | ITGB8 | MAOB | NR3C1 | SEPP1 | SLC16A6 | SRD5A1 | SRD5A2 | SST | TXNIP |
| 17β Estradiol | 114 genes: ABCC9 | ABI3BP | ACTA2 | AKR1C1 | AKR1C2 | AKR1C3 | ANKH | ANKRD1 | AREG | ARHGAP28 | ARNT | BMP7 | C10ORF10 | C1R | CCNE2 | CD55 | CFD | CLDN1 | CNIH3 | CNR1 | CPM | CXCL14 | CYB5A | CYP19A1 | CYP26A1 | DDX5 | DICER1 | DIO2 | DKK1 | DUSP1 | EGFR | ELAVL1 | ERRFI1 | ESR2 | FBLN1 | FBN1 | FKBP5 | FOS | GPX3 | GREB1 | HDAC1 | HDAC2 | HERC5 | HS3ST3B1 | HSD17B1 | HSD17B2 | IDO1 | IFIT1 | IGF1 | IGFBP1 | IGFBP6 | IHH | IL15 | IL1R1 | IL7R | ITGA2 | ITGB1 | ITGB8 | KLF13 | KLF9 | KRAS | LMOD1 | LTF | MAOA | MAOB | MED1 | MED14 | METTL7A | MIR21 | MYLIP | NCOA1 | NCOA6 | NCOR1 | NEDD4L | NR2F2 | NR3C1 | NR3C2 | NR4A1 | NR5A1 | NRP1 | NTRK3 | OLFM4 | OSR2 | PAPPA | PGR | PRL | PRLR | PTGER2 | PTGER4 | RARB | RASGRP1 | RGS4 | RORB | RXFP1 | SEPP1 | SLC16A6 | SLC1A1 | SLC20A1 | SLC40A1 | SLC7A8 | SMPDL3A | SPARCL1 | SRD5A2 | STC2 | SULF2 | TACSTD2 | TAGLN | TGFB2 | THRA | TNC | TOB1 | TRH | TXNIP | VCAN ZEB2 |
| Dibutyl phthalate | 71 genes: ABI3BP | ACTA2 | AKR1C1 | ANKRD1 | AREG | ARNT | BMP7 | BRD8 | C1R | CCNE2 | CD55 | CLDN1| CNR1 | COPS2 | CYB5A | CYP19A1 | CYP26A1 | DDX5 | DICER1 | DUSP1 | EGFR | ELAVL1 | ENPP1 | ERRFI1 | ESR2 |FKBP5 | FOS | GPX3 | HDAC1 | HSD17B1 | IGF1 | IL1R1 | ITGB1 | ITGB8 | KLF9 | LMOD1 | MAOA | MAOB | MED1 |MED14 | MED17 | NR1D2 | NR2F2 | NR2F6 | NR3C1 | NR4A1 | NR5A1 | NRP1 | OSR2 | PAPPA | PGR | PRLR | PTGER2 |PTGER4 | RASL11A | SEPP1 | SLC16A6 | SLC20A1 | SLC40A1 | SLC7A8 | SMPDL3A | SRD5A1 | STC2 | SUCLG2 |SULF2 | TAGLN | TGFB2 | THRA | TOB1 | TXNIP | VCAN. |
| Diethylhexyl phthalate | 29 genes: CNR1 | CYP19A1 | CYP26A1 | EGFR | ESR2 | FKBP5 | FOS | HERC5 | IGF1 | IGFBP1 | ITGB1 | MAOA | NCOA1 | NR3C1 | NR4A1 | NR5A1 | PAX2 | PRL | PRLR | PTGER2 | PTGER4 | SRD5A1 | TGFB2 | DUSP1 | FMO2 | GPX3 | MED1 | NCOR1 | TXNIP |
| Dibutyl phthalate and diethyl-hexyl phthalate | 22 genes: CNR1 | CYP19A1 | CYP26A1 | DUSP1 | EGFR | ESR2 | FKBP5 | FOS | GPX3 | IGF1 | ITGB1 | MAOA | MED1 | NR3C1 | NR4A1 | NR5A1 | PRLR | PTGER2 | PTGER4 | SRD5A1 | TGFB2 | TXNIP |
| Bisphenol A | 80 genes: ABCC9 | ACTA2 | AREG | ARHGAP28 | ARNT | BMP7 | BRD8 | CCNE2 | COPS2 | CYB5A | CYP19A1 | CYP26A1 | DDX5 | DICER1 | DIO2 | DUSP1 | EGFR | ELAVL1 | ENPP1 | ERRFI1 | ESR2 | FKBP5 | FOS | GPX3 | GREB1 | HDAC1 | HDAC2 | HSD17B1 | HSD17B2 | IFNGR1 | IGF1 | IGFBP1 | IGFBP6 | ITGB8 | KLF9 | KRAS | LMOD1 | LTF | MAOA | MED1 | MED14 | MED16 | MED17 | MED4 | MIR21 | NCOA1 | NCOR1 | NR2C1 | NR2F2 | NR3C1 | NR3C2 | NR4A1 | NR5A1 | NRP1 | OLFM4 | PAPPA | PGR | PRL | PRLR | PTGER2 | PTGER4 | RASGRP1 | RASL11A | RGS4 | RORB | SLC1A1 | SLC40A1 | SLC7A8 | SMPDL3A | SRD5A1 | SRD5A2 | SST | STC2 | SULF2 | TACSTD2 | TAGLN | TGFB2 | THRA | TNC | VCAN |
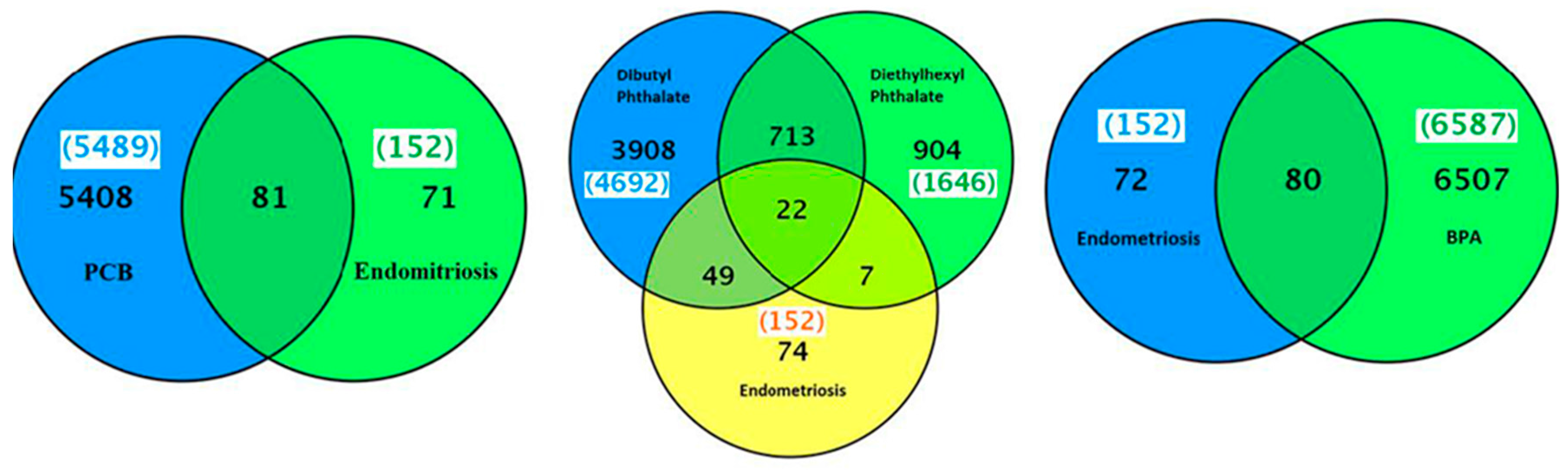
Several hundred genes were altered by exposure to PCBs, phthalate or BPA (
Figure 5). The genes related to PCB and PCB congeners-3,4,3′,4′-tetrachlorobiphenyl (77), 3,4,5,3′,4′-pentachlorobiphenyl (126), and 2,4,5,2′,4′,5′-hexachlorobiphenyl (153) in endometriosis were 19, 11, 36 and 18, respectively (
Table 6). All these PCBs or their congener-related endometriosis genes were also associated with 17β-estradiol. The top interacting genes with PCBs and endometriosis were
ESR2,
NR3C1,
CYP19A1,
EGFR,
FKBP5,
ITGB8,
MAOB,
PGR,
PRLR,
SLC16A6,
SST, and
TXNIP. There were 80 common genes found between BPA and endometriosis (
Figure 5). The two phthalates with the most gene interactions were: dibutyl phthalate and diethylhexyl phthalate. The genes related to dibutyl phthalate and diethylhexyl phthalate were 4692 and 1646, respectively. There were 71 common genes associated between dibutyl phthalate and endometriosis and 29 common genes between diethylhexyl phthalate and endometriosis (
Figure 5). There were 22 genes in common between both phthalates-dibutyl phthalate and diethylhexyl phthalate, and endometriosis, as shown in
Table 6. Interactions among these genes are shown in
Figure S2. Enrichment pathway analysis revealed that some of these genes are part of: (1) pathways in cancer (KEGG:05200); (2) signal transduction (REACT:111102); and (3) MAPK signaling pathway ( KEGG:04150) (
Table 3).
Figure 5.
A Venn diagram of list of genes common between endometriosis and PCBs, phthalates or bisphenol A.
Figure 5.
A Venn diagram of list of genes common between endometriosis and PCBs, phthalates or bisphenol A.
2.6. Integration of Genes Overlapped among EDCs, Breast Cancer and Endometriosis
Integration of genes associated with exposure to PCBs, and breast cancer and endometriosis based enriched disease analysis showed that there were 16 endometriosis genes overlapped with breast neoplasms—
AREG,
C10ORF10,
CLDN1,
CYP19A1,
DKK1,
EGFR,
ESR2,
FOS,
IGF1,
KRAS,
NCOA1,
NCOR1,
NR2F6,
PGR,
RARB, and
STC2 (
Table 2 and
Table 6). All of these genes were also associated with estrogen in breast neoplasms. Out of these 16 genes, there were 14 genes—
AREG,
CLDN,
CYP19A1,
DKK1,
EGFR,
ESR2,
FOS,
IGF1,
KRAS,
NCOA1,
NCOR1,
NR2F6,
PGR,
RARB, and
STC2—common among 17β-estradiol, breast cancer, and endometriosis (
Table 2,
Table 6 and
Table 7). Total PCBs associated with
AREG,
CYP19A1,
ESR2,
FOS,
KRAS and
STC2 genes; PCB 126 associated with
AREG,
CYP19A1, and
STC2 genes and PCB 15 associated with
CYP19A1,
EGFR,
ESR2,
FOS, and
IGF1 genes overlapped with 17β-estradiol, breast cancer, and endometriosis (
Table 2,
Table 6 and
Table 7). Similarly, we identified dibutyl phthalate and diethyl-hexyl phthalate associated overlapping genes with 17β-estradiol, breast cancer, and endometriosis:
AREG,
CLDN1,
CYP19A1,
EGFR,
ESR2,
FOS,
IGF1,
NR2F6,
PGR and STC2; and
CYP19A1,
EGFR,
ESR2,
FOS,
IGF1, and
NCOA1. There were five common overlapped genes between these two phthalates, 17β-estradiol, breast cancer and endometriosis:
CYP19A1,
EGFR,
ESR2,
FOS, and
IGF1. We also identified another 11 EDC–BPA associated genes that were common among 17β-estradiol, breast cancer and endometriosis:
AREG,
CYP19A1,
EGFR,
ESR2,
FOS,
IGF1,
KRAS,
NCOA1,
NCOR1,
PGR, and STC2. Five genes—CYP19A1, EGFR, ESR2, FOS, and IGF1—were common among all three EDCs–PCBs, phthalates and BPA, 17β-estradiol, breast cancer, and endometriosis. For the gene ontology terms associated with each gene, please see
Table 8.
Since both of these diseases are dependent on unopposed estrogen for their growth, we examined whether estrogen receptor signaling pathway genes are common among estrogen, EDCs, breast cancer and endometriosis. PCBs and congeners 3,4,5,3′,4′-pentachlorobiphenyl (126) and 2,4,5,2′,4′,5′-hexachlorobiphenyl (153) were associated with some of the same estrogen receptor signaling pathway genes—
AR,
ESR1,
ESR2,
NCOA3, and
PPARGC1B;
AR,
BRCA1,
ESR1,
IGF1, and
PAK1; and
AR,
BRCA1,
CTNNB1,
ESR1,
ESR2,
IGF1, and
SRC, respectively (
Table 7). The following were also observed with 17β-estradiol—
AR,
BRCA1,
CCNE1,
CTNNB1,
ESR1,
ESR2,
FHL2,
FOXA1,
IGF1,
NCOA1,
NCOA2,
NCOA3,
NRIP1,
PAK1,
PGR,
PHB,
PPARGC1B,
RB1,
SFRP1,
SRC, and
ZNF366. Similarly, common genes of estrogen receptor signaling pathways were also observed with another three EDCs. Dibutyl phthalate associated genes,
AR,
BRCA1,
CCNE1,
CTNNB1,
ESR1,
ESR2,
FHL2,
HEYL,
IGF1,
PGR,
RB1, and
SRC; and diethylhexyl phthalate associated genes,
AR,
CTNNB1,
ESR1,
ESR2,
IGF1,
NCOA1, and
PPARGC1B, and BPA associated
AR,
BRCA1,
CCNE1,
CTNNB1,
ESR1,
ESR2,
FHL2,
IGF1,
NCOA1,
NCOA2,
NCOA3,
NRIP1,
PAK1,
PGR,
PHB,
RB1,
SFRP1,
SIRT1, and
SRC, are also associated with 17β-estradiol in breast neoplasms (
Table 7).
Table 7.
EDCs observed in breast neoplasms that are associated with estrogen responsive gene interactions, endometriosis, and inflammation.
Table 7.
EDCs observed in breast neoplasms that are associated with estrogen responsive gene interactions, endometriosis, and inflammation.
| EDC Interacting with Genes in Breast Neoplasms | Steroid Hormone Receptor Signaling Pathway | Endometriosis | Inflammation |
|---|
| 17β Estradiol | AR, BRCA1, CCNE1, CTNNB1, ESR1, ESR2, FHL2, FOXA1, IGF1, NCOA1, NCOA2, NCOA3, NRIP1, PAK1, PGR, PHB, PPARGC1B, RB1, SFRP1, SRC, ZNF366 | AREG, CLDN1, CYP19A1, DKK1, EGFR, ESR2, FOS, IGF1, KRAS, NCOA1, NCOR1, PGR, RARB, STC2 | AHR, CSF3, CXCL2, CXCL8, HMOX1, IFNG, IL1B, IL6, LEP, MIF, MMP9, NOS2, NOS3, PARP1, PTGS2, SOD2, TFRC, TNF |
| PCBs | AR, ESR1, ESR2, NCOA3, PPARGC1B | AREG, CYP19A1, SR2, FOS, KRAS, STC2 | AHR, CXCL2, HMOX1, IFNG, IL6, PTGS2, SOD2, TNF |
| 3,4,5,3′,4′-Pentachlorobiphenyl (126) | AR, BRCA1, ESR1, IGF1, PAK1 | AREG, CYP19A1, STC2 | AHR, CXCL8, HMOX1, IL1B, IL6, MMP9, NOS2, NOS3, PARP1, PTGS2, TNF |
| 2,4,5,2′,4′,5′-Hexachlorobiphenyl (153) | AR, BRCA1, CTNNB1, ESR1, ESR2, IGF1, SRC | CYP19A1, EGF, ESR2, FOS, IGF1 | AHR, IFNG, IL1B, PARP1, PTGS2, TNF |
| Dibutyl Phthalate | AR, BRCA1, CCNE1, CTNNB1, ESR1, ESR2, FHL2, HEYL, IGF1, PGR, RB1, SRC | AREG, CLDN1, CYP19A1, EGFR, ESR2, FOS, IGF1, R2F6, PGR, STC2 | AHR, CXCL8, HMOX1, IL1B, IL6, MIF, MMP9, PARP1, SOD2, TFRC, TNF |
| Diethylhexyl Phthalate | AR, CTNNB1, ESR1, ESR2, IGF1, NCOA1, PPARGC1B | CYP19A1, EGFR, ESR2, FOS, IGF1, NCOA1 | AHR, CSF2, CXCL8, IFNG, LEP, MMP9, SOD2, TNF |
| Bisphenol A | AR, BRCA1, CCNE1, CTNNB1, ESR1, ESR2, FHL2, IGF1, NCOA1, NCOA2, NCOA3, NRIP1, PAK1, PGR, PHB, RB1, SFRP1, SIRT1, SRC | AREG, CYP19A1, EGFR, ESR2, FOS, IGF1, KRAS, NCOA1, NCOR1, PGR, STC2 | AHR, CSF2, HMOX1, IFNG, IL1B, IL6, LEP, MIF, MMP9, NOS2, NOS3, PARP1, PTGS2, SOD2, TNF |
Table 8.
Integration of changes in the expression of genes showing common genes modified in EDCs, breast cancer and endometriosis. The underlined gene names show a total of five genes that were common among all three EDCs (PCBs, phthalate and bisphenol A), breast cancer, and endometriosis. Environmentally responsive genes are indicated in database column.
Table 8.
Integration of changes in the expression of genes showing common genes modified in EDCs, breast cancer and endometriosis. The underlined gene names show a total of five genes that were common among all three EDCs (PCBs, phthalate and bisphenol A), breast cancer, and endometriosis. Environmentally responsive genes are indicated in database column.
| Gene Name | Gene ID | Location | Database * | Gene Function |
|---|
| AREG | 374 | 4q13–q21 | E | Amphiregulin |
| CYP19A1 | 1588 | 15q21.1 | E | Cytochrome P450, family 19, subfamily A, polypeptide 1 |
| EGFR | 1956 | 7p12 | E | Epidermal growth factor receptor |
| ESR2 | 3468 | 14q23.2 | H | Estrogen receptor 2 (ER β) |
| FOS | 2353 | 14q24.3 | E | v-Fos FBJ murine osteosarcoma viral oncogene homolog |
| IGF1 | 3479 | 12q22-q23 | E | Insulin-like growth factor 1 (somatomedin C) |
| KRAS | 6407 | 12p12.1 | H | Kirsten rat sarcoma viral oncogene homolog |
| NCOA1 | 7668 | 2p23 | H | Nuclear receptor coactivator 1 |
| NCOR1 | 7672 | 17p11.2 | H | Nuclear receptor corepressor 1 |
| PGR | 5241 | 11q22-q23 | E | Progesterone receptor |
| STC2 | 11374 | 5q35.1 | H | Stanniocalcin 2 |
Another factor that appears to be common in both diseases is inflammation. Therefore, we also examined whether inflammation associated genes are common among estrogen, EDCs, and breast cancer. PCBs and congeners 3,4,5,3′,4′-pentachlorobiphenyl (126) and 2,4,5,2′,4′,5′-hexachlorobiphenyl (153) were associated with the following inflammation related genes—
AHR,
CXCL2,
HMOX1,
IFNG,
IL6,
PTGS2,
SOD2, and
TNF;
AHR,
CXCL8,
HMOX1,
IL1B,
IL6,
MMP9,
NOS2,
NOS3,
PARP1,
PTGS2, and
TNF; and
AHR,
IFNG,
IL1B,
PARP1,
PTGS2, and
TNF, respectively (
Table 7). Dibutyl phthalate, diethyl-hexyl phthalate and BPA-associated set of inflammation-related genes were
AHR,
CXCL8,
HMOX1,
IL1B,
IL6,
MIF,
MMP9,
PARP1,
SOD2,
TFRC, and
TNF;
AHR,
CSF2,
CXCL8,
IFNG,
LEP,
MMP9,
SOD2, and
TNF; and
AHR,
CSF2,
HMOX1,
IFNG,
IL1B,
IL6,
LEP,
MIF,
MMP9,
NOS2,
NOS3,
PARP1,
PTGS2,
SOD2, and
TNF, respectively. All of these genes were also associated with 17β-estradiol in breast neoplasms. In summary, EDC associated set of genes from inflammation pathways in breast neoplasms are estrogen responsive.
2.7. Literature Based Validation of Genes Showing Links between Endometriosis and Breast Cancer
The set of estrogen responsive genes from EDCs, environmental, inflammation, and toxicogenomics showing a link between endometriosis and breast cancer is shown in
Table 7. Research supporting the potential involvement and importance of all EDC responsive common genes in breast cancer and endometriotic lesions was found in the literature and human genome databases. The search of the environmental genome project databases showed that six genes out of 12 PCBs associated genes—
AREG,
CYP19A1,
EGFR,
FOS,
IGF1, and
PGR were environmentally responsive genes (
Table 8). These common genes were then compared to a curated list of genes in PCB exposed human cell lines. PCB congeners 77 and 153 increased the expression of the following estrogen responsive genes
AREG,
CYP19A1,
EGFR,
ESR2,
FOS,
IGF1,
KRAS,
NCOA1,
NCOR1,
NR2F6,
PGR,
STC2 [
52]. The expression of estrogen responsive genes common to breast cancer:
AREG,
CYP19A1,
EGFR,
ESR2,
FOS,
IGF1,
KRAS,
NCOA1,
NCOR1,
NR2F6,
PGR,
STC2 genes was upregulated in human endometriosis lesions [
53,
54,
55].
We also analyzed the interaction among
AREG,
CYP19A1,
EGFR,
ESR2,
FOS,
IGF1,
KRAS,
NCOA1,
NCOR1,
NR2F6,
PGR, and
STC2 genes using enrichment pathway analysis (
Figure 6). In order to investigate connections between PCBs responsive gene lists in breast cancer and endometriosis, we performed Bayesian network analysis. The Bayesian network analysis on the Cancer Genome Atlas (TCGA) Research Network data available through cbioportal.org identified the maximum likelihood structure of PCBs associated genes in breast neoplasms (
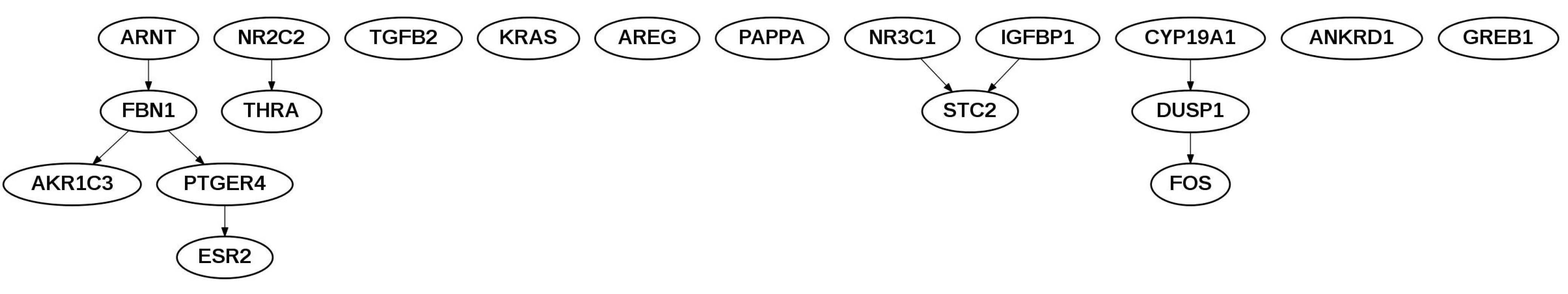
Figure 7).
Figure 7 shows plausible interactions among genes. Parents of a variable in Bayesian networks are defined as variables that arcs are originated to that variable. For example, in
Figure 7, parents of the gene
BCHE are
PTGS2 and
HMOX1. Ancestors of a variable are all the parents of the variable, all parents of parents, and so on. Arcs in
Figure 7 indicate correlations and they indicate Markov conditions. In
Figure 7, from the arcs, the relationship between
PTGS2 and
BCHE was the strongest among all pairwise relationships, but also they formed a special Y structure [
56] that indicates plausible causality,
i.e.,
PTGS2 regulating
BCHE. Similarly we have analyzed mRNA expression endometriosis data (
Figure 7). These genes were more sparsely connected.
Figure 6.
Interaction of common genes between estrogen, PCBs and breast neoplasms—AREG, CYP19A1, EGFR, ESR2, FOS, IGF1, KRAS, NCOA1, NCOR1, NR2F6, PGR, and STC2.
Figure 6.
Interaction of common genes between estrogen, PCBs and breast neoplasms—AREG, CYP19A1, EGFR, ESR2, FOS, IGF1, KRAS, NCOA1, NCOR1, NR2F6, PGR, and STC2.
Figure 7.
Identification of the maximum likelihood structure of PCBs associated genes in breast neoplasm using the Bayesian network analysis on the Cancer Genome Atlas (TCGA) Research Network data.
Figure 7.
Identification of the maximum likelihood structure of PCBs associated genes in breast neoplasm using the Bayesian network analysis on the Cancer Genome Atlas (TCGA) Research Network data.
Some of the common estrogen responsive interacting genes are part of steroid hormone biosynthesis; metabolic pathways; MAPK signaling pathway; ErbB signaling pathway; chemokine signaling pathway; p53 signaling pathway; mTOR signaling pathway; VEGF signaling pathway; focal adhesion; adherens junction; tight junction; gap junction; toll-like receptor signaling pathway; natural killer cell mediated cytotoxicity; T cell receptor signaling pathway; B cell receptor signaling pathway; Fc epsilon RI signaling pathway; regulation of actin cytoskeleton; insulin signaling pathway; GnRH signaling pathway; and pathways in cancer (
Table 3). We also compared these common genes to a curated list of genes in breast cancer, endometriosis as well as EDC exposed populations. The search of the environmental databases showed that some of these common genes were environmentally responsive. All these EDC associated set of genes are estrogen responsive (
Table 8). All these PCB, Phthalate and BPA associated common genes are altered in human breast tumor, uterine tumor tissues and endometriosis lesions (
Table 8).



 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





