
Coupling and Coordination in Gene Expression Processes with Pre-mRNA Splicing

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
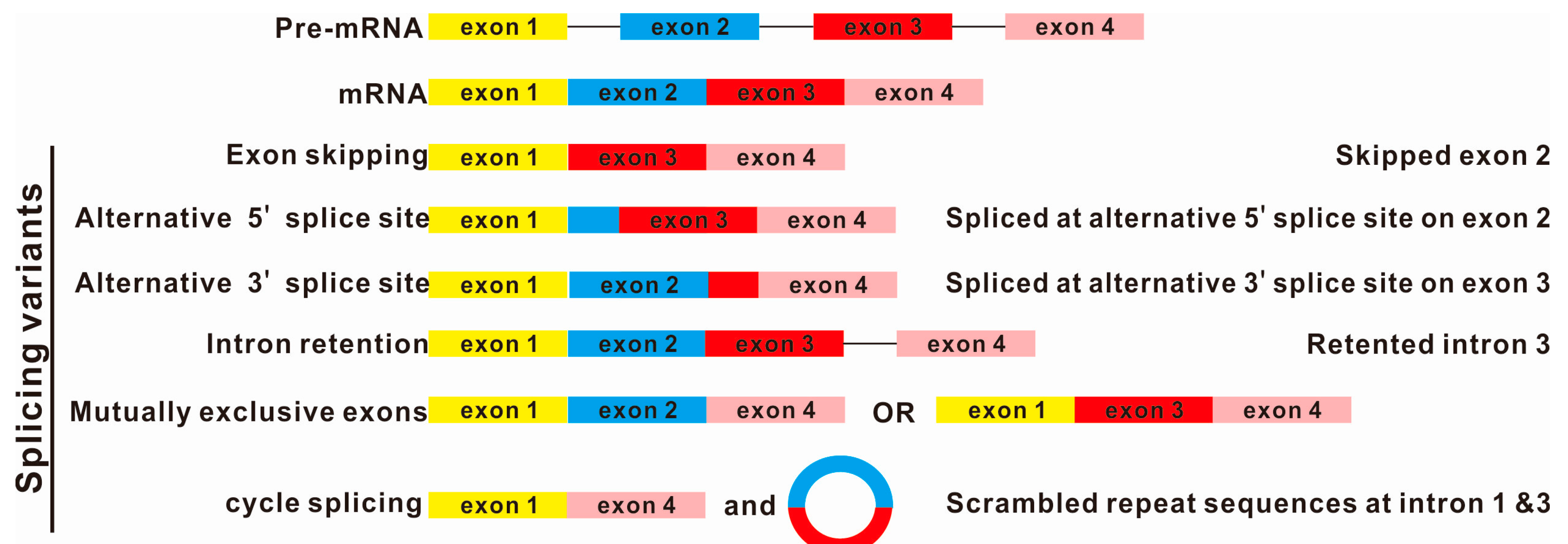
:1. Introduction


2. Splicing and Nuclear RNA Surveillance
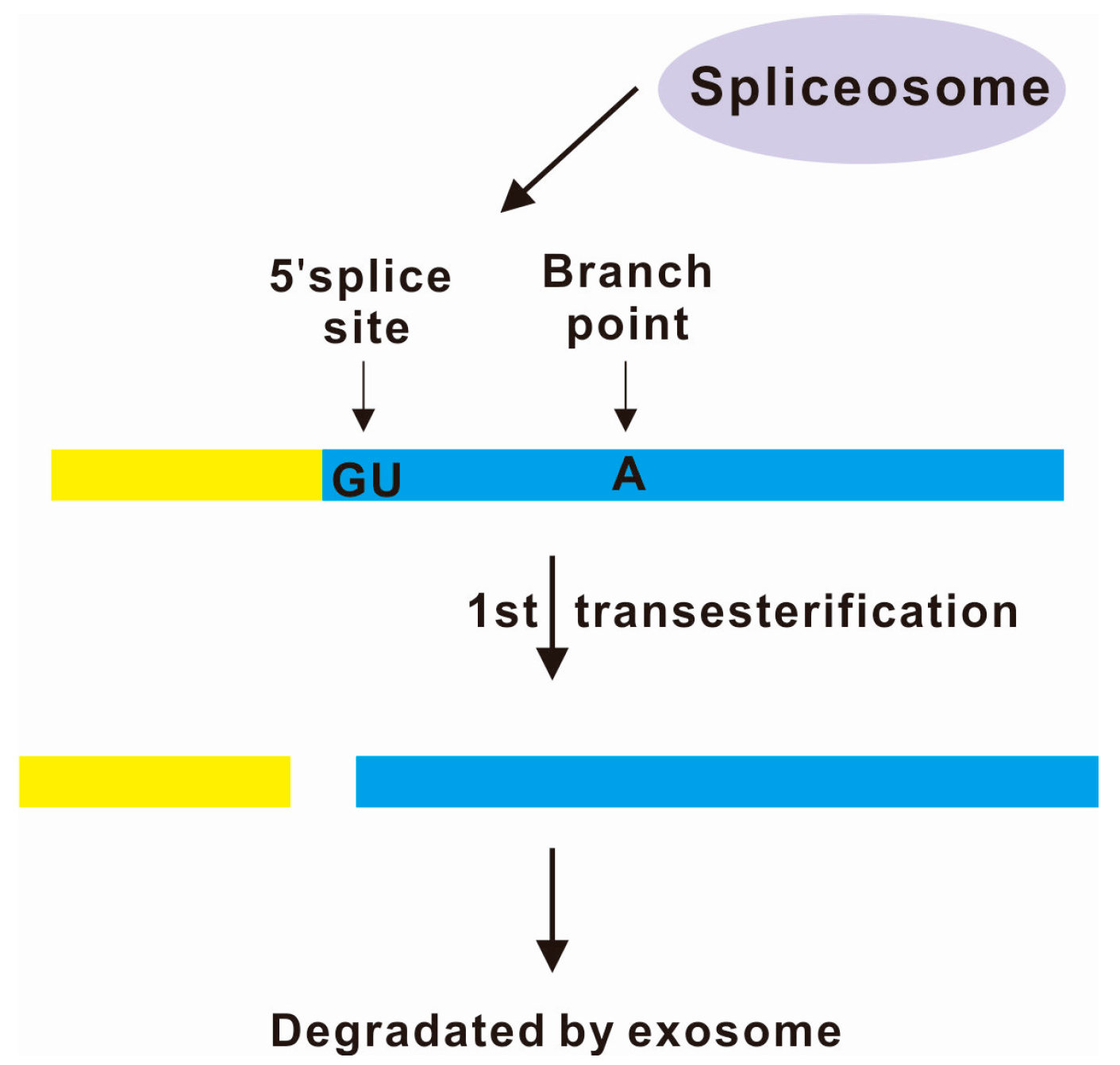
3. Spliceosome-Mediated Decay

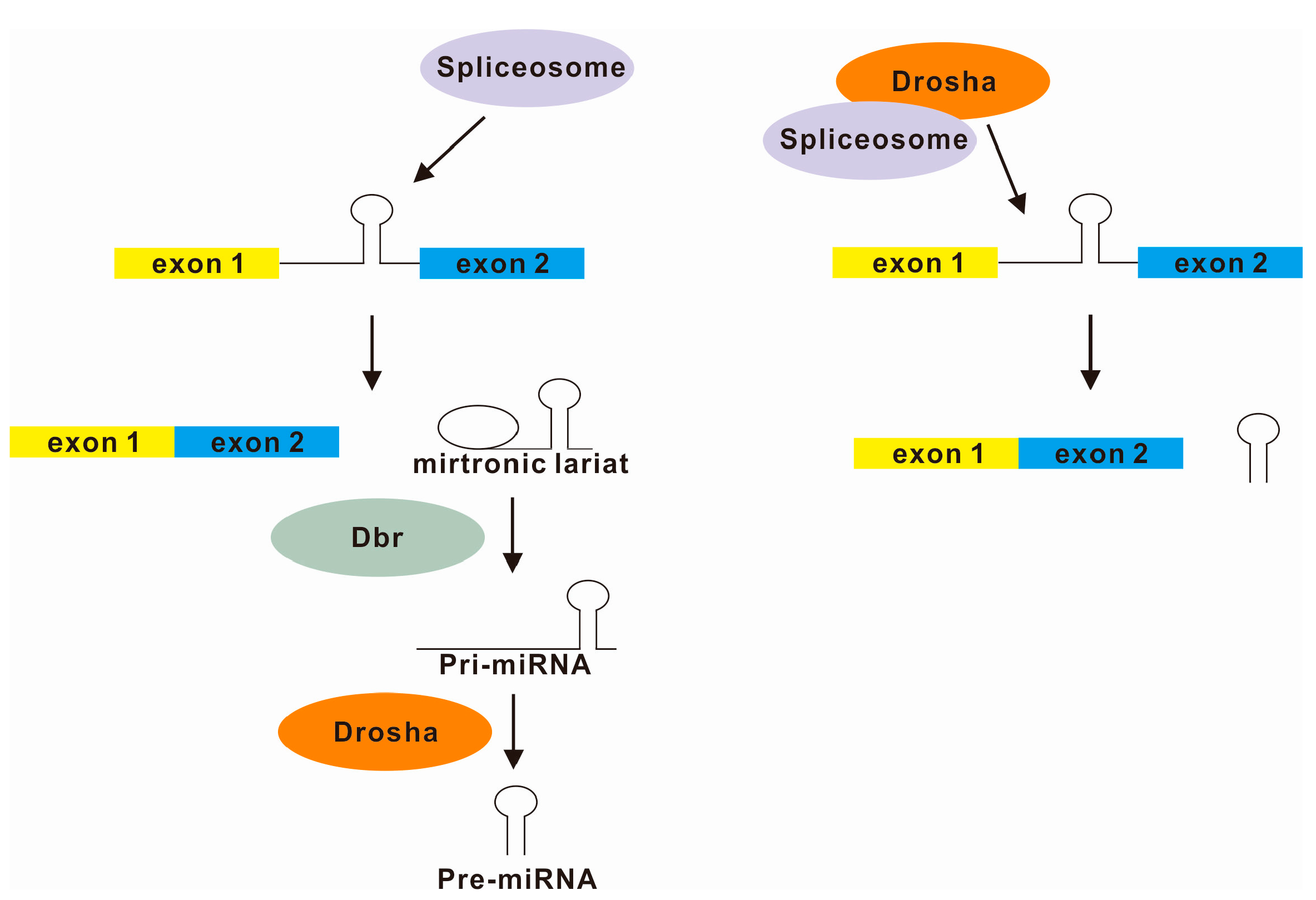
4. Splicing and microRNA Processing

5. Splicing and Circular RNAs
6. Conclusions and Perspectives
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Neverov, A.D.; Artamonova, II; Nurtdinov, R.N.; Frishman, D.; Gelfand, M.S.; Mironov, A.A. Alternative splicing and protein function. BMC Bioinform. 2005, 6, 266. [Google Scholar]
- Wu, J.Y.; Tang, H.; Havlioglu, N. Alternative pre-mRNA splicing and regulation of programmed cell death. Regul. Altern. Splicing 2003, 31, 153–185. [Google Scholar]
- Rino, J.; Carvalho, T.; Braga, J.; Desterro, J.M.; Luhrmann, R.; Carmo-Fonseca, M. A stochastic view of spliceosome assembly and recycling in the nucleus. PLoS Comput. Biol. 2007, 3, 2019–2031. [Google Scholar] [CrossRef] [PubMed]
- Montemayor, E.J.; Katolik, A.; Clark, N.E.; Taylor, A.B.; Schuermann, J.P.; Combs, D.J.; Johnsson, R.; Holloway, S.P.; Stevens, S.W.; Damha, M.J.; et al. Structural basis of lariat RNA recognition by the intron debranching enzyme Dbr1. Nucleic Acids Res. 2014, 42, 10845–10855. [Google Scholar] [CrossRef] [PubMed]
- Armakola, M.; Higgins, M.J.; Figley, M.D.; Barmada, S.J.; Scarborough, E.A.; et al. Inhibition of RNA lariat debranching enzyme suppresses TDP-43 toxicity in ALS disease models. Nat. Genet. 2012, 44, 1302–1309. [Google Scholar] [CrossRef] [PubMed]
- Hesselberth, J.R. Lives that introns lead after splicing. Wiley Interdiscip. Rev. RNA 2013, 4, 677–691. [Google Scholar] [PubMed]
- Baralle, D.; Baralle, M. Splicing in action: Assessing disease causing sequence changes. J. Med. Genet. 2005, 42, 737–748. [Google Scholar] [CrossRef] [PubMed]
- Chasin, L.A. Searching for splicing motifs. Adv. Exp. Med. Biol. 2007, 623, 85–106. [Google Scholar] [PubMed]
- Fairbrother, W.G.; Yeh, R.F.; Sharp, P.A.; Burge, C.B. Predictive identification of exonic splicing enhancers in human genes. Science 2002, 297, 1007–1013. [Google Scholar] [CrossRef] [PubMed]
- Krawczak, M.; Reiss, J.; Cooper, D.N. The mutational spectrum of single base-pair substitutions in mRNA splice junctions of human genes: Causes and consequences. Hum. Genet. 1992, 90, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Cartegni, L.; Chew, S.L.; Krainer, A.R. Listening to silence and understanding nonsense: Exonic mutations that affect splicing. Nat. Rev. Genet. 2002, 3, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.H.; Ferraris, L.; Filloux, M.E.; Raphael, B.J.; Fairbrother, W.G. Using positional distribution to identify splicing elements and predict pre-mRNA processing defects in human genes. Proc. Natl. Acad. Sci. USA 2011, 108, 11093–11098. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, J.; Li, K.; Zhao, W.; Cui, Q. SpliceDisease database: Linking RNA splicing and disease. Nucleic Acids Res. 2012, 40, D1055–D1059. [Google Scholar] [CrossRef] [PubMed]
- Kapustin, Y.; Chan, E.; Sarkar, R.; Wong, F.; Vorechovsky, I.; Winston, R.M.; Tatusova, T.; Dibb, N.J. Cryptic splice sites and split genes. Nucleic Acids Res. 2011, 39, 5837–5844. [Google Scholar] [CrossRef] [PubMed]
- Faustino, N.A.; Cooper, T.A. Pre-mRNA splicing and human disease. Genes Dev. 2003, 17, 419–437. [Google Scholar] [CrossRef] [PubMed]
- Hui, J. Regulation of mammalian pre-mRNA splicing. Sci. China Ser. C Life Sci. 2009, 52, 253–260. [Google Scholar] [CrossRef]
- Iborra, S.; Hirschfeld, M.; Jaeger, M.; Zur Hausen, A.; Braicu, I.; Sehouli, J.; Gitsch, G.; Stickeler, E. Alterations in expression pattern of splicing factors in epithelial ovarian cancer and its clinical impact. Int. J. Gynecol. Cancer 2014, 23, 990–996. [Google Scholar] [CrossRef]
- Das, S.; Krainer, A.R. Emerging functions of SRSF1, splicing factor and oncoprotein, in RNA metabolism and cancer. Mol. Cancer Res. 2014, 12, 1195–1204. [Google Scholar] [CrossRef] [PubMed]
- Hickey, C.J.; Kim, J.H.; Ahn, E.Y. New discoveries of old SON: A link between RNA splicing and cancer. J. Cell Biochem. 2014, 115, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Vaz-Drago, R.; Pinheiro, M.T.; Martins, S.; Enguita, F.; Carmo-Fonseca, M.; Custódio, N. Transcription-coupled RNA surveillance in human genetic diseases caused by splice site mutations. Hum. Mol. Genet. 2015. [Google Scholar] [CrossRef]
- Wood, M.J.; Gait, M.J.; Yin, H. RNA-targeted splice-correction therapy for neuromuscular disease. Brain 2010, 133, 957–972. [Google Scholar] [CrossRef] [PubMed]
- Kole, R.; Leppert, B.J. Targeting mRNA splicing as a potential treatment for duchenne muscular dystrophy. Discov. Med. 2012, 14, 59–69. [Google Scholar] [PubMed]
- Kole, R.; Krainer, A.R.; Altman, S. RNA therapeutics: Beyond RNA interference and antisense oligonucleotides. Nat. Rev. Drug Discov. 2012, 11, 125–140. [Google Scholar] [PubMed]
- Ward, A.J.; Cooper, T.A. The pathobiology of splicing. J. Pathol. 2010, 220, 152–163. [Google Scholar] [PubMed]
- Cooper, T.A.; Wan, L.; Dreyfuss, G. RNA and disease. Cell 2009, 136, 777–793. [Google Scholar] [CrossRef] [PubMed]
- Wahl, M.C.; Will, C.L.; Luhrmann, R. The spliceosome: Design principles of a dynamic RNP machine. Cell 2009, 136, 701–718. [Google Scholar] [CrossRef] [PubMed]
- Keren, H.; Lev-Maor, G.; Ast, G. Alternative splicing and evolution: Diversification, exon definition and function. Nat. Rev. Genet. 2010, 11, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Black, D.L. Mechanisms of alternative pre-messenger RNA splicing. Annu. Rev. Biochem. 2003, 72, 291–336. [Google Scholar] [CrossRef] [PubMed]
- Stoilov, P.; Daoud, R.; Nayler, O.; Stamm, S. Human tra2-β1 autoregulates its protein concentration by influencing alternative splicing of its pre-mRNA. Hum. Mol. Genet. 2004, 13, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.T.; Sandberg, R.; Luo, S.; Khrebtukova, I.; Zhang, L.; Mayr, C.; Kingsmore, S.F.; Schroth, G.P.; Burge, C.B. Alternative isoform regulation in human tissue transcriptomes. Nature 2008, 456, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.M.; Castle, J.; Garrett-Engele, P.; Kan, Z.; Loerch, P.M.; Armour, C.D.; Santos, R.; Schadt, E.E.; Stoughton, R.; Shoemaker, D.D. Genome-wide survey of human alternative pre-mRNA splicing with exon junction microarrays. Science 2003, 302, 2141–2144. [Google Scholar] [CrossRef] [PubMed]
- Salzman, J.; Gawad, C.; Wang, P.L.; Lacayo, N.; Brown, P.O. Circular RNAs are the predominant transcript isoform from hundreds of human genes in diverse cell types. PLoS One 2012, 7, e30733. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.Y.; Liu, H.J.; Hung, L.Y.; Kuo, H.C.; Chuang, T.J. Is an observed non-co-linear RNA product spliced in trans, in cis or just in vitro? Nucleic Acids Res. 2014, 42, 9410–9423. [Google Scholar] [CrossRef] [PubMed]
- Valdmanis, P.N.; Kay, M.A. The expanding repertoire of circular RNAs. Mol. Ther. 2013, 21, 1112–1114. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.O.; Wang, H.B.; Zhang, Y.; Lu, X.; Chen, L.L.; Yang, L. Complementary sequence-mediated exon circularization. Cell 2014, 159, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Vicens, Q.; Westhof, E. Biogenesis of circular RNAs. Cell 2014, 159, 13–14. [Google Scholar] [CrossRef] [PubMed]
- Baejen, C.; Torkler, P.; Gressel, S.; Essig, K.; Soding, J.; Cramer, P. Transcriptome maps of mRNP biogenesis factors define pre-mRNA recognition. Mol. Cell 2014, 55, 745–757. [Google Scholar] [CrossRef] [PubMed]
- Delan-Forino, C.; Tollervey, D. Lighting up pre-mRNA recognition. Mol. Cell 2014, 55, 649–651. [Google Scholar] [CrossRef] [PubMed]
- Koodathingal, P.; Staley, J.P. Splicing fidelity: DEAD/H-box ATPases as molecular clocks. RNA Biol 2013, 10, 1073–1079. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Wang, X.Y.; Zhang, ZM.; Pu, J.; Fan, Y.J.; Zhou, J.; Query, C.C.; Xu, Y.Z. Splicing proofreading at 5' splice sites by ATPase Prp28p. Nucleic Acids Res. 2013, 41, 4660–4670. [Google Scholar] [CrossRef] [PubMed]
- Bonde, M.M.; Voegeli, S.; Baudrimont, A.; Seraphin, B.; Becskei, A. Quantification of pre-mRNA escape rate and synergy in splicing. Nucleic Acids Res. 2014, 42, 12847–12860. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, T.; Pellegrini, M.; Chanfreau, G.F. Nonsense-mediated mRNA decay mutes the splicing defects of spliceosome component mutations. RNA 2009, 15, 2236–2247. [Google Scholar] [CrossRef] [PubMed]
- Eberle, A.B.; Hessle, V.; Helbig, R.; Dantoft, W.; Gimber, N.; Visa, N. Splice-site mutations cause Rrp6-mediated nuclear retention of the unspliced RNAs and transcriptional down-regulation of the splicing-defective genes. PLoS One 2010, 5, e11540. [Google Scholar] [CrossRef] [PubMed]
- Niemela, E.H.; Oghabian, A.; Staals, RH.; Greco, D.; Pruijn, G.J.; Frilander, M.J. Global analysis of the nuclear processing of transcripts with unspliced U12-type introns by the exosome. Nucleic Acids Res. 2014, 42, 7358–7369. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, T.; Douglass, S.; Gabunilas, J.; Pellegrini, M.; Chanfreau, G.F. Widespread use of non-productive alternative splice sites in Saccharomyces cerevisiae. PLoS Genet. 2014, 10, e1004249. [Google Scholar] [CrossRef] [PubMed]
- Chanfreau, G.F. A dual role for RNA splicing signals. EMBO Rep 2010, 11, 720–721. [Google Scholar] [CrossRef] [PubMed]
- Davidson, L.; Kerr, A.; West, S. Co-transcriptional degradation of aberrant pre-mRNA by Xrn2. EMBO J. 2012, 31, 2566–2578. [Google Scholar] [CrossRef] [PubMed]
- Egecioglu, D.E.; Kawashima, T.R.; Chanfreau, G.F. Quality control of MATa1 splicing and exon skipping by nuclear RNA degradation. Nucleic Acids Res. 2012, 40, 1787–1796. [Google Scholar] [CrossRef] [PubMed]
- Isken, O.; Maquat, L.E. Quality control of eukaryotic mRNA: Safeguarding cells from abnormal mRNA function. Genes Dev. 2007, 21, 1833–1856. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.F.; Imam, J.S.; Wilkinson, M.F. The nonsense-mediated decay RNA surveillance pathway. Annu. Rev. Biochem. 2007, 76, 51–74. [Google Scholar] [CrossRef] [PubMed]
- Shyu, A.B.; Wilkinson, M.F.; van Hoof, A. Messenger RNA regulation: To translate or to degrade. EMBO J. 2008, 27, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Galy, V.; Gadal, O.; Fromont-Racine, M.; Romano, A.; Jacquier, A.; Nehrbass, U. Nuclear retention of unspliced mRNAs in yeast is mediated by perinuclear Mlp1. Cell 2004, 116, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Gencheva, M.; Lin, T.Y.; Wu, X.; Yang, L.; Richard, C.; Jones, M.; Lin, S.B.; Lin, R.J. Nuclear retention of unspliced pre-mRNAs by mutant DHX16/hPRP2, a spliceosomal DEAH-box protein. J. Biol. Chem. 2010, 285, 35624–35632. [Google Scholar] [CrossRef] [PubMed]
- Wery, M.; Ruidant, S.; Schillewaert, S.; Lepore, N.; Lafontaine, D.L. The nuclear poly(A) polymerase and Exosome cofactor Trf5 is recruited cotranscriptionally to nucleolar surveillance. RNA 2009, 15, 406–419. [Google Scholar] [CrossRef] [PubMed]
- Vasiljeva, L.; Kim, M.; Terzi, N.; Soares, L.M.; Buratowski, S. Transcription termination and RNA degradation contribute to silencing of RNA polymerase II transcription within heterochromatin. Mol. Cell 2008, 29, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Schmid, M.; Jensen, T.H. Quality control of mRNP in the nucleus. Chromosoma 2008, 117, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Kress, T.L.; Krogan, N.J.; Guthrie, C. A single SR-like protein, Npl3, promotes pre-mRNA splicing in budding yeast. Mol. Cell 2008, 32, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Milliman, E.J.; Goulet, I.; Cote, J.; Jackson, C.A.; Vollbracht, J.A.; Yu, M.C. Protein arginine methylation facilitates cotranscriptional recruitment of pre-mRNA splicing factors. Mol. Cell Biol. 2010, 30, 5245–5256. [Google Scholar] [CrossRef] [PubMed]
- Lenasi, T.; Peterlin, B.M.; Barboric, M. Cap-binding protein complex links pre-mRNA capping to transcription elongation and alternative splicing through positive transcription elongation factor b (P-TEFb). J. Biol. Chem. 2011, 286, 22758–22768. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Mayoral, M.F.; Hollingworth, D.; Masino, L.; Diaz-Moreno, I.; Kelly, G.; Gherzi, R.; Chou, C.F.; Chen, C.Y.; Ramos, A. The structure of the C-terminal KH domains of KSRP reveals a noncanonical motif important for mRNA degradation. Structure 2007, 15, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Golisz, A.; Sikorski, P.J.; Kruszka, K.; Kufel, J. Arabidopsis thaliana LSM proteins function in mRNA splicing and degradation. Nucleic Acids Res. 2013, 41, 6232–6249. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.M.; Qiu, H.; Hu, C.; Dong, J.; Hinnebusch, A.G. Yeast cap binding complex impedes recruitment of cleavage factor IA to weak termination sites. Mol. Cell Biol. 2007, 27, 6520–6531. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.M.; Tang, H.M.; Kong, K.Y.; Wong, G.W.; Qiu, H.; Jin, D.Y.; Hinnebusch, A.G. Yeast arginine methyltransferase Hmt1p regulates transcription elongation and termination by methylating Npl3p. Nucleic Acids Res. 2010, 38, 2217–2228. [Google Scholar] [CrossRef] [PubMed]
- Hessle, V.; von Euler, A.; Gonzalez de Valdivia, E.; Visa, N. Rrp6 is recruited to transcribed genes and accompanies the spliced mRNA to the nuclear pore. RNA 2012, 18, 1466–1474. [Google Scholar] [CrossRef] [PubMed]
- Houseley, J.; Tollervey, D. The nuclear RNA surveillance machinery: The link between ncRNAs and genome structure in budding yeast? Biochim. Biophys. Acta 2008, 1779, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Synowsky, S.A.; van Wijk, M.; Raijmakers, R.; Heck, A.J. Comparative multiplexed mass spectrometric analyses of endogenously expressed yeast nuclear and cytoplasmic exosomes. J. Mol. Biol. 2009, 385, 1300–1313. [Google Scholar] [CrossRef] [PubMed]
- Stutz, F.; Izaurralde, E. The interplay of nuclear mRNP assembly, mRNA surveillance and export. Trends Cell Biol. 2003, 13, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Vasudevan, S.; Peltz, S.W. Nuclear mRNA surveillance. Curr. Opin. Cell Biol. 2003, 15, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, K.; Butler, J.S. Nuclear RNA surveillance: role of TRAMP in controlling exosome specificity. Wiley Interdiscip. Rev. RNA 2013, 4, 217–231. [Google Scholar] [CrossRef] [PubMed]
- LaCava, J.; Houseley, J.; Saveanu, C.; Petfalski, E.; Thompson, E.; Jacquier, A.; Tollervey, D. RNA degradation by the exosome is promoted by a nuclear polyadenylation complex. Cell 2005, 121, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Kong, K.Y.; Tang, H.M.; Pan, K.; Huang, Z.; Lee, T.H.; Hinnebusch, A.G.; Jin, D.Y.; Wong, C.M. Cotranscriptional recruitment of yeast TRAMP complex to intronic sequences promotes optimal pre-mRNA splicing. Nucleic Acids Res. 2014, 42, 643–660. [Google Scholar] [CrossRef] [PubMed]
- San Paolo, S.; Vanacova, S.; Schenk, L.; Scherrer, T.; Blank, D.; Keller, W.; Gerber, A.P. Distinct roles of non-canonical poly(A) polymerases in RNA metabolism. PLoS Genet. 2009, 5, e1000555. [Google Scholar]
- Nag, A.; Steitz, J.A. Tri-snRNP-associated proteins interact with subunits of the TRAMP and nuclear exosome complexes, linking RNA decay and pre-mRNA splicing. RNA Biol. 2012, 9, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Hackmann, A.; Wu, H.; Schneider, U.M.; Meyer, K.; Jung, K.; Krebber, H. Quality control of spliced mRNAs requires the shuttling SR proteins Gbp2 and Hrb1. Nat. Commun. 2014, 5, 3123. [Google Scholar] [CrossRef] [PubMed]
- Volanakis, A.; Passoni, M.; Hector, R.D.; Shah, S.; Kilchert, C.; Granneman, S.; Vasiljeva, L. Spliceosome-mediated decay (SMD) regulates expression of nonintronic genes in budding yeast. Genes Dev. 2013, 27, 2025–2038. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Hou, J.; Liu, L.; Chen, L.; Wang, M.; Shen, Y.; Zhang, Z.; Bao, X. Interplay between BDF1 and BDF2 and their roles in regulating the yeast salt stress response. FEBS J. 2013, 280, 1991–2001. [Google Scholar] [CrossRef] [PubMed]
- Alexandrov, A.; Colognori, D.; Shu, M.D.; Steitz, J.A. Human spliceosomal protein CWC22 plays a role in coupling splicing to exon junction complex deposition and nonsense-mediated decay. Proc. Natl. Acad. Sci. USA 2012, 109, 21313–21318. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.K.; Kim, V.N. Processing of intronic microRNAs. EMBO J. 2007, 26, 775–783. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.; Griffiths-Jones, S.; Ashurst, J.L.; Bradley, A. Identification of mammalian microRNA host genes and transcription units. Genome Res. 2004, 14, 1902–1910. [Google Scholar] [CrossRef] [PubMed]
- Westholm, J.O.; Lai, E.C. Mirtrons: MicroRNA biogenesis via splicing. Biochimie 2011, 93, 1897–1904. [Google Scholar] [CrossRef] [PubMed]
- Melamed, Z.; Levy, A.; Ashwal-Fluss, R.; Lev-Maor, G.; Mekahel, K.; Atias, N.; Gilad, S.; Sharan, R.; Levy, C.; Kadener, S.; et al. Alternative splicing regulates biogenesis of miRNAs located across exon-intron junctions. Mol. Cell 2013, 50, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Mattioli, C.; Pianigiani, G.; Pagani, F. Cross talk between spliceosome and microprocessor defines the fate of pre-mRNA. Wiley Interdiscip. Rev. RNA 2014, 5, 647–658. [Google Scholar] [PubMed]
- Kataoka, N.; Fujita, M.; Ohno, M. Functional association of the Microprocessor complex with the spliceosome. Mol. Cell Biol. 2009, 29, 3243–3254. [Google Scholar] [CrossRef] [PubMed]
- Shomron, N.; Levy, C. MicroRNA-biogenesis and pre-mRNA splicing crosstalk. J. Biomed. Biotechnol. 2009, 2009, 594678. [Google Scholar] [PubMed]
- Agranat-Tamir, L.; Shomron, N.; Sperling, J.; Sperling, R. Interplay between pre-mRNA splicing and microRNA biogenesis within the supraspliceosome. Nucleic Acids Res. 2014, 42, 4640–4651. [Google Scholar] [CrossRef] [PubMed]
- Szweykowska-Kulinska, Z.; Jarmolowski, A.; Vazquez, F. The crosstalk between plant microRNA biogenesis factors and the spliceosome. Plant Signal. Behav. 2013, 8, e26955. [Google Scholar] [CrossRef] [PubMed]
- Shefer, K.; Sperling, J.; Sperling, R. The Supraspliceosome—A multi-task machine for regulated pre-mRNA processing in the cell nucleus. Comput. Struct. Biotechnol. J. 2014, 11, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Guil, S.; Caceres, J.F. The multifunctional RNA-binding protein hnRNP A1 is required for processing of miR-18a. Nat. Struct. Mol. Biol. 2007, 14, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Sun, S.; Tu, K.; Gao, Y.; Xie, B.; Krainer, A.R.; Zhu, J. A splicing-independent function of SF2/ASF in microRNA processing. Mol. Cell 2010, 38, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Michlewski, G.; Caceres, J.F. Antagonistic role of hnRNP A1 and KSRP in the regulation of let-7a biogenesis. Nat. Struct. Mol. Biol. 2010, 17, 1011–1018. [Google Scholar] [CrossRef] [PubMed]
- Trabucchi, M.; Briata, P.; Garcia-Mayoral, M.; Haase, A.D.; Filipowicz, W.; Ramos, A.; Gherzi, R.; Rosenfeld, M.G. The RNA-binding protein KSRP promotes the biogenesis of a subset of microRNAs. Nature 2009, 459, 1010–1014. [Google Scholar] [CrossRef] [PubMed]
- Janas, M.M.; Khaled, M.; Schubert, S.; Bernstein, J.G.; Golan, D.; Veguilla, R.A.; Fisher, D.E.; Shomron, N.; Levy, C.; Novina, C.D. Feed-forward microprocessing and splicing activities at a microRNA-containing intron. PLoS Genet. 2011, 7, e1002330. [Google Scholar] [CrossRef] [PubMed]
- Ruby, J.G.; Jan, C.H.; Bartel, D.P. Intronic microRNA precursors that bypass Drosha processing. Nature 2007, 448, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Havens, M.A.; Reich, A.A.; Hastings, M.L. Drosha promotes splicing of a pre-microRNA-like alternative exon. PLoS Genet. 2014, 10, e1004312. [Google Scholar] [CrossRef] [PubMed]
- Mattioli, C.; Pianigiani, G.; Pagani, F. A competitive regulatory mechanism discriminates between juxtaposed splice sites and pri-miRNA structures. Nucleic Acids Res. 2013, 1, 8680–8691. [Google Scholar] [CrossRef]
- Ashwal-Fluss, R.; Meyer, M.; Pamudurti, N.R.; Ivanov, A.; Bartok, O.; Hanan, M.; Evantal, N.; Memczak, S.; Rajewsky, N.; Kadener, S. circRNA biogenesis competes with pre-mRNA splicing. Mol. Cell 2014, 56, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Wilusz, J.E.; Sharp, P.A. Molecular biology. A circuitous route to noncoding RNA. Science 2013, 340, 440–441. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Z. Efficient backsplicing produces translatable circular mRNAs. RNA 2015, 21, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Lasda, E.; Parker, R. Circular RNAs: Diversity of form and function. RNA 2014, 20, 1829–1842. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Chen, L.L. Competition of RNA splicing: Line in or circle up. Sci. China Life Sci. 2014, 57, 1232–1233. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Wilusz, J.E. Short intronic repeat sequences facilitate circular RNA production. Genes Dev. 2014, 28, 2233–2247. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, X.O.; Chen, T.; Xiang, J.F.; Yin, Q.F.; Xing, Y.H.; Zhu, S.; Yang, L.; Chen, L.L. Circular intronic long noncoding RNAs. Mol. Cell 2013, 51, 792–806. [Google Scholar] [CrossRef] [PubMed]
- Jeck, W.R.; Sharpless, N.E. Detecting and characterizing circular RNAs. Nat. Biotechnol. 2014, 32, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Ghosal, S.; Das, S.; Sen, R.; Basak, P.; Chakrabarti, J. Circ2Traits: A comprehensive database for circular RNA potentially associated with disease and traits. Front. Genet. 2013, 4, 283. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.L.; Bao, Y.; Yee, M.C.; Barrett, S.P.; Hogan, G.J.; Olsen, M.N.; Dinneny, J.R.; Brown, P.O.; Salzman, J. Circular RNA is expressed across the eukaryotic tree of life. PLoS One 2014, 9, e90859. [Google Scholar] [CrossRef] [PubMed]
- Starke, S.; Jost, I.; Rossbach, O.; Schneider, T.; Schreiner, S.; Hung, L.H.; Bindereif, A. Exon circularization requires canonical splice signals. Cell Rep. 2015, 10, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Tsukahara, T. A view of pre-mRNA splicing from RNase R resistant RNAs. Int. J. Mol. Sci. 2014, 15, 9331–9342. [Google Scholar] [CrossRef] [PubMed]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA 2013, 19, 141–157. [Google Scholar] [CrossRef] [PubMed]
- Westholm, J.O.; Miura, P.; Olson, S.; Shenker, S.; Joseph, B.; Sanfilippo, P.; Celniker, S.E.; Graveley, B.R.; Lai, E.C. Genome-wide analysis of Drosophila circular RNAs reveals their structural and sequence properties and age-dependent neural accumulation. Cell Rep. 2014, 9, 1966–1980. [Google Scholar] [CrossRef] [PubMed]
- Lukiw, W.J. Circular RNA (circRNA) in Alzheimer’s disease (AD). Front. Genet. 2013, 4, 307. [Google Scholar] [PubMed]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Bak, R.O.; Hollensen, A.K.; Mikkelsen, J.G. Managing microRNAs with vector-encoded decoy-type inhibitors. Mol. Ther. 2013, 21, 1478–1485. [Google Scholar] [CrossRef] [PubMed]
- Ozsolak, F.; Milos, P.M. RNA sequencing: Advances, challenges and opportunities. Nat. Rev. Genet. 2011, 12, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Sorenson, M.R.; Stevens, S.W. Rapid identification of mRNA processing defects with a novel single-cell yeast reporter. RNA 2014, 20, 732–745. [Google Scholar] [CrossRef] [PubMed]
- Marinov, G.K.; Williams, B.A.; McCue, K.; Schroth, G.P.; Gertz, J.; Myers, R.M.; Wold, B.J. From single-cell tocell-pool transcriptomes: Stochasticity in gene expression and RNA splicing. Genome Res. 2014, 24, 496–510. [Google Scholar] [CrossRef] [PubMed]
- Schamberger, A.; Orban, T.I. Experimental validation of predicted mammalian microRNAs of mirtron origin. Methods Mol. Biol. 2014, 1182, 245–263. [Google Scholar] [PubMed]
- Wang, Z.; Rolish, M.E.; Yeo, G.; Tung, V.; Mawson, M.; Burge, C.B. Systematic identification and analysis of exonic splicing silencers. Cell 2004, 119, 831–845. [Google Scholar] [CrossRef] [PubMed]
- Barash, Y.; Garcia, J.V. Predicting alternative splicing. Methods Mol. Biol. 2014, 1126, 411–423. [Google Scholar] [PubMed]
- Yadav, A.R.; Mace, C.R.; Miller, B.L. Examining the interactions of the splicing factor MBNL1 with target RNA sequences via a label-free, multiplex method. Anal. Chem. 2014, 86, 1067–1075. [Google Scholar] [CrossRef] [PubMed]
- Hsu, J.B.; Huang, K.Y.; Weng, T.Y.; Huang, C.H.; Lee, T.Y. Incorporating significant amino acid pairs and protein domains to predict RNA splicing-related proteins with functional roles. J. Comput. Aided Mol. Des. 2014, 28, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Thompson, B.A.; Martins, A.; Spurdle, A.B. A review of mismatch repair gene transcripts: Issues for interpretation of mRNA splicing assays. Clin. Genet. 2015, 87, 100–108. [Google Scholar] [CrossRef]
- Hoffmann, S.; Otto, C.; Doose, G.; Tanzer, A.; Langenberger, D.; Christ, S.; Kunz, M.; Holdt, L.M.; Teupser, D.; Hackermüller, J.; et al. A multi-split mapping algorithm for circular RNA, splicing, trans-splicing and fusion detection. Genome Biol 2014, 15, R34. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.; Lenser, T.; Jahn, N.; Gausmann, U.; Friedel, S.; Szafranski, K.; Huse, K.; Rosenstiel, P.; Hampe, J.; Schuster, S.; et al. TassDB2—A comprehensive database of subtle alternative splicing events. BMC Bioinform. 2010, 11, 216. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, K.; Sloan, S.A.; Bennett, M.L.; Scholze, A.R.; O’Keeffe, S.; Phatnani, H.P.; Guarnieri, P.; Caneda, C.; Ruderisch, N.; et al. An RNA-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex. J. Neurosci. 2014, 34, 11929–11947. [Google Scholar] [CrossRef] [PubMed]
- Hatje, K.; Kollmar, M. Kassiopeia: A database and web application for the analysis of mutually exclusive exomes of eukaryotes. BMC Genomics 2014, 15, 115. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, K.; Lee, J.T.H.; Huang, Z.; Wong, C.-M. Coupling and Coordination in Gene Expression Processes with Pre-mRNA Splicing. Int. J. Mol. Sci. 2015, 16, 5682-5696. https://doi.org/10.3390/ijms16035682
Pan K, Lee JTH, Huang Z, Wong C-M. Coupling and Coordination in Gene Expression Processes with Pre-mRNA Splicing. International Journal of Molecular Sciences. 2015; 16(3):5682-5696. https://doi.org/10.3390/ijms16035682
Chicago/Turabian StylePan, Kewu, Jimmy Tsz Hang Lee, Zhe Huang, and Chi-Ming Wong. 2015. "Coupling and Coordination in Gene Expression Processes with Pre-mRNA Splicing" International Journal of Molecular Sciences 16, no. 3: 5682-5696. https://doi.org/10.3390/ijms16035682
APA StylePan, K., Lee, J. T. H., Huang, Z., & Wong, C. -M. (2015). Coupling and Coordination in Gene Expression Processes with Pre-mRNA Splicing. International Journal of Molecular Sciences, 16(3), 5682-5696. https://doi.org/10.3390/ijms16035682