
Vitamin E Content and Composition in Tomato Fruits: Beneficial Roles and Bio-Fortification


 ,
,
Abstract
:1. Introduction
2. Beneficial Roles of Vitamin E

{kind=link}
{kind=link}
| Compounds | Effect | Reference |
|---|---|---|
| VTE group | Enhanced humoral and cell immune responses reduced risk of Alzheimer’s disease | [19] |
| Membrane repair by preventing the formation of oxidised phospholipids | [20] | |
| Reduced risks of type 2 diabetes and prostate cancer | [3,22,23] | |
| Prevention of retinopathy and cataracts | [24] | |
| Reduced risk of myocardial infarction | [28] | |
| α-Tocopherol | Inhibition of HL-60 leukemic cell differentiation, reduced risk of low-density lipoprotein (LDL) oxidation and prostate carcinoma cell proliferation | [14] |
| Reduction in the serum C-reactive protein level | [30] | |
| γ-Tocopherol | Decreased adenosine diphosphate-induced platelet aggregation | [21] |
| Mixed tocotrienols | Normalized hepatic echogenic response | [29] |
3. Factors Affecting Vitamin E Content in Tomato Fresh Fruit
| Factor | Effect | Reference |
|---|---|---|
| Ripening | Increasing of α and β-tocopherols from green to red stage; increasing of γ-tocopherol from green to yellow stage | [17] |
| Water | Increasing of α-tocopherols in rain fed plants; increasing of γ-tocopherol after irrigation | [40] |
| Light | Increasing of tocopherols under high light intensity | [41] |
| Temperature | Increasing of tocopherols under low temperature | |
| salt (NaCl) | Increasing of tocopherols under salinity stress | [45] |
| Potassium | Inhibition of synthesis and/or accumulation of total tocopherols under high potassium level | [47] |
| Variety | Different level α-tocopherol in different tomato cultivars | [48,49] |
4. Impact of Processing
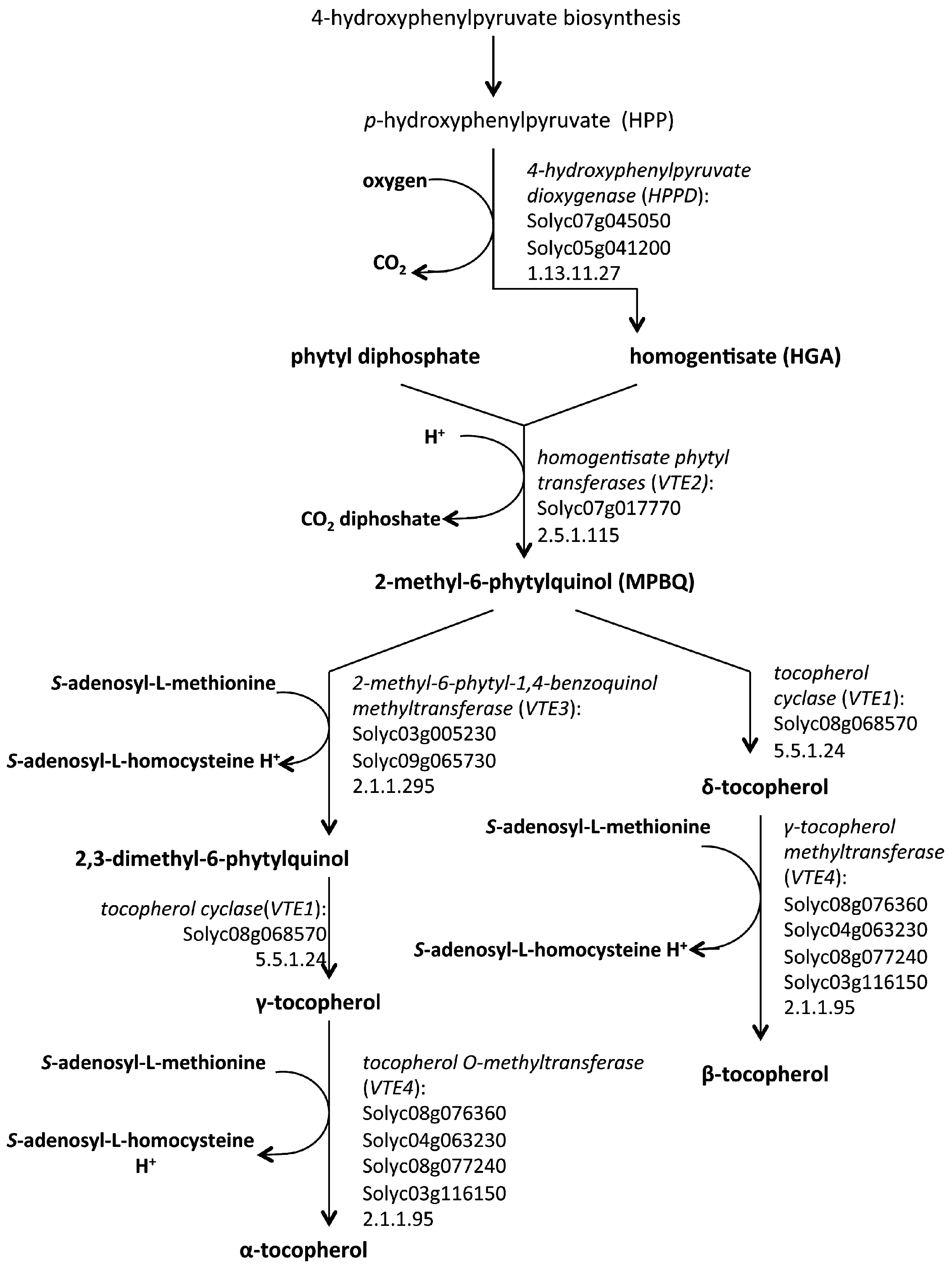
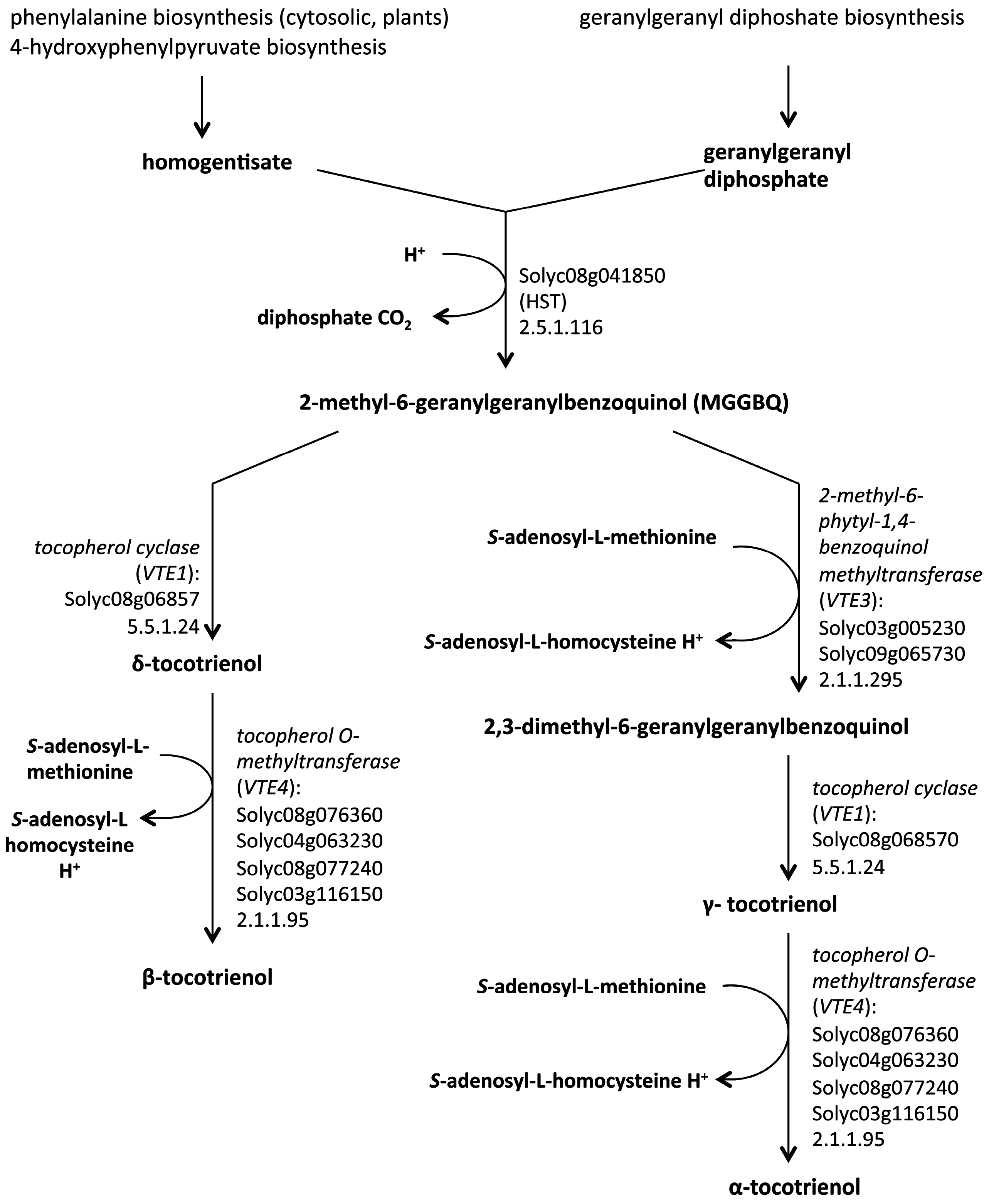
5. Biosynthetic Pathways of Vitamin E


6. Biofortification of Vitamin E Content
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- FAO. Increasing fruit and vegetable consumption becomes a global priority. Available online: http://www.fao.org/english/newsroom/focus/2003/fruitveg1.htm (accessed on 20 October 2015).
- Canene-Adams, K.; Campbell, J.K.; Zaripheh, S.; Jeffery, E.H.; Erdman, J.W., Jr. The tomato as a functional food. J. Nutr. 2005, 135, 1226–1230. [Google Scholar] [PubMed]
- Raiola, A.; Rigano, M.M.; Calafiore, R.; Frusciante, L.; Barone, A. Enhancing the health promoting effects of tomato fruit for biofortified food. Mediat. Inflamm. 2014. [Google Scholar] [CrossRef] [PubMed]
- Traber, M.G. Vitamin E. Adv. Nutr. 2012, 3, 330–331. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q. Natural forms of vitamin E: Metabolism, antioxidant, and anti-inflammatory activities and their role in disease prevention and therapy. Free Radic. Biol. Med. 2014, 72, 76–90. [Google Scholar] [CrossRef] [PubMed]
- Munné-Bosch, S. α-Tocopherol: A multifaceted molecule in plants. Vitam. Horm. 2007, 76, 375–392. [Google Scholar] [PubMed]
- Quadrana, L.; Almeida, J.; Otaiza, S.N.; Duffy, T.; Corrêa da Silva, J.V.; de Godoy, F.; Asís, R.; Bermúdez, L.; Fernie, A.R.; Carrari, F.; et al. Transcriptonal regulation of tocopherol biosynthesis in tomato. Plant Mol. Biol. 2013, 81, 309–325. [Google Scholar] [CrossRef] [PubMed]
- Trebst, A. Function of β-carotene and tocopherol in photosystem II. Z. Naturforsch. C 2003, 58, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Horvath, G.; Wessjohann, L.; Bigirimana, J.; Jansen, M.; Guisez, Y.; Caubergs, R.; Horemans, N. Differential distribution of tocopherols and tocotrienols in photosynthetic and non-photosynthetic tissues. Phytochemistry 2006, 67, 1185–1195. [Google Scholar] [CrossRef] [PubMed]
- Grusak, M.A.; DellaPenna, D. Improving the nutrient composition of plants toenhance human nutrition and health. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 133–161. [Google Scholar] [CrossRef] [PubMed]
- Traber, M.G.; Arai, H. Molecular mechanisms of vitamin E transport. Annu. Rev. Nutr. 1999, 19, 343–355. [Google Scholar] [CrossRef] [PubMed]
- Stampfer, M.J.; Hennekens, C.H.; Manson, J.E.; Colditz, G.A.; Rosner, B.; Willett, W. Vitamin E consumption and the risk of coronary disease in women. N. Engl. J. Med. 1993, 328, 1444–1449. [Google Scholar] [CrossRef] [PubMed]
- Institute of Medicine (IOE). Dietary Reference Intakes for Vitamin C, Vitamin E, Selenium, and Carotenoids; National Academy Press: Washington, DC, USA, 2000. [Google Scholar]
- Zanfini, A.; Corbini, G.; la Rosa, C.; Dreass, E. Antioxidant activity of tomato lipophilic extracts and interactions between carotenoids and α-tocopherol in synthetic mixtures. Food Sci. Technol. 2010, 43, 67–72. [Google Scholar] [CrossRef]
- Raffo, A.; Leonardi, C.; Fogliano, V.; Ambrosino, P.; Salucci, M.; Gennaro, L.; Bugianesi, R.; Giuffrida, F.; Quaglia, G. Nutritional value of cherry tomatoes (Lycopersicon esculentum Cv. Naomi F1) harvested at different ripening stages. J. Agric. Food Chem. 2002, 50, 6550–6556. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, C.; Ambrosino, P.; Esposito, F.; Fogliano, V. Antioxidative activity and carotenoid and tomatine contents in different typologies of fresh consumption tomatoes. J. Agric. Food Chem. 2000, 48, 4723–4727. [Google Scholar] [CrossRef] [PubMed]
- Dumas, Y.; Dadomo, M.; di Lucca, G.; Grolier, P. Effects of environmental factors and agricultural techniques on antioxidant content of tomatoes. J. Sci. Food Agric. 2003, 83, 369–382. [Google Scholar] [CrossRef]
- Abushita, A.A.; Daood, H.G.; Biacs, P.A. Change in carotenoids and antioxidant vitamins in tomato as a function of varietal and technological factors. J. Agric. Food Chem. 2000, 48, 2075–2081. [Google Scholar]
- Rizvi, S.; Raza, S.T.; Ahmed, F.; Ahmad, A.; Abbas, S.; Mahdi, F. The role of vitamin E in human health and some diseases. SQU Med. J. 2014, 14, e158–e165. [Google Scholar]
- Howard, A.C.; Anna, K.; McNeil, A.K.; McNeil, P.L. Promotion of plasma membrane repair by vitamin E. Nat. Commun. 2011, 20, 597. [Google Scholar] [CrossRef] [PubMed]
- Singh, I.; Turner, A.H.; Sinclair, A.J.; Li, D.; Hawley, J.A. Effects of γ-tocopherol supplementation on thrombotic risk factors. Asia Pac. J. Clin. Nutr. 2007, 16, 422–428. [Google Scholar] [PubMed]
- Kirsh, V.A.; Hayes, R.B.; Mayne, S.T.; Chatterjee, N.; Subar, A.F.; Dixon, L.B.; Albanes, D.; Andriole, G.L.; Urban, D.A.; Peters, U. Supplemental and dietary vitamin E, β-carotene, and vitamin C intakes and prostate cancer risk. J. Natl. Cancer Inst. 2006, 98, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Montonen, J.; Knekt, P.; Järvinen, R.; Reunanen, A. Dietary antioxidant intake and risk of type 2 diabetes. Diabetes Care 2004, 27, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Rathore, G.S.; Suthar, M.; Pareek, A.; Gupta, R.N. Nutritional antioxidants: A battle for better health. J. Nat. Pharm. 2011, 2, 2–14. [Google Scholar] [CrossRef]
- Manor, D.; Morley, S. The α-tocopherol transfer protein. Vitam. Horm. 2007, 76, 45–65. [Google Scholar] [PubMed]
- Miller, E.R.; Pastor-Barriuso, R.; Dalal, D.; Riemersma, R.A.; Appel, L.J.; Guallar, E. Meta-analysis: High-dosage vitamin E supplementation may increase all-cause mortality. Ann. Intern. Med. 2005, 142, 37–46. [Google Scholar] [CrossRef] [PubMed]
- National Institute of Health. Vitamin E. 2013. Available online: https://ods.od.nih.gov/factsheets/VitaminE-HealthProfessional/ (accessed on 15 November 2015). [Google Scholar]
- Loffredo, L.; Perri, L.; di Castelnuovo, A.; Iacoviello, L.; de Gaetano, G.; Violi, F. Supplementation with vitamin E alone is associated with reduced myocardial infarction: A meta-analysis. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 354–363. [Google Scholar] [CrossRef] [PubMed]
- Magosso, E.; Ansari, M.A.; Gopalan, Y.; Shuaib, I.L.; Wong, J.W.; Khan, N.A.; Abu Bakar, M.R.; Ng, B.H.; Yuen, K.H. Tocotrienols for normalisation of hepatic echogenic response in nonalcoholic fatty liver: A randomised placebo-controlled clinical trial. Nutr. J. 2013, 12. [Google Scholar] [CrossRef] [PubMed]
- Saboori, S.; Shab-Bidar, S.; Speakman, J.R.; Yousefi Rad, E.; Djafarian, K. Effect of vitamin E supplementation on serum C-reactive protein level: A meta-analysis of randomized controlled trials. Eur. J. Clin. Nutr. 2015, 69, 867–873. [Google Scholar] [CrossRef] [PubMed]
- Upritchard, J.E.; Sutherland, W.H.; Mann, J.I. Effect of supplementation with tomato juice, vitamin E, and vitamin C on LDL oxidation and products of inflammatory activity in type 2 diabetes. Diabetes Care 2000, 23, 733–738. [Google Scholar] [CrossRef] [PubMed]
- Böhm, F.; Edge, R.; Land, E.J.; McGarvey, D.J.; Truscott, T.G. Carotenoids enhance vitamin E antioxidant efficiency. J. Am. Chem. Soc. 1997, 119, 621–622. [Google Scholar] [CrossRef]
- Packer, L. Protective role of vitamin E in biological systems. Am. J. Clin. Nutr. 1991, 53, 1050S–1055S. [Google Scholar] [PubMed]
- Rissanen, T.H.; Voutilainen, S.; Nyyssönen, K.; Salonen, R.; Kaplan, G.A.; Salonen, J.T. Serum lycopene concentrations and carotid atherosclerosis: The Kuopio ischaemic heart disease risk factor study. Am. J. Clin. Nutr. 2003, 77, 133–138. [Google Scholar] [PubMed]
- European Food Safety Authority. Scientific opinion on dietary reference values for vitamin E as α-tocopherol. EFSA J. 2015, 13, 4149. [Google Scholar] [CrossRef] [Green Version]
- Beecher, G.R. Nutrient content of tomatoes and tomato products. Proc. Soc. Exp. Biol. Med. 1998, 218, 98–100. [Google Scholar] [CrossRef] [PubMed]
- Piironen, V.; Syvaoja, E.L.; Varo, P.; Salminen, K.; Koivistoinen, P. Tocopherols and tocotrienols in Finnish foods: Vegetables, fruits, and berries. J. Agric. Food Chem. 1986, 34, 742–746. [Google Scholar] [CrossRef]
- Frusciante, L.; Carli, P.; Ercolano, M.R.; Pernice, R.; di Matteo, A.; Fogliano, V.; Pellegrini, P. Antioxidant nutritional quality of tomato. Mol. Nutr. Food Res. 2007, 51, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Seybold, C.; Fröhlich, K.; Bitsch, R.; Otto, K.; Böhm, V. Changes in contents of carotenoids and vitamin E during tomato processing. J. Agric. Food Chem. 2004, 52, 7005–7010. [Google Scholar] [CrossRef] [PubMed]
- Pék, Z.; Szuvandzsiev, P.; Daood, H.; Neményi, A.; Helyes, L. Effect of irrigation on yield parameters and antioxidant profiles of processing cherry tomato. Cent. Eur. J. Biol. 2014, 9, 383–395. [Google Scholar] [CrossRef]
- Lushchak, V.I. Environmentally induced oxidative stress in aquatic animals. Aquat. Toxicol. 2011, 101, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; DellaPenna, D. Tocopherol functions in photosynthetic organisms. Curr. Opin. Plant Biol. 2007, 10, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.B.; Chu, L.Y.; Lu, Z.H.; Kang, C.M. Primary antioxidant free radical scavenging and redox signaling pathways in higher plant cells. Int. J. Biol. Sci. 2008, 4, 8–14. [Google Scholar] [CrossRef]
- Loyola, J.; Verdugo, I.; González, E.; Casaretto, J.A.; Ruiz-Lara, S. Plastidic isoprenoid biosynthesis in tomato: Physiological andmolecular analysis in genotypes resistant and sensitive to drought stress. Plant Biol. 2012, 14, 149–156. [Google Scholar] [PubMed]
- Skłodowska, M.; Gapińska, M.; Gajewska, E.; Gabara, B. Tocopherol content and enzymatic antioxidant activitiesin chloroplasts from NaCl-stressed tomato plants. Acta Phys. Plant. 2009, 31, 393–400. [Google Scholar] [CrossRef]
- Del Giudice, R.; Raiola, A.; Tenore, G.; Frusciante, L.; Barone, A.; Monti, D.M.; Rigano, M.M. Antioxidant bioactive compounds in tomato fruits at different ripening stages and their effects on normal and cancer cells. J. Funct. Foods 2015, 18, 83–94. [Google Scholar] [CrossRef]
- Caretto, S. Influence of potassium and genotype on vitamin E content and reducing sugar of tomato fruits. Hortscience 2008, 43, 2048–2051. [Google Scholar]
- Lenucci, M.S.; Cadinu, D.; Taurino, M.; Piro, G.; Dalessandro, G. Antioxidant composition in cherry and high-pigment tomato cultivars. J. Agric. Food Chem. 2006, 54, 2606–2613. [Google Scholar] [CrossRef] [PubMed]
- Maršić, N.K.; Sircelj, H.; Kastelec, D. Lipophilic antioxidants and some carpometric characteristics of fruits often processing tomato varieties, grown in different climatic conditions. J. Agric. Food Chem. 2010, 58, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Bramley, P.M.; Rice-Evans, C. Effects of food processing on flavonoids and lycopene status in a Mediterranean tomato variety. Free Radic. Res. 2002, 36, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Van Het Hof, K.; West, C.E.; Weststrate, J.A.; Hautvast, J.G. Dietary factors that affect the bioavailability of carotenoids. J. Nutr. 2000, 130, 503–506. [Google Scholar] [PubMed]
- Capanoglu, E.; Beekwilder, J.; Boyacioglu, D.; Hall, R.; de Vos, R. Changes in antioxidant and metabolite profilesduring production of tomato paste. J. Agric. Food Chem. 2008, 56, 964–973. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.; Lee, J.; Ye, L.; Exler, J.; Eitenmiller, R.R. Tocopherol and tocotrienol contents of raw and processed fruits and vegetables in the United States diet. J. Food Compos. Anal. 2006, 19, 196–204. [Google Scholar] [CrossRef]
- Lavelli, V.; Kerr, W.; Sri Harsha, P.S. Phytochemical stability in dried tomato pulp and peel as affected by moisture properties. J. Agric. Food Chem. 2013, 61, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.S.; Stacewicz-Sapuntzakis, M.; Bowen, P.E. Effects of heat treatment on the carotenoid and tocopherol composition of tomato. J. Food Sci. 2012, 77, C1109–C1114. [Google Scholar] [CrossRef] [PubMed]
- Kalogeropoulos, N.; Chiou, A.; Pyriochou, V.; Peristeraki, A.; Karathanos, V.T. Bioactive phytochemicals in industrial tomatoes and their processing byproducts. Food Sci. Technol. 2012, 49, 213–216. [Google Scholar] [CrossRef]
- Soll, J.; Schultz, G. Comparison of geranylgeranyl and phytylsubstituted methylquinols in the tocopherol synthesis of spinachchloroplasts. Biochem. Biophys. Res. Commun. 1979, 91, 715–720. [Google Scholar] [CrossRef]
- Sun, Q.; Zybailov, B.; Majeran, W.; Friso, G.; Olinares, P.D.B.; van Wijk, K.J. PPDB, the plant proteomics database at Cornell. Nucleic Acids Res. 2009, 37, 969–974. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K. The 1-deoxy-d-xylulose-5-phosphate pathway of isoprenoid biosynthesis in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 47–65. [Google Scholar] [CrossRef] [PubMed]
- DellaPenna, D.; Pogson, B.J. Vitamin synthesis in plants: Tocopherols and carotenoids. Annu. Rev. Plant Biol. 2006, 57, 711–738. [Google Scholar] [CrossRef] [PubMed]
- Caspi, R.; Altman, T.; Billington, R.; Dreher, K.; Foerster, H.; Fulcher, C.A.; Holland, T.A.; Keseler, I.M.; Kothari, A.; Kubo, A.; et al. The MetaCyc database of metabolic pathways and enzymes and the BioCyc collection of Pathway/Genome Databases. Nucleic Acids Res. 2014, 42. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Z.; Sun, X.; Tang, K. The current opinions on the functions of tocopherol based on the genetic manipulation of tocopherol biosynthesis in plants. J. Integr. Plant Biol. 2008, 50, 1057–1069. [Google Scholar] [CrossRef] [PubMed]
- Almeida, L.; Asís, R.; Molineri, V.N.; Sestari, I.; Lira, B.S.; Carrari, F.; Peres, L.E.; Rossi, M. Fruits from ripening impaired, chlorophyll degraded and jasmonate insensitive tomato mutants have altered tocopherol content and composition. Phytochemistry 2015, 111, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Rijzaani, H.; Karcher, D.; Ruf, S.; Bock, R. Efficient metabolic pathway engineering in transgenic tobacco and tomato plastids with synthetic multigene operons. Proc. Natl. Acad. Sci. USA 2013, 4, E623–E632. [Google Scholar] [CrossRef] [PubMed]
- Fraser, P.D.; Enfissi, E.M.; Halket, J.H.; Truesdale, M.R.; Yu, D.M.; Gerrish, C.; Bramley, P.M. Manipulation of phytoene levels intomato fruit: Effects on isoprenoids, plastids and intermediary metabolism. Plant Cell 2007, 19, 3194–3211. [Google Scholar] [CrossRef] [PubMed]
- Perez-Fons, L.; Wells, T.; Corol, D.I.; Ward, J.L.; Gerrish, C.; Beale, M.H.; Seymour, G.B.; Bramley, P.M.; Fraser, P.D. A genome-wide metabolomics resource for tomato fruit from Solanum pennellii. Sci. Rep. 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Eshed, Y.; Zamir, D. An introgression line population of Lycopersicon pennellii in the cultivated tomato enables the identification and fine mapping of yield-associated QTL. Genetics 1995, 141, 1147–1162. [Google Scholar] [PubMed]
- Gur, A.; Zamir, D. Unused natural variation can lift yield barriers in plant breeding. PLoS Biol. 2004, 2, 1610–1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousseaux, M.C.; Jones, C.M.; Adams, D.; Chetelat, R.; Bennett, A.; Powell, A. QTL analysis of fruit antioxidants in tomato using Lycopersicon pennellii introgression lines. Theor. Appl. Genet. 2005, 111, 1396–1408. [Google Scholar] [CrossRef] [PubMed]
- Stevens, R.; Buret, M.; Garchery, C.; Carretero, Y.; Causse, M. Technique for rapid small-scale analysis of vitamin C levels in fruit and application to a tomato mutant collection. J. Agric. Food Chem. 2006, 54, 6159–6165. [Google Scholar] [CrossRef] [PubMed]
- Schauer, N.; Semel, Y.; Roessner, U.; Gur, A.; Balbo, I.; Carrari, F.; Pleban, T.; Perez-Melis, A.; Bruedigam, C.; Kopka, J.; et al. Comprehensive metabolic profiling and phenotyping of interspecific introgression lines for tomato improvement. Nat. Biotechnol. 2006, 24, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.S.; Kim, S.J.; Harn, C.H.; Kim, W.T. Ectopic expression of apple fruit homogentisate phytyltransferase gene (MdHPT1) increases tocopherol in transgenic tomato (Solanum lycopersicum Cv. Micro-Tom) leaves and fruits. Phytochemistry 2011, 72, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Rigano, M.M.; Scotti, N.; Cardi, T. Unsolved problems in plastid transformation. Bioengineered 2012, 3, 329–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raiola, A.; Tenore, G.C.; Barone, A.; Frusciante, L.; Rigano, M.M. Vitamin E Content and Composition in Tomato Fruits: Beneficial Roles and Bio-Fortification. Int. J. Mol. Sci. 2015, 16, 29250-29264. https://doi.org/10.3390/ijms161226163
Raiola A, Tenore GC, Barone A, Frusciante L, Rigano MM. Vitamin E Content and Composition in Tomato Fruits: Beneficial Roles and Bio-Fortification. International Journal of Molecular Sciences. 2015; 16(12):29250-29264. https://doi.org/10.3390/ijms161226163
Chicago/Turabian StyleRaiola, Assunta, Gian Carlo Tenore, Amalia Barone, Luigi Frusciante, and Maria Manuela Rigano. 2015. "Vitamin E Content and Composition in Tomato Fruits: Beneficial Roles and Bio-Fortification" International Journal of Molecular Sciences 16, no. 12: 29250-29264. https://doi.org/10.3390/ijms161226163
APA StyleRaiola, A., Tenore, G. C., Barone, A., Frusciante, L., & Rigano, M. M. (2015). Vitamin E Content and Composition in Tomato Fruits: Beneficial Roles and Bio-Fortification. International Journal of Molecular Sciences, 16(12), 29250-29264. https://doi.org/10.3390/ijms161226163