
Primary Phenomenon in the Network Formation of Endothelial Cells: Effect of Charge

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
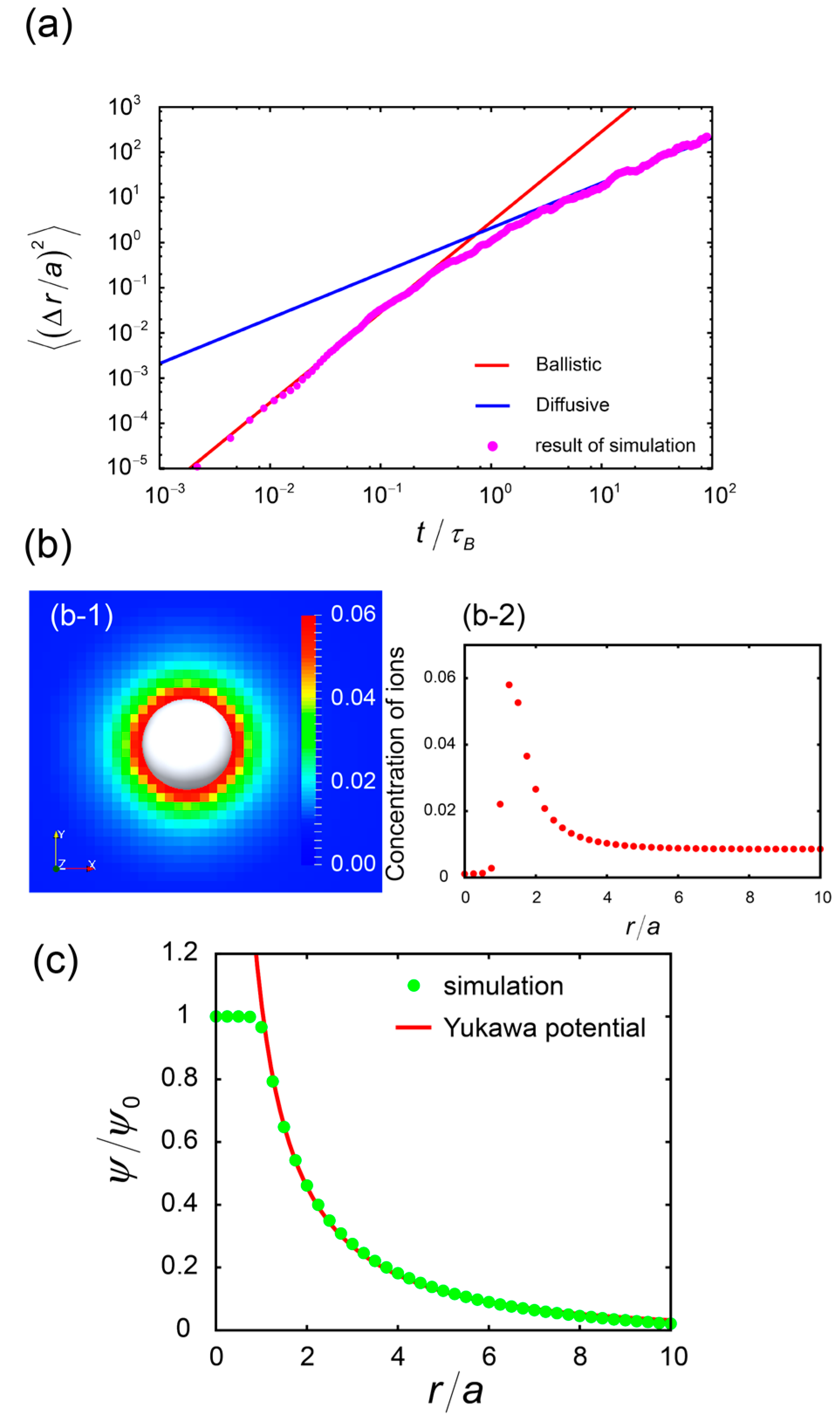{kind=link}
Abstract
:1. Introduction
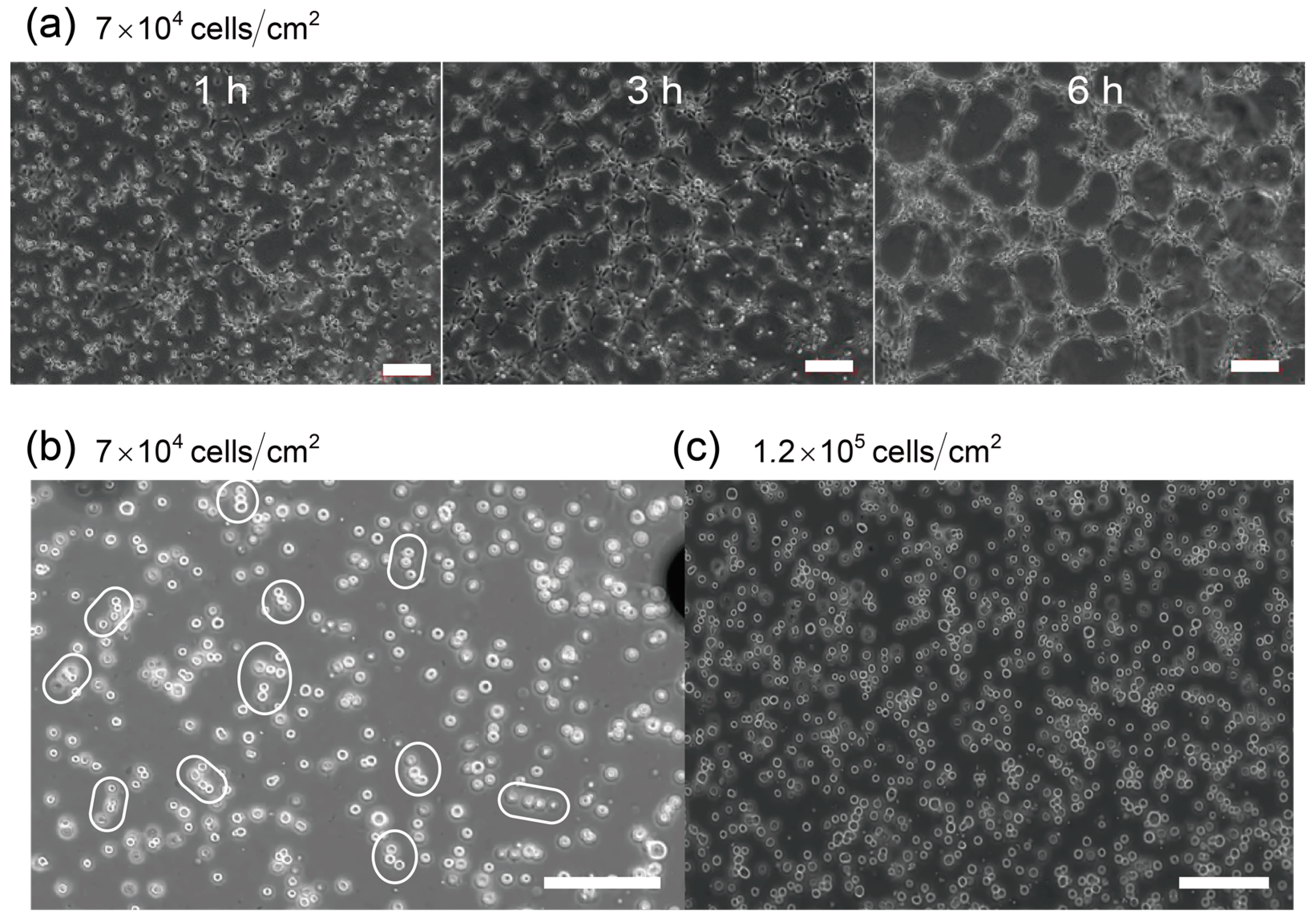
Network Formations of Endothelial Cells

2. Results and Discussion
- (1)
- Cellular Potts model
- (2)
- Continuous model
- (3)
- Lattice free particle dynamics
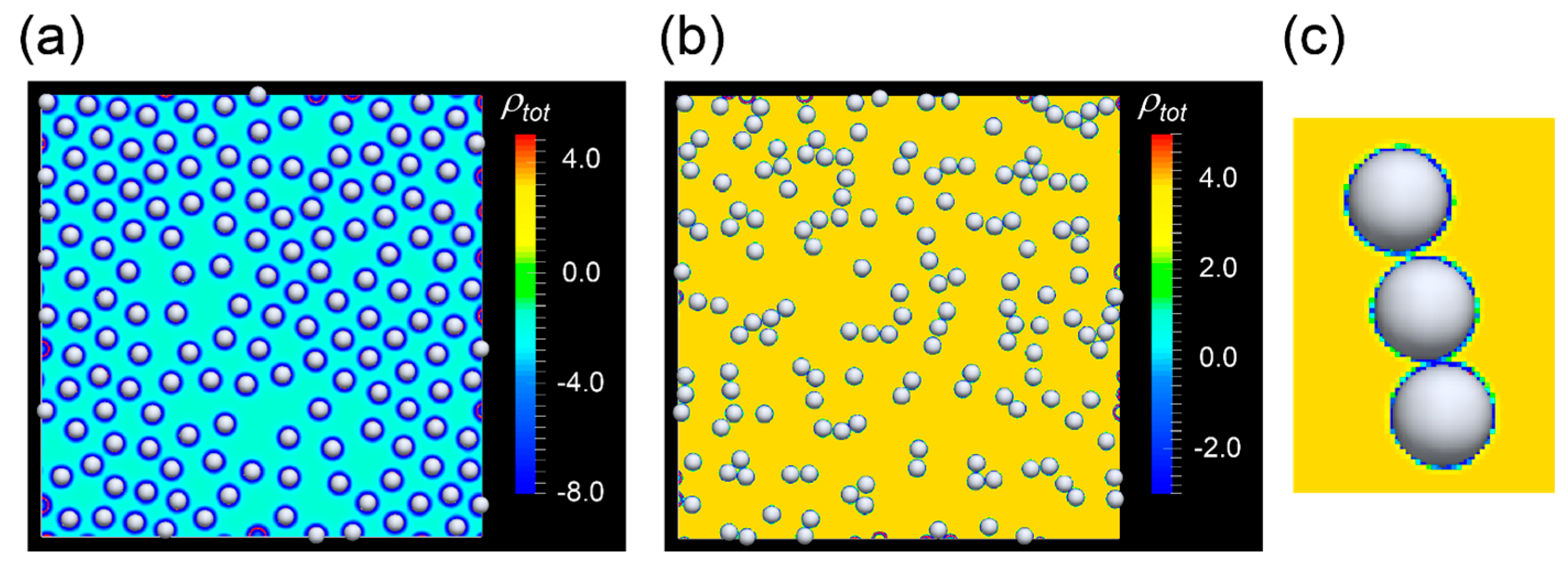
2.1. Results of FPD Simulation Including the Effect of Surface Charge of Cells

2.2. Effects of Hydrodynamic Interactions on the Formation of the Chain-Like Structure

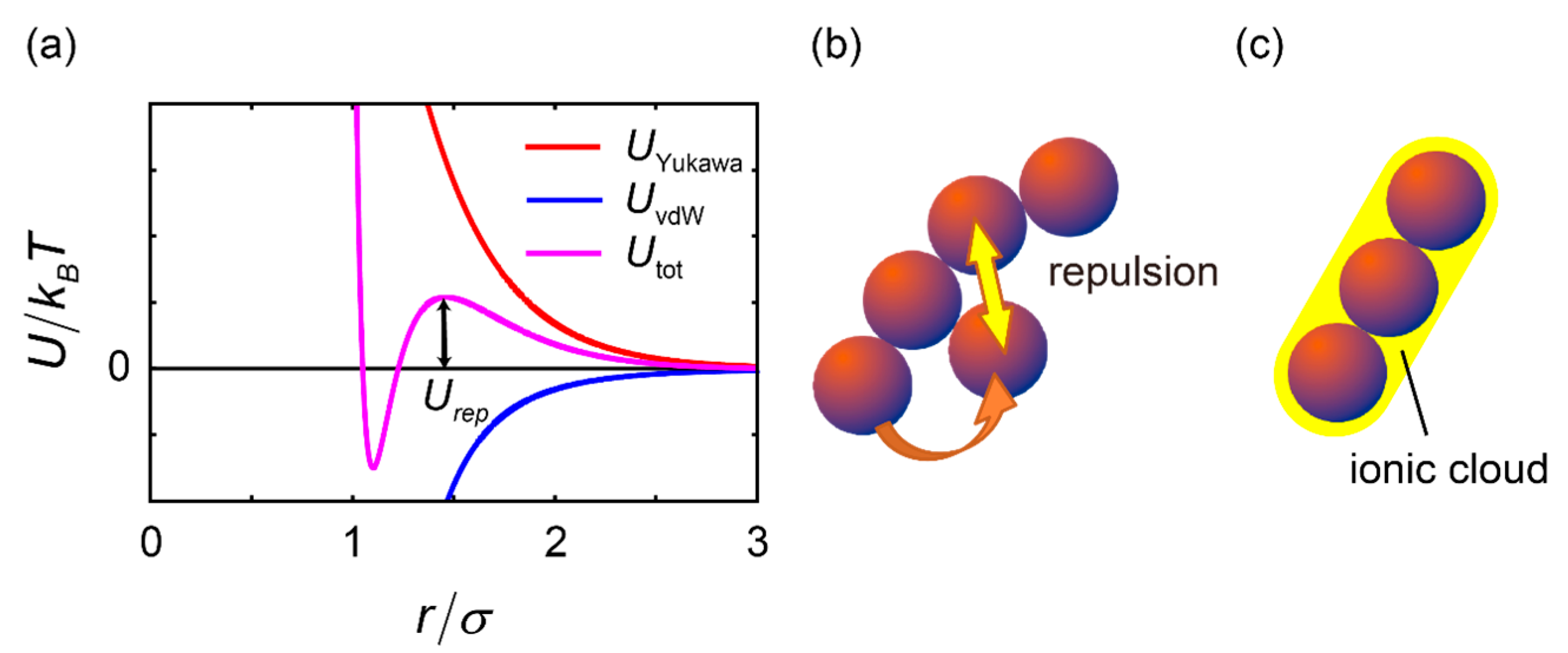
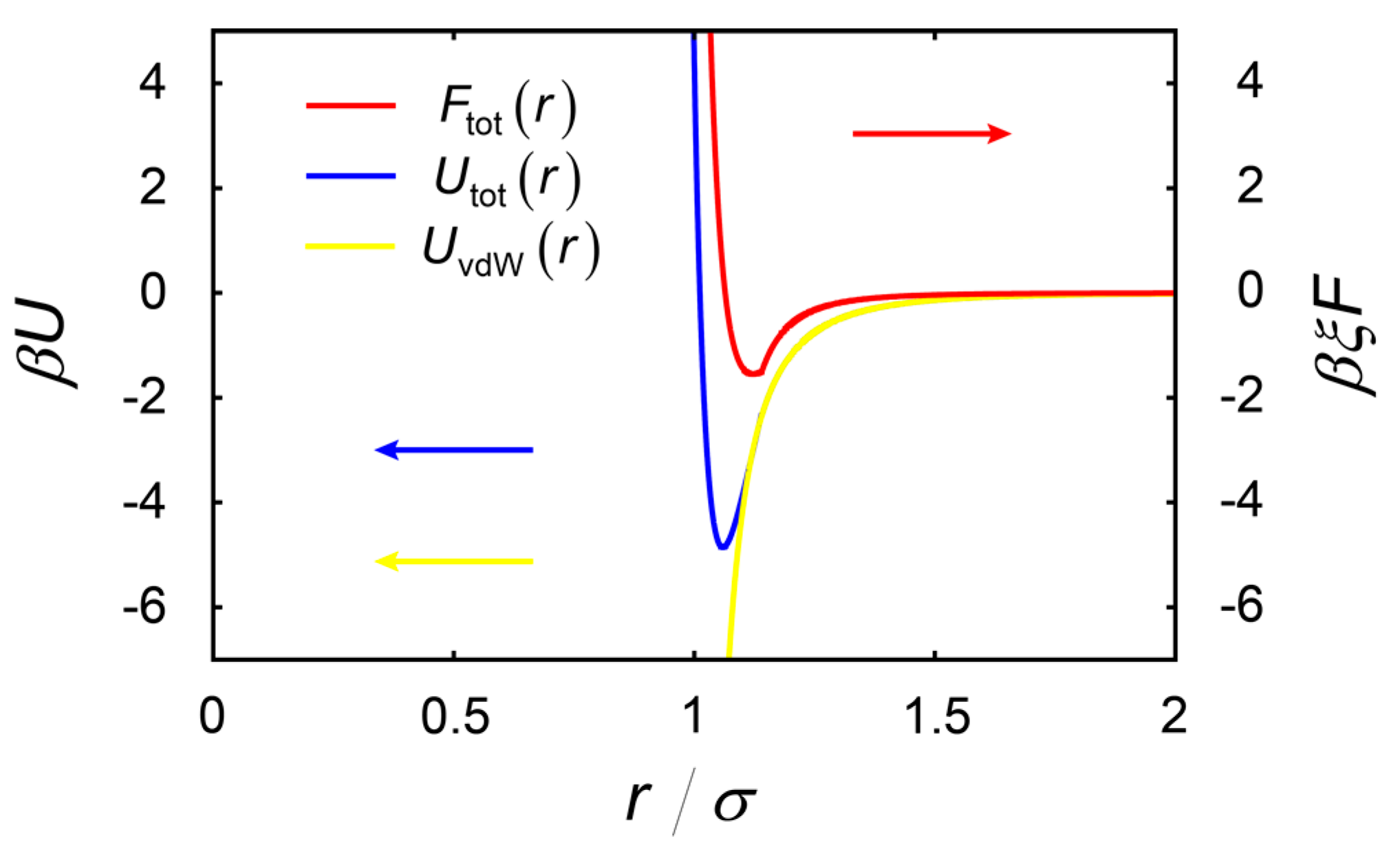
2.3. Effects of Charge on the Formation of Chain-Like Structures
2.4. Other Systems for Chain Formation


3. Materials and Methods
3.1. Simulation of Charged Particles

3.2. Dynamics of a Single Cell

3.3. Distribution of Ions and Electrostatic Behavior
3.4. Langevin Dynamics Simulation
4. Conclusions
Supplementary Materials
Acknowledgments
Conflicts of Interest
References
- Carmeliet, P. Mechanisms of angiogenesis and arteriogenesis. Nat. Med. 2000, 6, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Helmlinger, G.; Endo, M.; Ferrara, N.; Hlatky, L.; Jain, R.K. Growth factors: Formation of endothelial cell networks. Nature 2000, 405, 139–141. [Google Scholar] [CrossRef] [PubMed]
- Herrero, M.A.; Kohn, A.; Perez-Pmares, J.M. Modelling vascular morphogenesis: Current views on blood vessels development. Math. Model. Methods Appl. Sci. 2009, 19, 1483–1537. [Google Scholar] [CrossRef]
- Scianna, M.; Bell, C.; Preziosi, L. A review of mathematical models for the formation of vascular networks. J. Theor. Biol. 2013, 333, 174–209. [Google Scholar] [CrossRef] [PubMed]
- Gamba, A.; Ambrosi, D.; Coniglio, A.; de Candia, A.; Talia, S.D.; Giraudo, E.; Serini, G.; Preziosi, L.; Bussolino, F. Percolation, Morphogenesis, and Burgers Dynamics in Blood Vessels Formation. Phys. Rev. Lett. 2003, 90. [Google Scholar] [CrossRef] [PubMed]
- Serini, G.; Ambrosi, D.; Giraudo, E.; Gamba, A.; Preziosi, L.; Bussolino, F. Modeling the early stages of vascular network assembly. EMBO J. 2003, 22, 1771–1779. [Google Scholar] [CrossRef] [PubMed]
- Merks, R.M.H.; Brodsky, S.V.; Goligorksy, M.S.; Newman, S.A.; Glazier, J.A. Cell elongation is key to in silico replication of in vitro vasculogenesis and subsequent remodeling. Dev. Biol. 2006, 289, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Szabo, A.; Perryn, E.D.; Czirok, A. Network Formation of Tissue Cells via Preferential Attraction to Elongated Structures. Phys. Rev. Lett. 2007, 98. [Google Scholar] [CrossRef] [PubMed]
- Palm, M.M.; Merks, R.M.H. Vascular networks due to dynamically arrested crystalline ordering of elongated cells. Phys. Rev. E 2013, 87. [Google Scholar] [CrossRef] [PubMed]
- Lorente, S.; Wechsatol, W.; Bejan, A. Tree-shaped flow structures designed by minimizing path lengths. Int. J. Heat Mass Transf. 2002, 45, 3299–3312. [Google Scholar] [CrossRef]
- Tosin, A.; Ambrosi, D.; Preziosi, L. Mechanics and chemotaxis in the morphogenesis of vascular networks. Bull. Math. Biol. 2006, 68, 1819–1836. [Google Scholar] [CrossRef] [PubMed]
- Van Oers, R.F.M.; Rens, E.G.; la Valley, D.J.; Reinhart-King, C.A.; Merks, R.M.H. Mechanical cell-matrix feedback explains pairwise and collective endothelial cell behavior in vitro. PLoS Comput. Biol. 2014, 10, e1003774. [Google Scholar] [CrossRef] [PubMed]
- Miura, T.; Tanaka, R. In vitro Vasculogenesis Models Revisited—Measurement of VEGF Diffusion in Matrigel. Math. Model. Natl. Phenom. 2009, 4, 118–130. [Google Scholar] [CrossRef]
- Matsunaga, Y.T.; Suehiro, J.; Arai, S.; Tanaka, G. Observation of primary phenomenon in the network formation of endothelial cells in low ionic strength. In preparation. 2015. [Google Scholar]
- Tanaka, S. Simulation Frameworks for Morphogenetic Problems. Computation 2015, 3, 197–221. [Google Scholar] [CrossRef]
- Glazier, J.A.; Balter, A.; Popławski, N.J. Magnetization to Morphogenesis: A Brief History of the Glazier-Graner-Hogeweg Model. In Single-Cell-Based Models in Biology and Medicine; Alexander, R.A., Anderson, M.A.J., Katarzyna, C., Rejniak, A., Eds.; Birkhäuser Basel: Boston, MA, USA, 2007; pp. 79–106. [Google Scholar]
- Keller, E.F.; Segel, L.A. Initiation of Slime Mold Aggregation Viewed as an Instability. J. Theor. Biol. 1970, 26, 399–415. [Google Scholar] [CrossRef]
- Dillon, R.; Othmer, H.G. Mathematical Model for Outgrowth and Spatial Patterning of the Vertebrate Limb Bud. J. Theor. Biol. 1999, 197, 295–330. [Google Scholar] [CrossRef] [PubMed]
- Manoussaki, D.; Lubkin, S.R.; Vernon, R.B.; Murray, J.D. A mechanical model for the formation of vascular networks in vitro. Acta Biotheor. 1996, 44, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Doi, M.; Onuki, A. Dynamic coupling between stress and composition in polymer solutions and blends. J. Phys. II France 1992, 2, 1631–1656. [Google Scholar] [CrossRef]
- Buttà, P.; Cerreti, F.; Servedio, V.D.P.; Triolo, L. Molecular dynamics simulation of vascular network formation. J. Stat. Mech. Theory Exp. 2009, 2009. [Google Scholar] [CrossRef]
- Tanaka, H.; Araki, T. Simulation Method of Colloidal Suspensions with Hydrodynamic Interactions: Fluid Particle Dynamics. Phys. Rev. Lett. 2000, 85. [Google Scholar] [CrossRef] [PubMed]
- Hohenberg, P.C.; Halperin, B.I. Theory of dynamic critical phenomena. Rev. Modern Phys. 1977, 49. [Google Scholar] [CrossRef]
- Tanaka, H. Roles of hydrodynamic interactions in structure formation of soft matter: Protein folding as an example. J. Phys. Cond. Matter 2005, 17, S2795–S2803. [Google Scholar] [CrossRef]
- Kamata, K.; Araki, T.; Tanaka, H. Hydrodynamic Selection of the Kinetic Pathway of a Polymer Coil-Globule Transition. Phys. Rev. Lett. 2009, 102. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, A.; Tanaka, H. Key Role of Hydrodynamic Interactions in Colloidal Gelation. Phys. Rev. Lett. 2010, 104. [Google Scholar] [CrossRef] [PubMed]
- Vargas, F.F.; Osorio, M.H.; Ryan, U.S.; de Jesus, M. Surface Charge of Endothelial Cells Estimated from Electrophoretic Mobility. Membr. Biochem. 1989, 8, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.C.; Chen, I.C. Evaluation of Surface Charge Density and Surface Potential by Electrophoretic Mobility for Solid Lipid Nanoparticles and Human Brain-Microvascular Endothelial Cells. J. Phys. Chem. B 2007, 111, 11228–11236. [Google Scholar] [CrossRef] [PubMed]
- Antl, L.; Goodwin, W.J.; Hill, R.D.; Ottewill, R.H.; Owens, S.M.; Papworth, S.; Waters, J.A. The preparation of poly(methyl methacrylate) latices in non-aqueous media. Coll. Surf. 1986, 17, 67–78. [Google Scholar] [CrossRef]
- Bosma, G.; Pathmamanoharan, C.; de Hoog, E.H.; Kegel, W.K.; van Blaaderen, A.; Lekkerkerker, H.N. Preparation of Monodisperse, Fluorescent PMMA-Latex Colloids by Dispersion Polymerization. J. Coll. Interface Sci. 2002, 245, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.M.; Manoharan, V.N.; Pine, D.J.; Lange, F.F. Preparation of monodisperse PMMA microspheres in nonpolar solvents by dispersion polymerization with a macromonomeric stabilizer. Colloids Polym. Sci. 2003, 282, 7–13. [Google Scholar] [CrossRef]
- Israelachvili, J.N. Intermolecular and Surface Forces, 3rd ed.; Academic Press: Burlington, MA, USA, 2011. [Google Scholar]
- Nakayama, Y.; Yamamoto, R. Simulation method to resolve hydrodynamic interactions in colloidal dispersions. Phys. Rev. E 2005, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onuki, A. Phase Transition Dynamics; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Kodama, H.; Takeshita, K.; Araki, T.; Tanaka, H. Fluid particle dynamics simulation of charged colloidal suspensions. J. Phys. Cond. Matter 2004, 16. [Google Scholar] [CrossRef]
- Van Vlimmeren, B.A.C.; Maurits, N.M.; Zvelindovsky, A.V.; Sevink, G.J.A.; Fraaije, J.G.E.M. Simulation of 3D Mesoscale Structure Formation in Concentrated Aqueous Solution of the Triblock Polymer Surfactants (Ethylene Oxide)13(Propylene Oxide)30(Ethylene Oxide)13 and (Propylene Oxide)19(Ethylene Oxide)33(Propylene Oxide)19. Application of Dynamic Mean-Field Density Functional Theory. Macromolecules 1999, 32, 646–656. [Google Scholar]
- Karma, A.; Rappel, W.J. Phase-field model of dendritic sidebranching with thermal noise. Phys. Rev. E 1999, 60. [Google Scholar] [CrossRef]
- Abe, Y.; Ozaki, Y.; Kasuya, J.; Yamamoto, K.; Ando, J.; Sudo, R.; Ikeda, M.; Tanishita, K. Endothelial Progenitor Cells Promote Directional Three-Dimensional Endothelial Network Formation by Secreting Vascular Endothelial Growth Factor. PLoS ONE 2013, 8, e82085. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arai, S. Primary Phenomenon in the Network Formation of Endothelial Cells: Effect of Charge. Int. J. Mol. Sci. 2015, 16, 29148-29160. https://doi.org/10.3390/ijms161226149
Arai S. Primary Phenomenon in the Network Formation of Endothelial Cells: Effect of Charge. International Journal of Molecular Sciences. 2015; 16(12):29148-29160. https://doi.org/10.3390/ijms161226149
Chicago/Turabian StyleArai, Shunto. 2015. "Primary Phenomenon in the Network Formation of Endothelial Cells: Effect of Charge" International Journal of Molecular Sciences 16, no. 12: 29148-29160. https://doi.org/10.3390/ijms161226149
APA StyleArai, S. (2015). Primary Phenomenon in the Network Formation of Endothelial Cells: Effect of Charge. International Journal of Molecular Sciences, 16(12), 29148-29160. https://doi.org/10.3390/ijms161226149