
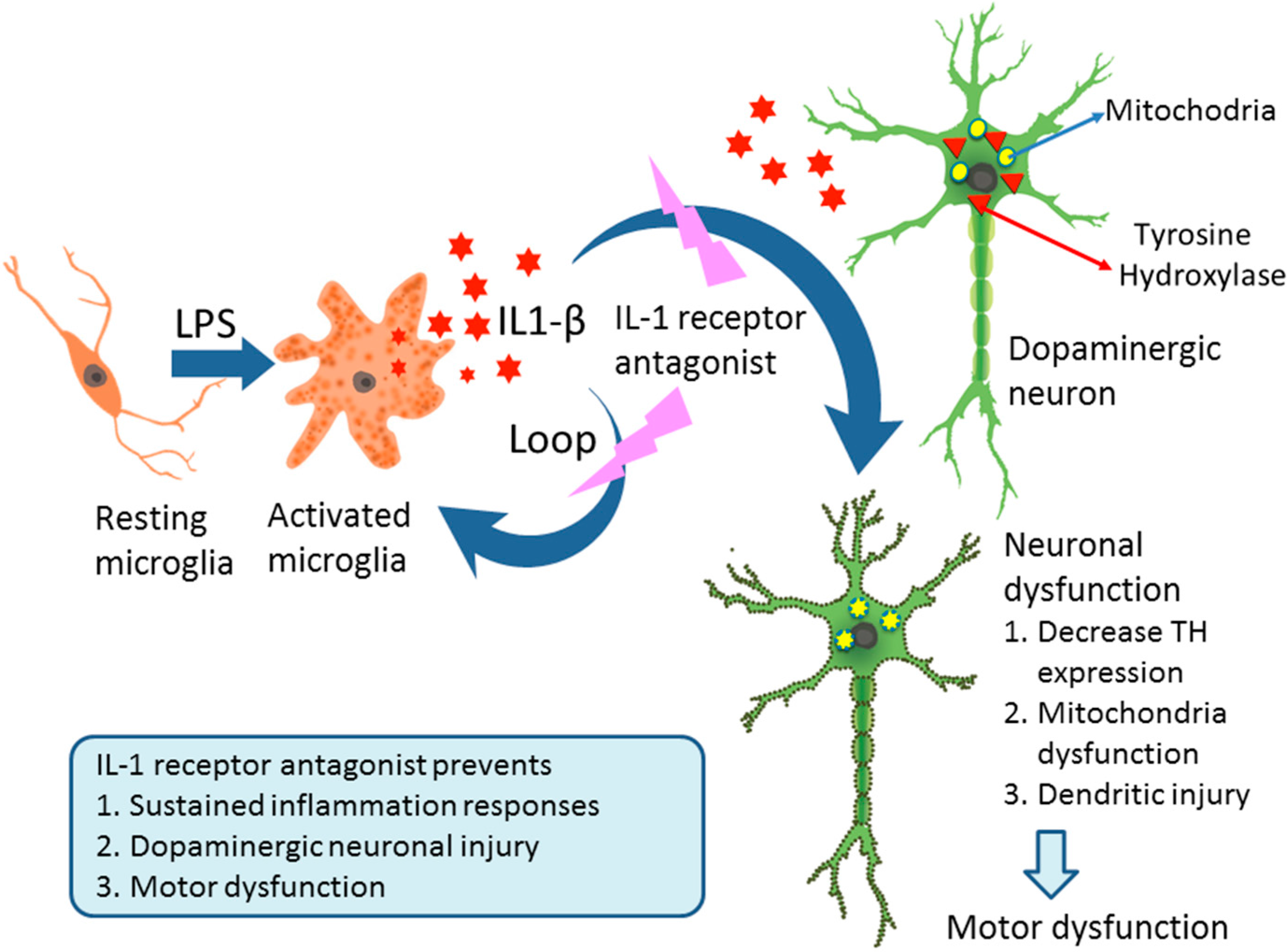
Interleukin-1 Receptor Antagonist Reduces Neonatal Lipopolysaccharide-Induced Long-Lasting Neurobehavioral Deficits and Dopaminergic Neuronal Injury in Adult Rats


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
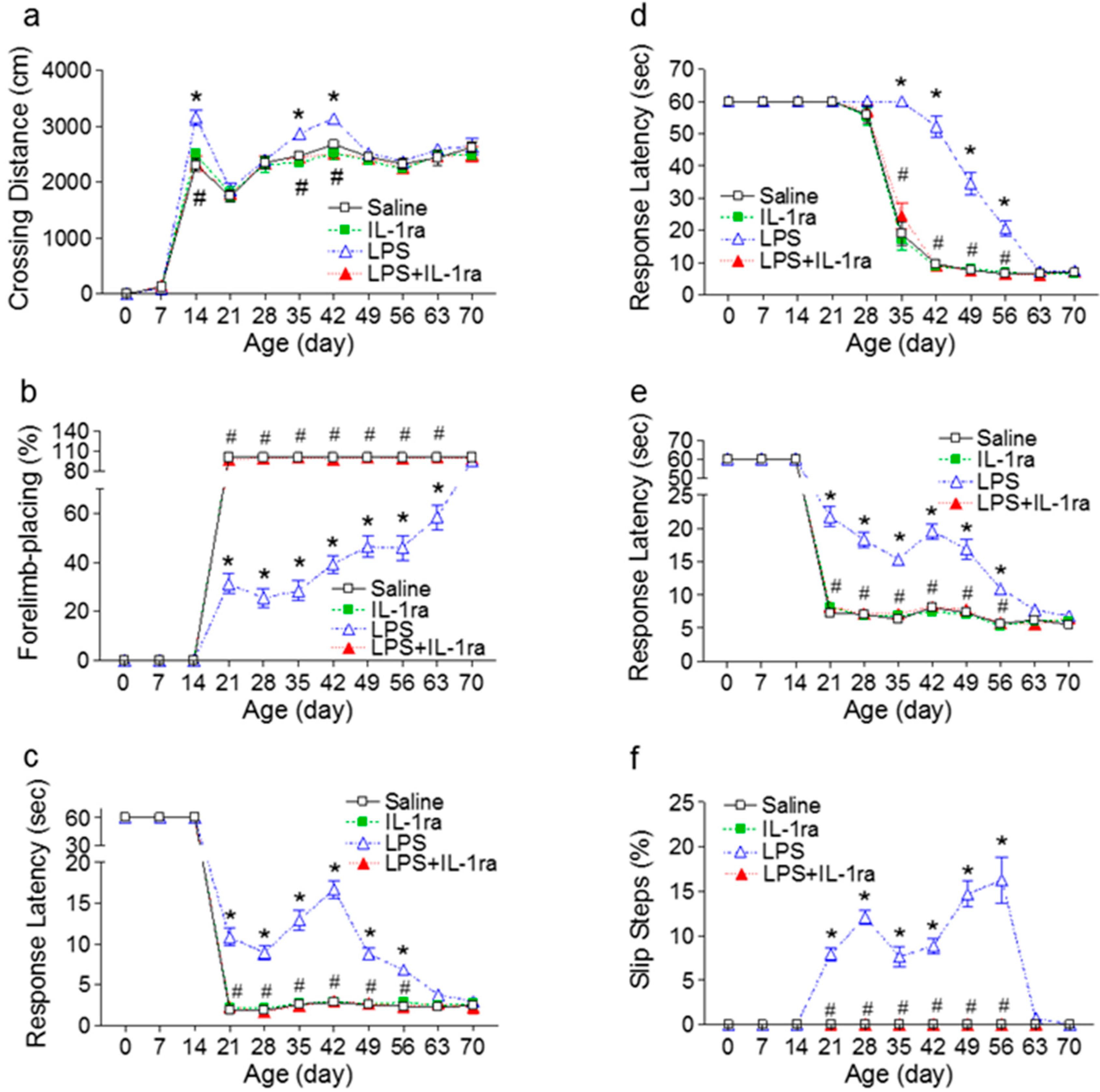
2.1. IL-1ra Attenuated Neurobehavioral Deficits Induced by Lipopolysaccharide (LPS) Exposure

2.1.1. Locomotion
2.1.2. Vibrissa-Elicited Forelimb-Placing Test
2.1.3. Movement Initiation Test
2.1.4. Pole Test
2.1.5. Tapered/Ledged Beam Walking Test
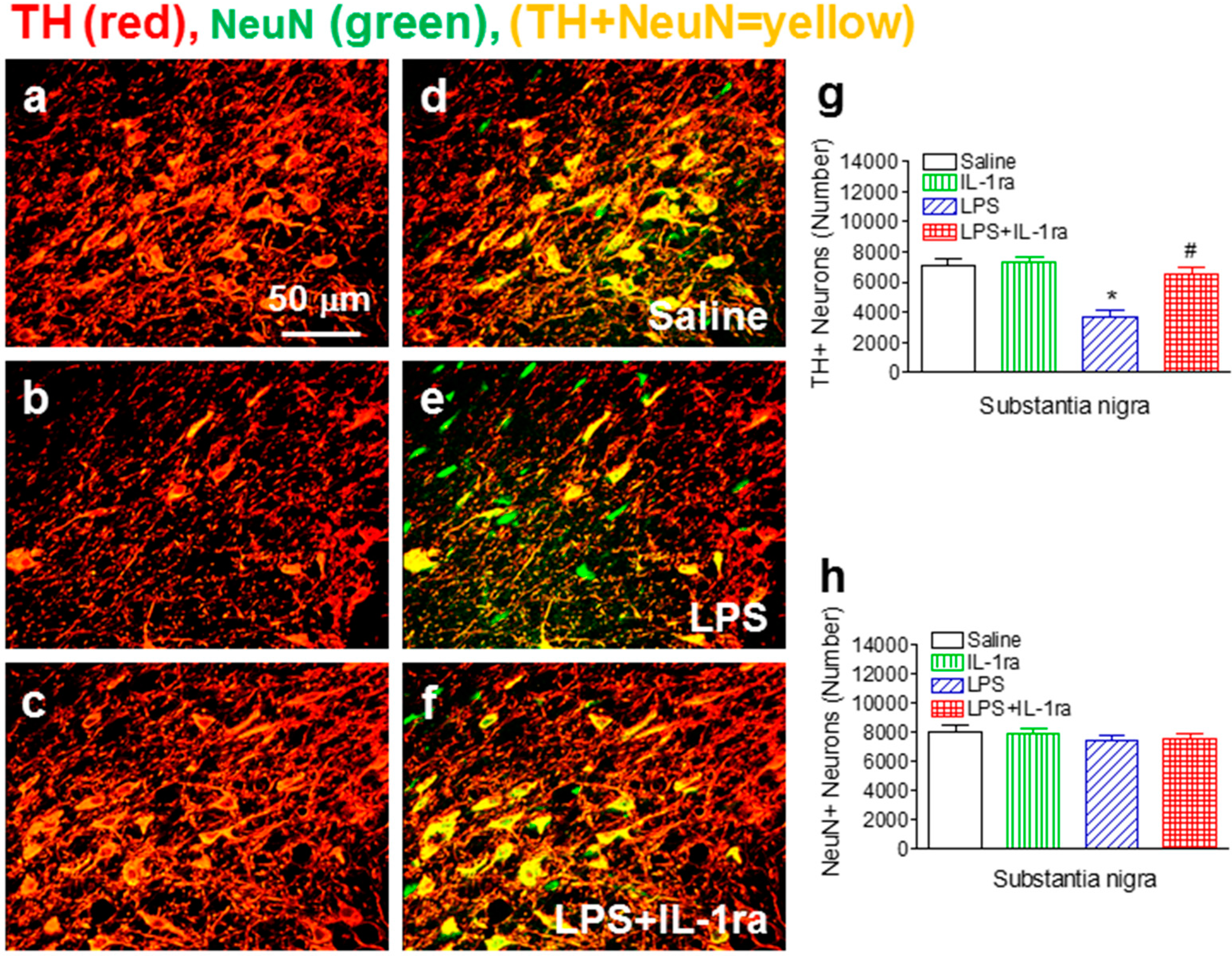
2.2. IL-1ra Attenuated Neonatal LPS Exposure-Induced Losses of TH-Immunoreactive Neurons

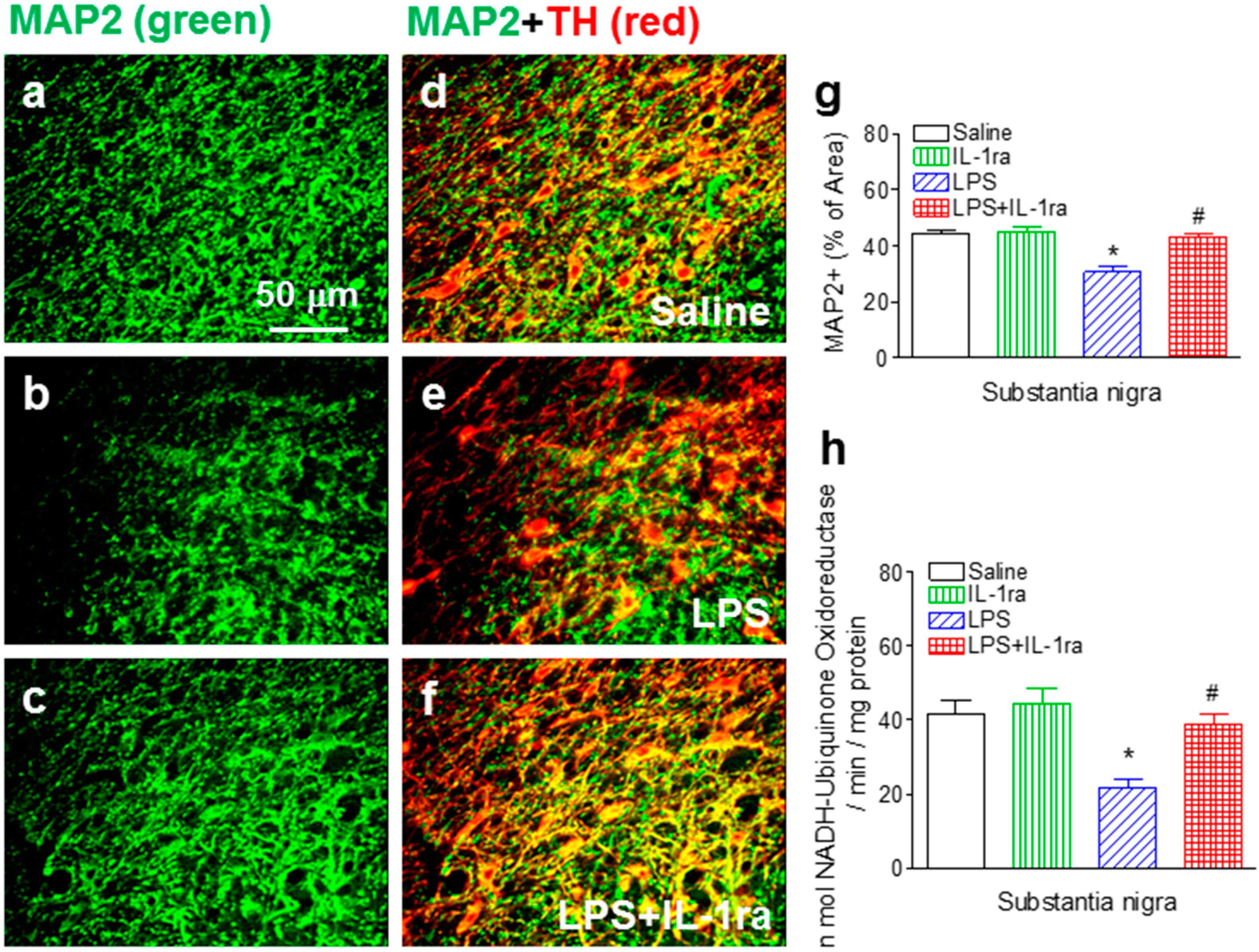
2.3. IL-1ra Attenuated Neonatal LPS Exposure-Induced Loss of Dopaminergic Dendrites and Reduction in Mitochondrial Complex I Activity

2.4. IL-1ra Suppressed Chronic Microglia Activation Following LPS Exposure

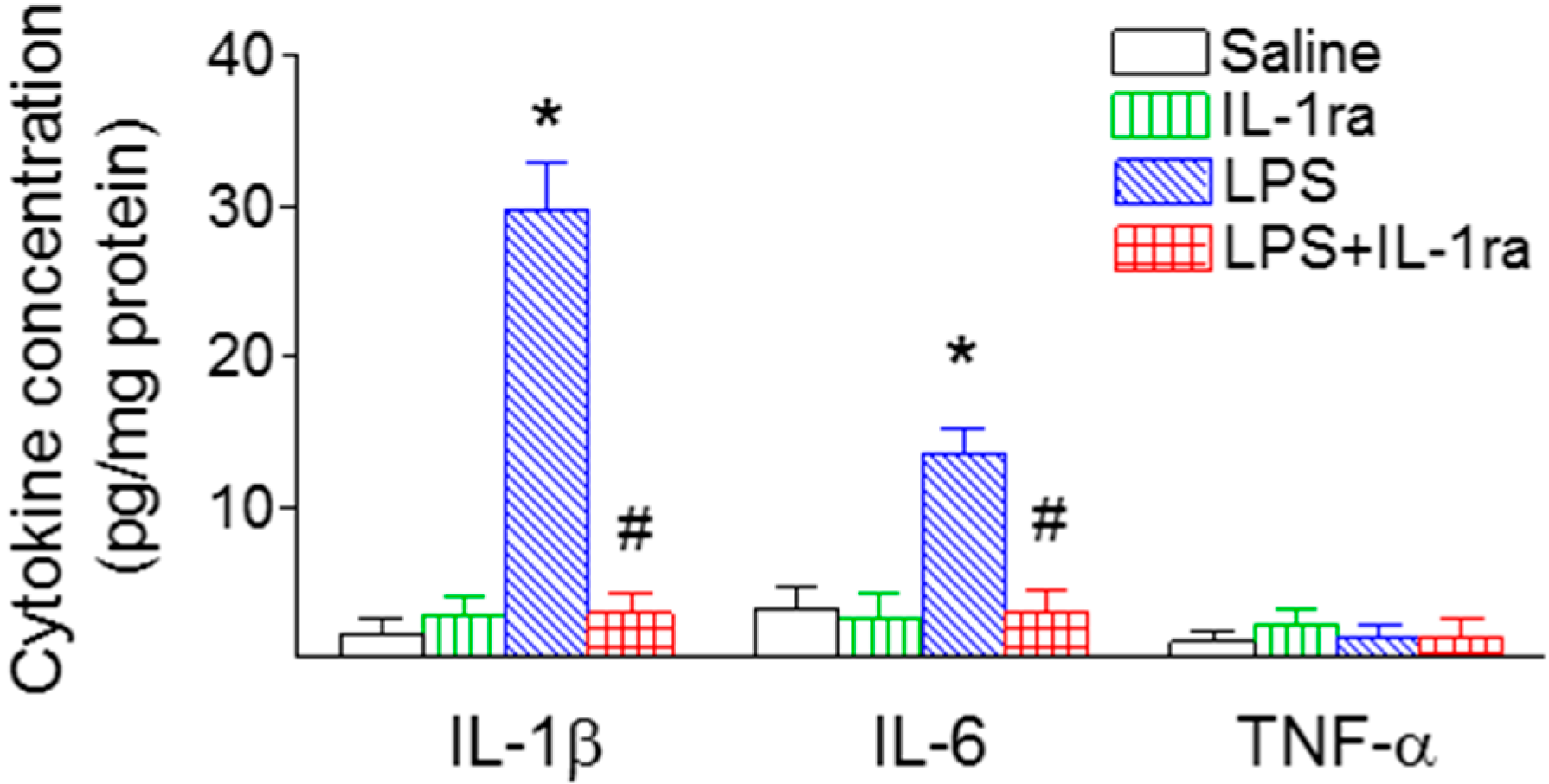
2.5. IL-1ra Significantly Reduced Proinflammatory Cytokine Production Following LPS Exposure

2.6. IL-1ra Attenuated Enhanced Locomotor Activity in LPS-Exposed Rats Following Methamphetamine Stimulation


3. Experimental Section
3.1. Chemicals
3.2. Animals
3.3. Surgical Procedures and Animal Treatment
3.4. Behavioral Testing
3.4.1. Locomotor Activity
3.4.2. Vibrissa-Elicited Forelimb-Placing Test
3.4.3. Movement Initiation Test
3.4.4. Pole Test
3.4.5. Tapered/Ledged Beam Walking Test
3.4.6. Methamphetamine-Induced Locomotion
3.5. Immunohistochemistry Studies
3.6. Stereological Estimates of the Total Number of Neurons in the Substantia Nigra (SN)
3.7. Determination of Mitochondrial Complex I Activity
3.8. Determination of IL-1β, IL6 and TNFα Protein by ELISA
3.9. Quantification of Immunostaining Data
3.10. Statistics
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hagberg, H.; Gressens, P.; Mallard, C. Inflammation during fetal and neonatal life: Implications for neurologic and neuropsychiatric disease in children and adults. Ann. Neurol. 2012, 71, 444–457. [Google Scholar] [CrossRef] [PubMed]
- Volpe, J.J. Brain injury in premature infants: A complex amalgam of destructive and developmental disturbances. Lancet Neurol. 2009, 8, 110–124. [Google Scholar] [CrossRef] [PubMed]
- Kadhim, H.; Tabarki, B.; de Prez, C.; Sebire, G. Cytokine immunoreactivity in cortical and subcortical neurons in periventricular leukomalacia: Are cytokines implicated in neuronal dysfunction in cerebral palsy? Acta Neuropathol. (Berl.) 2003, 105, 209–216. [Google Scholar]
- Yoon, B.H.; Romero, R.; Kim, C.J.; Koo, J.N.; Choe, G.; Syn, H.C.; Chi, J.G. High expression of tumor necrosis factor-α and interleukin-6 in periventricular leukomalacia. Am. J. Obstet. Gynecol. 1997, 177, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Yoon, B.H.; Romero, R.; Yang, S.H.; Jun, J.K.; Kim, I.O.; Choi, J.H.; Syn, H.C. Interleukin-6 concentrations in umbilical cord plasma are elevated in neonates with white matter lesions associated with periventricular leukomalacia. Am. J. Obstet. Gynecol. 1996, 174, 1433–1440. [Google Scholar] [CrossRef] [PubMed]
- Yoon, B.H.; Jun, J.K.; Romero, R.; Park, K.H.; Gomez, R.; Choi, J.H.; Kim, I.O. Amniotic fluid inflammatory cytokines (interleukin-6, interleukin-1β, and tumor necrosis factor-α), neonatal brain white matter lesions, and cerebral palsy. Am. J. Obstet. Gynecol. 1997, 177, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Raetz, C.R.; Whitfield, C. Lipopolysaccharide endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700. [Google Scholar] [CrossRef] [PubMed]
- Boksa, P. Effects of prenatal infection on brain development and behavior: A review of findings from animal models. Brain. Behav. Immun. 2010, 24, 881–897. [Google Scholar] [CrossRef] [PubMed]
- Dutta, G.; Zhang, P.; Liu, B. The lipopolysaccharide Parkinson’s disease animal model: Mechanistic studies and drug discovery. Fundam. Clin. Pharmacol. 2008, 22, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.W.; Tien, L.T.; Lin, R.C.; Simpson, K.L.; Rhodes, P.G.; Cai, Z. Neonatal exposure to lipopolysaccharide enhances vulnerability of nigrostriatal dopaminergic neurons to rotenone neurotoxicity in later life. Neurobiol. Dis. 2011, 44, 304–316. [Google Scholar] [CrossRef] [PubMed]
- Feleder, C.; Tseng, K.Y.; Calhoon, G.G.; O’Donnell, P. Neonatal intrahippocampal immune challenge alters dopamine modulation of prefrontal cortical interneurons in adult rats. Biol. Psychiatry 2010, 67, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Ling, Z.; Zhu, Y.; Tong, C.; Snyder, J.A.; Lipton, J.W.; Carvey, P.M. Progressive dopamine neuron loss following supra-nigral lipopolysaccharide (LPS) infusion into rats exposed to LPS prenatally. Exp. Neurol. 2006, 199, 499–512. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.W.; Pang, Y.; Lin, S.; Tien, L.T.; Ma, T.; Rhodes, P.G.; Cai, Z. Minocycline reduces lipopolysaccharide-induced neurological dysfunction and brain injury in the neonatal rat. J. Neurosci. Res. 2005, 82, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.W.; Tien, L.T.; Mitchell, H.J.; Rhodes, P.G.; Cai, Z. α-Phenyl-n-tert-butyl-nitrone ameliorates hippocampal injury and improves learning and memory in juvenile rats following neonatal exposure to lipopolysaccharide. Eur. J. Neurosci. 2008, 27, 1475–1484. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.W.; Tien, L.T.; Zheng, B.; Pang, Y.; Lin, R.C.; Simpson, K.L.; Ma, T.; Rhodes, P.G.; Cai, Z. Dopaminergic neuronal injury in the adult rat brain following neonatal exposure to lipopolysaccharide and the silent neurotoxicity. Brain Behav. Immun. 2011, 25, 286–297. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, N.J. Functions and mechanisms of interleukin-1 in the brain. Trends Pharmaceut. Sci. 1991, 12, 430–436. [Google Scholar] [CrossRef]
- Godoy, M.C.; Tarelli, R.; Ferrari, C.C.; Sarchi, M.I.; Pitossi, F.J. Central and systemic IL-1 exacerbates neurodegeneration and motor symptoms in a model of Parkinson’s disease. Brain 2008, 131, 1880–1894. [Google Scholar] [CrossRef] [PubMed]
- Koprich, J.B.; Reske-Nielsen, C.; Mithal, P.; Isacson, O. Neuroinflammation mediated by IL-1β increases susceptibility of dopamine neurons to degeneration in an animal model of Parkinson’s disease. J. Neuroinflamm. 2008, 5. [Google Scholar] [CrossRef]
- Rubio-Perez, J.M.; Morillas-Ruiz, J.M. A review: Inflammatory process in Alzheimer’s disease, role of cytokines. Sci. World J. 2012, 2012. [Google Scholar] [CrossRef]
- Cai, Z.; Pang, Y.; Lin, S.; Rhodes, P.G. Differential roles of tumor necrosis factor-α and interleukin-1β in lipopolysaccharide-induced brain injury in the neonatal rat. Brain Res. 2003, 975, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.C.; Wang, S.J.; Fan, L.W.; Cai, Z.; Rhodes, P.G; Tien, L.T. Interleukin-1 receptor antagonist ameliorates neonatal lipopolysaccharide-induced long-lasting hyperalgesia in the adult rats. Toxicology 2011, 279, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.C.; Ulane, C.M.; Burke, R.E. Clinical progression in Parkinson disease and the neurobiology of axons. Ann. Neurol. 2010, 67, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Lesage, S.; Brice, A. Parkinson’s disease: From monogenic forms to genetic susceptibility factors. Hum. Mol. Genet. 2009, 18, R48–R59. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.; Cai, Z.; Rhodes, P.G. Disturbance of oligodendrocyte development, hypomyelination and white matter injury in the neonatal rat brain after intracerebral injection of lipopolysaccharide. Brain Res. Dev. Brain Res. 2003, 140, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Kreutzberg, G.W. Microglia: A sensor for pathological events in the CNS. Trends. Neurosci. 1996, 19, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Karila, L.; Weinstein, A.; Aubin, H.J.; Benyamina, A.; Reynaud, M.; Batki, S.L. Pharmacological approaches to methamphetamine dependence: A focused review. Br. J. Clin. Pharmacol. 2010, 69, 578–592. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.G.; Aschner, M.; Vitalone, A.; Syversen, T.; Soldin, O.P. Developmental neuropathology of environmental agents. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 87–110. [Google Scholar] [CrossRef] [PubMed]
- Fortier, M.E.; Joober, R.; Luheshi, G.N.; Boksa, P. Maternal exposure to bacterial endotoxin during pregnancy enhances amphetamine-induced locomotion and startle responses in adult rat offspring. J. Psychiatr. Res. 2004, 38, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Ba, A.; Seri, B.V. Psychomotor functions in developing rats: Ontogenetic approach to structure-function relationships. Neurosci. Biobehav. Rev. 1995, 19, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.W.; Mirchell, H.J.; Rhodes, P.G.; Cai, Z. α-Phenyl-n-tert-butyl-nitrone attenuates lipopolysaccharide-induced neuronal injury in the neonatal rat brain. Neuroscience 2008, 151, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.W.; Chen, R.F.; Mitchell, H.J.; Lin, R.C.S.; Simpson, K.L.; Rhodes, P.G.; Cai, Z. α-Phenyl-n-tert-butyl-nitrone attenuates lipopolysaccharide-induced brain injury and improves neurological reflexes and early sensorimotor behavioral performance in juvenile rats. J. Neurosci. Res. 2008, 86, 3536–3547. [Google Scholar] [CrossRef] [PubMed]
- Lehnardt, S.; Lachance, C.; Patrizi, S.; Lefebvre, S.; Follett, P.; Jensen, F.E.; Rosenberg, P.A.; Volpe, J.J.; Vartanian, T. The Toll-like receptor TLP4 is necessary for lipopolysaccharide-induced oligodendrocyte injury in the CNS. J. Neurosci. 2002, 22, 2478–2486. [Google Scholar] [PubMed]
- Pont-Lezica, L.; Bechade, C.; Belarif-Cantaut, Y.; Pascual, O.; Bessis, A. Physiological roles of microglia during development. J. Neurochem. 2011, 119, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Harry, G.J.; Kraft, A.D. Microglia in the developing brain: A potential target with lifetime effects. Neurotoxicology 2012, 33, 191–206. [Google Scholar] [CrossRef] [PubMed]
- Glass, C.K.; Saijo, K.; Winner, B.; Marchetto, M.C.; Gage, F.H. Mechanisms underlying inflammation in neurodegeneration. Cell 2010, 140, 918–934. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Flood, P.M.; Hong, J.S. Neuroinflammation is a key player in Parkinson’s disease and a prime target for therapy. J. Neural. Transm. 2010, 117, 971–979. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.W.; Mirchell, H.J.; Tien, L.T.; Zheng, B.; Pang, Y.; Rhodes, P.G.; Cai, Z. α-Phenyl-n-tert-butyl-nitrone reduces lipopolysaccharide-induced white matter injury in the neonatal rat brain. Dev. Neurol. 2008, 68, 365–378. [Google Scholar]
- Holmin, S.; Mathiesen, T. Intracerebral administration of interleukin-1β and induction of inflammation, apoptosis, and vasogenic edema. J. Neurosurg. 2000, 92, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Altman, J.; Sudarshan, K. Postnatal development of locomotion in the laboratory rat. Anim. Behav. 1975, 23, 896–920. [Google Scholar] [CrossRef] [PubMed]
- Altman, J.; Sudarshan, K.; Das, G.D.; McCormick, N.; Barnes, D. The influence of nutrition on neural and behavioral development: III. Development of some motor, particularly locomotor patterns during infancy. Dev. Psychobiol. 1971, 4, 97–114. [Google Scholar] [CrossRef] [PubMed]
- Hermans, R.H.; Hunter, D.E.; McGivern, R.F.; Cain, C.D.; Longo, L.D. Behavioral sequelae in young rats of acute intermittent antenatal hypoxia. Neurotoxicol. Teratol. 1992, 14, 119–129. [Google Scholar] [CrossRef] [PubMed]
- De Ryck, M.; van Reempts, J.; Duytschaever, H.; van Deuren, B.; Clincke, G. Neocortical localization of tactile/proprioceptive limb placing reactions in the rat. Brain Res. 1992, 573, 44–60. [Google Scholar] [CrossRef] [PubMed]
- Schallert, T.; Woodlee, M.T. Motor Systems: Orienting and Placing. In The Behaviour of the Laboratory Rat: A Handbook with Tests; Whishaw, I.Q., Kolb, B., Eds.; Oxford University Press: New York, NY, USA, 2005; pp. 129–140. [Google Scholar]
- Woodlee, M.T.; Asseo-garcia, A.M.; Zhao, X.; Liu, S.J.; Jones, T.A.; Schallert, T. Testing forelimb placing “across the midline” reveals distinct, lesion-dependent patterns of recovery in rats. Exp. Neurol. 2005, 191, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Fleming, S.M.; Zhu, C.; Fernagut, P.O.; Mehta, A.; Dicarlo, C.D.; Seaman, R.L.; Xhesselet, M.F. Behavioral and immunohistochemical effects of chronic intravenous and subcutaneous infusions of varying doses of rotenone. Exp. Neurol. 2004, 187, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Tillerson, J.L.; Cohen, A.D.; Caudle, W.M.; Zigmond, M.J.; Schallert, T.; Miller, G.W. Forced nonuse in unilateral parkinsonian rats exacerbates injury. J. Neurosci. 2002, 22, 6790–6799. [Google Scholar] [PubMed]
- Zhao, C.S.; Puurunen, K.; Schallert, T.; Sivenius, J.; Jolkkonen, J. Effect of cholinergic medication, before and after focal photothrombotic ischemic cortical injury, on histological and functional outcome in aged and young adult rats. Behav. Brain Res. 2005, 156, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Lokkegaard, A.; Nyengaard, J.R.; West, M.J. Stereological estimates of number and length of capillaries in subdivisions of the human hippocampal region. Hippocampus 2001, 11, 726–740. [Google Scholar] [CrossRef] [PubMed]
- Gundersen, H.J.G.; Jensen, E.B. The efficiency of systematic sampling in stereology and its prediction. J. Microsc. 1987, 147, 229–263. [Google Scholar] [CrossRef] [PubMed]
- Pakkenberg, B.; Gundersen, H.J. Total number of neurons and glial cells in human brain nuclei estimated by the disector and the fractionator. J. Microsc. 1988, 150, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Champy, P.; Höglinger, G.U.; Féger, J.; Gleye, C.; Hocquemiller, R.; Laurens, A.; Guérineau, V.; Laprévote, O.; Medja, F.; Lombès, A.; et al. Annonacin, a lipophilic inhibitor of mitochondrial complex I, induces nigral and striatal neurodegeneration in rats: Possible relevance for atypical parkinsonism in Guadeloupe. J. Neurochem. 2004, 88, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Hoglinger, G.U.; Lannuzel, A.; Khondiker, M.E.; Michel, P.P.; Duyckaerts, C.; Feger, J.; Champy, P.; Prigent, A.; Medja, F.; Lombes, A.; et al. The mitochondrial complex I inhibitor rotenone triggers a cerebral tauopathy. J. Neurochem. 2005, 95, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.R.; Chen, C.L.; Zhang, L.; Green-Church, K.B.; Zweier, J.L. Superoxide generation from mitochondrial NADH dehydrogenase induces self-inactivation with specific protein radical formation. J. Biol. Chem. 2005, 280, 37339–37348. [Google Scholar] [CrossRef] [PubMed]
- Hammond, J.B.; Kruger, N.J. The Bradford method for protein quantitation. Methods Mol. Biol. 1988, 3, 25–32. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pang, Y.; Tien, L.-T.; Zhu, H.; Shen, J.; Wright, C.F.; Jones, T.K.; Mamoon, S.A.; Bhatt, A.J.; Cai, Z.; Fan, L.-W. Interleukin-1 Receptor Antagonist Reduces Neonatal Lipopolysaccharide-Induced Long-Lasting Neurobehavioral Deficits and Dopaminergic Neuronal Injury in Adult Rats. Int. J. Mol. Sci. 2015, 16, 8635-8654. https://doi.org/10.3390/ijms16048635
Pang Y, Tien L-T, Zhu H, Shen J, Wright CF, Jones TK, Mamoon SA, Bhatt AJ, Cai Z, Fan L-W. Interleukin-1 Receptor Antagonist Reduces Neonatal Lipopolysaccharide-Induced Long-Lasting Neurobehavioral Deficits and Dopaminergic Neuronal Injury in Adult Rats. International Journal of Molecular Sciences. 2015; 16(4):8635-8654. https://doi.org/10.3390/ijms16048635
Chicago/Turabian StylePang, Yi, Lu-Tai Tien, Hobart Zhu, Juying Shen, Camilla F. Wright, Tembra K. Jones, Samir A. Mamoon, Abhay J. Bhatt, Zhengwei Cai, and Lir-Wan Fan. 2015. "Interleukin-1 Receptor Antagonist Reduces Neonatal Lipopolysaccharide-Induced Long-Lasting Neurobehavioral Deficits and Dopaminergic Neuronal Injury in Adult Rats" International Journal of Molecular Sciences 16, no. 4: 8635-8654. https://doi.org/10.3390/ijms16048635
APA StylePang, Y., Tien, L. -T., Zhu, H., Shen, J., Wright, C. F., Jones, T. K., Mamoon, S. A., Bhatt, A. J., Cai, Z., & Fan, L. -W. (2015). Interleukin-1 Receptor Antagonist Reduces Neonatal Lipopolysaccharide-Induced Long-Lasting Neurobehavioral Deficits and Dopaminergic Neuronal Injury in Adult Rats. International Journal of Molecular Sciences, 16(4), 8635-8654. https://doi.org/10.3390/ijms16048635