
Plasmids from Food Lactic Acid Bacteria: Diversity, Similarity, and New Developments

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Diversity and Similarity of Plasmids from LAB
2.1. Plasmids of Genus Lactobacillus
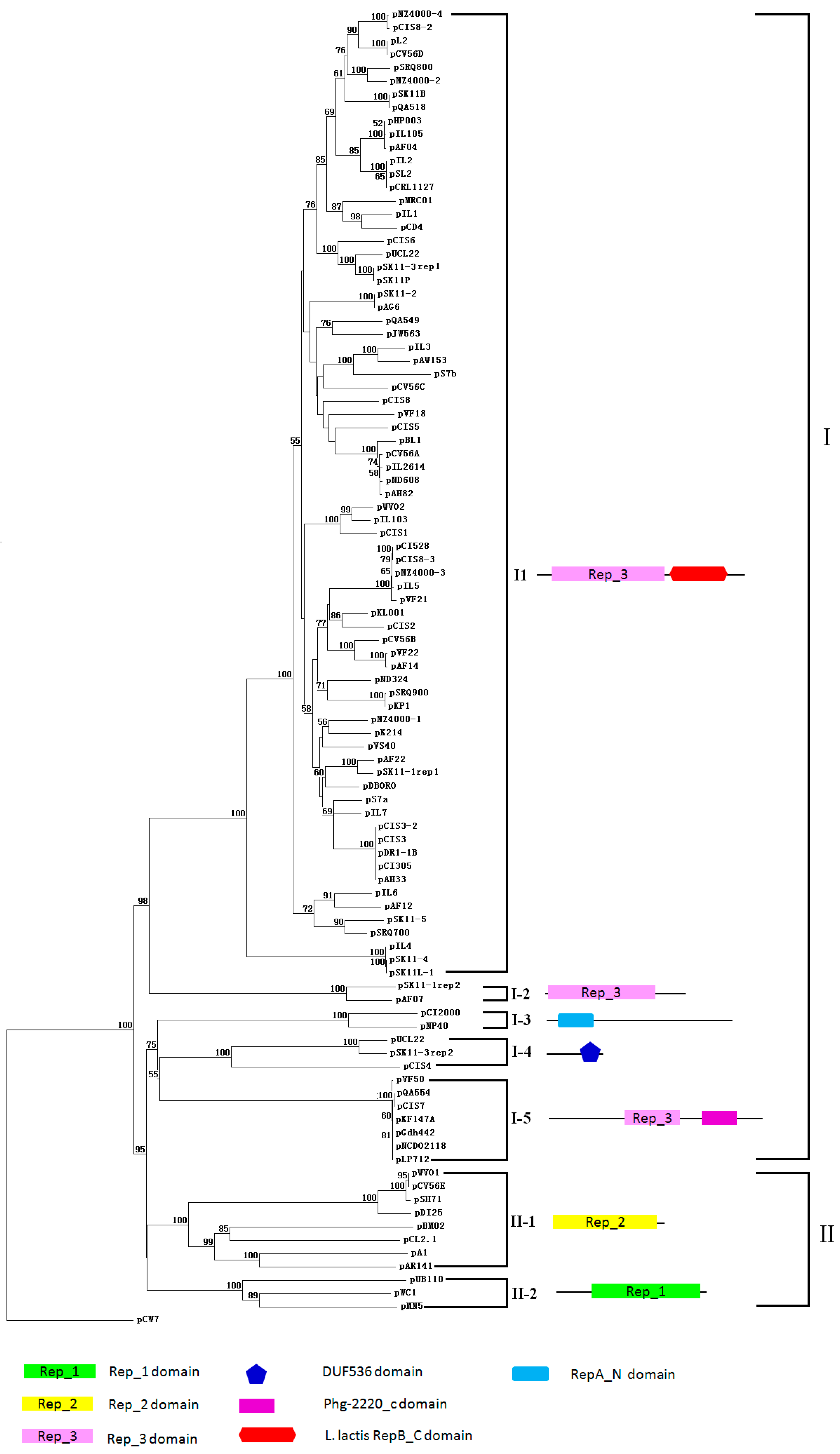
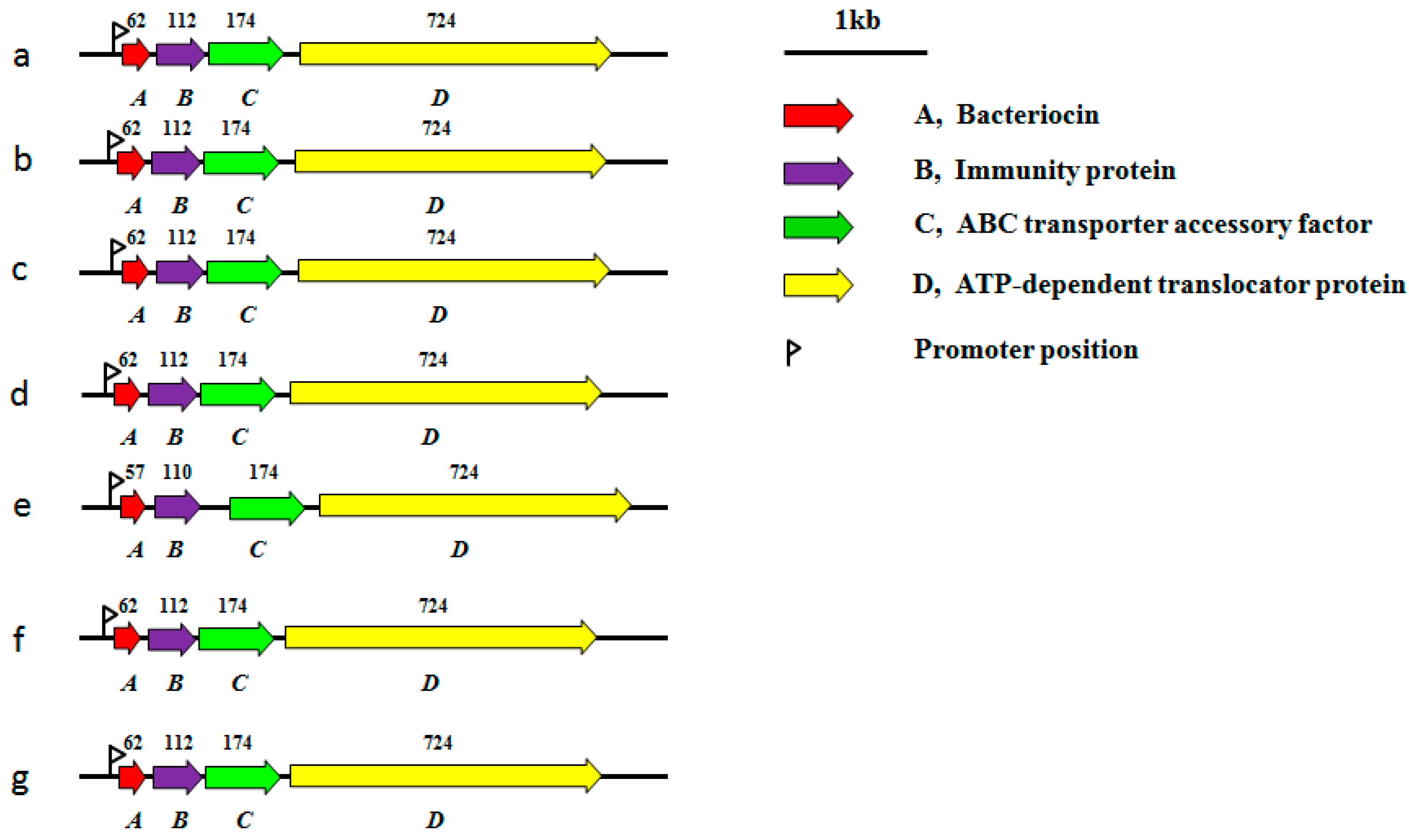
2.2. Plasmids of Genus Lactococcus

2.3. Plasmids of Genus Pediococcus

2.4. Plasmids of Genus Enterococcus
2.5. Plasmids of Streptococcus thermophilus
2.6. Plasmids of Genus Bifidobacterium
2.7. Plasmids of Genus Oenococcus
3. Plasmid Replication Mechanisms
3.1. Rolling-Circle Replicating Plasmids
3.2. Theta-Type-Replicating Plasmids
4. Mobility of LAB Plasmids
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sun, Z.H.; Yu, J.; Dan, T.; Zhang, W.Y.; Zhang, H.P. Phylogenesis and evolution of lactic acid Bacteria. In Lactic Acid Bacteria-Fundamentals and Practice, 1st ed.; Zhang, H.P., Cai, Y.M., Eds.; Springer Publishing Inc.: New York, NY, USA, 2014; pp. 1–101. [Google Scholar]
- Liu, W.J.; Pang, H.L.; Zhang, H.P.; Cai, Y.M. Biodiversity of lactic acid bacteria. In Lactic Acid Bacteria-Fundamentals and Practice, 1st ed.; Zhang, H.P., Cai, Y.M., Eds.; Springer Publishing Inc.: New York, NY, USA, 2014; pp. 103–203. [Google Scholar]
- Wegrzyn, G.; Wegrzyn, A. Stress responses and replication of plasmids in bacterial cells. Microb. Cell. Fact. 2002, 1, 2. [Google Scholar] [CrossRef] [PubMed]
- Gasson, M.J. In vivo genetic systems in lactic acid bacteria. FEMS Microbiol. Rev. 1990, 7, 43–60. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.Y.; Zhang, H.P. Genomics of Lactic Acid Bacteria. In Lactic Acid Bacteria-Fundamentals and Practice, 1st ed.; Zhang, H.P., Cai, Y.M., Eds.; Springer Publishing Inc.: New York, NY, USA, 2014; pp. 235–238. [Google Scholar]
- Wang, T.T.; Lee, B.H. Plasmids in Lactobacillus. Crit. Rev. Biotechnol. 1997, 17, 227–272. [Google Scholar] [CrossRef] [PubMed]
- Shareck, J.; Choi, Y.; Lee, B.; Miguez, C.B. Cloning vectors based on cryptic plasmids isolated from lactic acid bacteria: Their characteristics and potential applications in biotechnology. Crit. Rev. Biotechnol. 2004, 24, 155–208. [Google Scholar] [CrossRef] [PubMed]
- Schroeter, J.; Klaenhammer, T. Genomics of lactic acid bacteria. FEMS Microbiol. Lett. 2009, 292, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Mills, S.; McAuliffe, O.E.; Coffey, A.; Fitzgerald, G.F.; Ross, R.P. Plasmids of lactococci-genetic accessories or genetic necessities? FEMS Microbiol. Rev. 2006, 30, 243–273. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, S.; Stockdale, S.; Bottacini, F.; Mahony, J.; van Sinderen, D. The Lactococcus lactis plasmidome: Much learnt, yet still lots to discover. FEMS Microbiol. Rev. 2014, 38, 1066–1088. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Du, X.H.; Wang, W.H.; Zhang, J.C.; Liu, W.J.; Sun, Z.H.; Sun, T.S.; Zhang, H.P. Phenotypic and genotypic characteristics of lactic acid bacteria isolated from sour congee in Inner Mongolia of China. J. Gen. Appl. Microbiol. 2011, 57, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Galloway-Peña, J.R.; Sillanpaa, J.; Roh, J.H.; Nallapareddy, S.R.; Chowdhury, S.; Bourgogne, A.; Choudhury, T.; Muzny, D.M.; Buhay, C.J.; et al. Complete genome sequence of Enterococcus faecium strain TX16 and comparative genomic analysis of Enterococcus faecium genomes. BMC Microbiol. 2012, 12, 135. [Google Scholar] [CrossRef] [PubMed]
- Dellaglio, F.; Felis, G.E. Taxonomy oflactobacilli and bifidobacteria. In Probiotics and Prebiotics: Scientific Aspects, 1st ed.; Tannock, G.W., Ed.; Caister Academic Press: Norfolk, UK, 2005; pp. 25–49. [Google Scholar]
- Jiménez, E.; Martín, R.; Maldonado, A.; Martín, V.; de Segura, A.G.; Fernández, L.; Rodríguez, J.M. Complete genome sequence of Lactobacillus salivarius CECT 5713, a probiotic strain isolated from human milk and infant feces. J. Bacteriol. 2010, 192, 5266–5267. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Zhang, L.; Li, J.C.; Chen, T.; Chen, W.; Wang, G.K.; Yin, J.H. Characterization of pLP18, a novel cryptic plasmid of Lactobacillus plantarum PC518 isolated from Chinese pickle. Plasmid 2011, 65, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Siezen, R.J.; Johan, E.T.; van Hylckama Vlieg, J.E.T. Genomic diversity andversatility of Lactobacillus plantarum, a natural metabolic engineer. Microb. Cell. Fact. 2011, 10, S3. [Google Scholar] [CrossRef] [PubMed]
- Guidone, A.; Zotta, T.; Ross, R.P.; Stanton, C.; Rea, M.C.; Parente, E.; Ricciardi, A. Functional properties of Lactobacillus plantarum strains: A multivariate screening study. LWT—Food Sci. Technol. 2014, 56, 69–76. [Google Scholar] [CrossRef]
- Bove, P.; Gallone, A.; Russo, P.; Capozzi, V.; Albenzio, M.; Spano, G.; Fiocco, D. Probiotic features of Lactobacillus plantarum mutant strains. Appl. Microbiol. Biotechnol. 2012, 96, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Fiocco, D.; Capozzi, V.; Gallone, A.; Hols, P.; Guzzo, J.; Weidmann, S.; Rieu, A.; Msadek, T.; Spano, G. Characterization of the CtsR stress response regulon in Lactobacillus plantarum. J. Bacteriol. 2010, 196, 896–900. [Google Scholar] [CrossRef] [PubMed]
- Crowley, S.; Bottacini, F.; Mahony, J.; van Sinderen, D. Complete genome sequence of Lactobacillus plantarum strain 16, a broad-spectrum antifungal-producing lactic acid bacterium. Genome Announc. 2013, 1, e00533-13. [Google Scholar] [CrossRef] [PubMed]
- Ahn, C.; Collins-Thompson, D.; Duncan, C.; Stiles, M.E. Mobilization and location of the genetic determinant of chloramphenicol resistance from Lactobacillus plantarum caTC2R. Plasmid 1992, 27, 169–176. [Google Scholar] [CrossRef]
- Danielsen, M. Characterization of the tetracycline resistance plasmid pMD5057 from Lactobacillus plantarum 5057 reveals a composite structure. Plasmid 2002, 48, 98–103. [Google Scholar] [CrossRef]
- Huys, G.; D’Haene, K.; Swings, J. Genetic basis of tetracycline and minocycline resistance in potentially probiotic Lactobacillus plantarum strain CCUG 43738. Antimicrob. Agents Chemother. 2006, 50, 1550–1551. [Google Scholar] [CrossRef] [PubMed]
- Feld, L.; Bielak, E.; Hammer, K.; Wilcks, A. Characterization of a small erythromycin resistance plasmid pLFE1 from the food-isolate Lactobacillus plantarum M345. Plasmid 2009, 61, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Ai, L.Z.; Zhou, F.F.; Ren, J.; Sun, K.J.; Zhang, H.; Chen, W.; Guo, B.H. Complete nucleotide sequence of plasmid pST-III from Lactobacillus plantarum ST-III. Plasmid 2012, 67, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, T.; Doi, K.; Nishiyama, K.; Ohmomo, S.; Ogata, S. Characterization of a phage resistance plasmid, pLKS, of silage-making Lactobacillus plantarum NGRI0101. Biosci. Biotechnol. Biochem. 2000, 64, 751–756. [Google Scholar] [CrossRef] [PubMed]
- Van Reenen, C.A.; Chikindas, M.L.; van Zyl, W.H.; Dicks, L.M. Characterization and heterologous expression of a class IIa bacteriocin, plantaricin 423 from Lactobacillus plantarum 423, in Saccharomyces cerevisiae. Int. J. Food Microbiol. 2003, 81, 29–40. [Google Scholar] [CrossRef]
- Van Kranenburg, R.; Golic, N.; Bongers, R.; Leer, R.J.; de Vos, W.M.; Siezen, R.J.; Kleerebezem, M. Functional analysis of three plasmids from Lactobacillus plantarum. Appl. Environ. Microbiol. 2005, 71, 1223–1230. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Hao, Y.; Xie, Y.; Yin, S.; Zhai, Z.Y.; Han, B.Z. Characterization of a rolling-circle replication plasmid pXY3 from Lactobacillus plantarum XY3. Plasmid 2010, 64, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Cho, G.S.; Huch, M.; Mathara, J.M.; van Belkum, M.J.; Franz, C.M. Characterization of pMRI 5.2, a rolling-circle-type plasmid from Lactobacillus plantarum BFE 5092 which harbours two different replication initiation genes. Plasmid 2013, 69, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Hao, Y.L.; Zhai, Z.Y.; Li, R.Y.; Huang, Y.; Tian, H.T.; Luo, Y.B. Characterization of a cryptic plasmid pM4 from Lactobacillus plantarum M4. FEMS Microbiol. Lett. 2008, 285, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Xi, X.D.; Fan, J.; Hou, Y.; Gu, J.H.; Shen, W.J.; Li, Z.K.; Cui, Z.L. Characterization of three cryptic plasmids from Lactobacillus plantarum G63 that was isolated from Chinese pickle. Plasmid 2013, 70, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Jalilsood, T.; Baradaran, A.; Ling, F.H.; Mustafa, S.; Yusof, K.; Rahim, R.A. Characterization of pR18, a novel rolling-circle replication plasmid from Lactobacillus plantarum. Plasmid 2014, 73, 1–9. [Google Scholar]
- Flynn, S.; van Sinderen, D.; Thornton, G.M.; Holo, H.; Nes, I.F.; Collins, J.K. Characterization of the genetic locus responsible for the productionof ABP-118, a novel bacteriocin produced by the probiotic bacterium Lactobacillus salivarius subsp. salivarius UCC118. Microbiology 2002, 148, 973–984. [Google Scholar]
- Claesson, M.J.; Li, Y.; Leahy, S.; Canchaya, C.; van Pijkeren, J.P.; Cerdeño-Tárraga, A.M.; Parkhill, J.; Flynn, S.; O’Sullivan, G.C.; Collins, J.K.; et al. Multireplicon genome architecture of Lactobacillus salivarius. Proc. Natl. Acad. Sci. USA 2006, 103, 6718–6723. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Canchaya, C.; Fang, F.; Raftis, E.; Ryan, K.A.; van Pijkeren, J.-P.; van Sinderen, D.; O’Toole, P.W. Distribution of megaplasmids in Lactobacillus salivarius and other Lactobacilli. J. Bacteriol. 2007, 189, 6128–6139. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, J.; Ahmed, Z.; Bai, X.; Wang, J. Complete genome sequence of Lactobacillus kefiranofaciens ZW3. J. Bacteriol. 2011, 193, 4280–4281. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.; Flynn, S.; Li, Y.; Claesson, M.J.; van Pijkeren, J.P.; Collins, J.K.; van Sinderen, D.; O’Toole, P.W. Characterization of endogenous plasmids from Lactobacillus salivarius UCC118. Appl. Environ. Microbiol. 2008, 74, 3216–3228. [Google Scholar] [CrossRef] [PubMed]
- Sørvig, E.; Grönqvist, S.; Naterstad, K.; Mathiesen, G.; Eijsink, V.G.; Axelsson, L. Construction of vectors for inducible gene expressionin Lactobacillus sakei and L. plantarum. FEMS Microbiol. Lett. 2003, 229, 119–126. [Google Scholar] [CrossRef]
- Sørvig, E.; Mathiesen, G.; Naterstad, K.; Eijsink, V.G.; Axelsson, L. High-level, inducible gene expression in Lactobacillus sakei and Lactobacillus plantarum using versatile expression vectors. Microbiology 2005, 151, 2439–2449. [Google Scholar] [CrossRef] [PubMed]
- Halbmayr, E.; Nathiesen, G.; Nguyen, T.H.; Maischberger, T.; Peterbauer, C.K.; Eijsink, V.G.H.; Haltrich, D. High-level expression of recombinant β-galactosidases in Lactobacillus plantarum and Lactobacillus sakei using a sakacin P-based expression system. J. Agric. Food Chem. 2008, 56, 4710–4719. [Google Scholar] [CrossRef] [PubMed]
- Böhmer, N.; Lutz-Wahl, S.; Fischer, L. Recombinant production of hyperthermostable CelB from Pyrococcus furiosus in Lactobacillus sp. Appl. Microbiol. Biotechnol. 2012, 96, 903–912. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.; Mathiesen, G.; Fredriksen, L.; Kittl, R.; Nguyen, T.H.; Eijsink, V.G.H.; Haltrich, D.; Peterbauer, C.K. A food-grade system for inducible gene expression in Lactobacillus plantarum using an alanine racemase-encoding selection marker. J. Agric. Food Chem. 2011, 59, 5617–5624. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Zhai, Z.Y.; Wang, G.H.; An, H.R.; Luo, Y.B.; Hao, Y.L. A novel vector for lactic acid bacteria that uses a bile salt hydrolase gene as a potential food-grade selection marker. J. Biotechnol. 2011, 152, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Böhmer, N.; König, S.; Fischer, L. A novel manganese starvation-inducible expression system for Lactobacillus plantarum. FEMS Microbiol. Lett. 2013, 342, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Wada, T.; Noda, M.; Kashiwabara, F.; Jeon, H.J.; Shirakawa, A.; Yabu, H.; Matoba, Y.; Kumagai, T.; Sugiyama, M. Characterization of four plasmids harboured in a Lactobacillus brevis strain encoding a novel bacteriocin, brevicin 925A, and construction of a shuttle vector for lactic acid bacteria and Escherichia coli. Microbiology 2009, 155, 1726–1737. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.J.; Chang, H.C. Analysis of pYC2, a crypticplasmid in Lactobacillus sakei BM5 isolated from kimchi. Biotechnol. Lett. 2009, 31, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.J.; Chang, H.C. Construction and evaluation of shuttle vector, pGYC4α, based on pYC2 from Lactobacillus sakei. Biotechnol. Lett. 2011, 33, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Panya, M.; Lulitanond, V.; Tangphatsornruang, S.; Namwat, W.; Wannasutta, R.; Suebwongsa, N.; Mayo, B. Sequencing and analysis of three plasmids from Lactobacillus casei TISTR1341 and development of plasmid-derived Escherichia coli–L. casei shuttle vectors. Appl. Microbiol. Biotechnol. 2012, 93, 261–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Lin, J.; Ma, C.; Zhao, S.; She, Q.; Liang, Y. Characterization of pMC11, a plasmid with dual origins of replication isolated from Lactobacillus casei MCJ and construction of shuttle vectors with each replicon. Appl. Microbiol. Biotechnol. 2014, 98, 5977–5989. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D.; Fitzgerald, G.F.; McAuliffe, O. From field to fermentation: The origins of Lactococcus lactis and its domestication to the dairy environment. Food Microbiol. 2015, 47, 45–61. [Google Scholar] [CrossRef] [PubMed]
- Kelly, W.J.; Ward, L.J.; Leahy, S.C. Chromosomal diversity in Lactococcus lactis and the origin of dairy starter cultures. Genome Biol. Evol. 2010, 2, 729–744. [Google Scholar] [CrossRef] [PubMed]
- Van Hylckama Vlieg, J.E.; Rademaker, J.L.; Bachmann, H.; Molenaar, D.; Kelly, W.J.; Siezen, R.J. Natural diversity and adaptive responses of Lactococcus lactis. Curr. Opin. Biotechnol. 2006, 17, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Sanders, J.W.; Venema, G.; Kok, J. Environmental stress responses in Lactococcus lactis. FEMS Microbiol. Rev. 1999, 23, 483–501. [Google Scholar] [CrossRef]
- Ainsworth, S.; Zomer, A.; de Jager, V.; Bottacini, F.; van Hijum, S.A.; Mahony, J.; van Sinderen, D. Complete genome of Lactococcus lactis subsp. cremoris UC509.9, host for a model Lactococcal P335 bacteriophage. Genome Announc. 2013, 1, e00119-12. [Google Scholar]
- Dougherty, B.A.; Hill, C.; Weidman, J.F.; Richardson, D.R.; Venter, J.C.; Ross, R.P. Sequence and analysis of the 60 kb conjugative, bacteriocin-producing plasmid pMRC01 from Lactococcus lactis DPC3147. Mol. Microbiol. 1998, 29, 1029–1038. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, C.; Hernández de Rojas, A.; Martínez, B.; Argüelles, M.E.; Suárez, J.E.; Rodríguez, A.; Mayo, B. Nucleotide sequence and analysis of pBL1, a bacteriocin-producing plasmid from Lactococcus lactis IPLA 972. Plasmid 2000, 44, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Gajic, O.; Buist, G.; Kojic, M.; Topisirovic, L.; Kuipers, O.P.; Kok, J. Mechanism of bacteriocin secretion and immunity carried out by lactococcal multidrug resistance proteins. J. Biol. Chem. 2003, 278, 34291–34298. [Google Scholar] [CrossRef] [PubMed]
- Campelo, A.B.; Roces, C.; Mohedano, M.L.; López, P.; Rodríguez, A.; Martínez, B. A bacteriocin gene cluster able to enhance plasmid maintenance in Lactococcus lactis. Microb. Cell Fact. 2014, 13, 77. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.Q.; Dunn, N.W. Genetic analysis of regions involved in replication and cadmium resistance of the plasmid pND302 from Lactococcus lactis. Plasmid 1997, 38, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Fallico, V.; Ross, R.P.; Fitzgerald, G.F.; McAuliffe, O. Novel conjugative plasmids from the natural isolate Lactococcus lactis subspecies cremoris DPC3758, a repository of genes for the potential improvement of dairy starters. J. Dairy Sci. 2012, 95, 3593–3608. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Lu, Y.; Teng, K.L.; Chen, M.L.; Zheng, H.J.; Zhu, Y.Q.; Zhong, J. Complete genome sequence of Lactococcus lactis subsp. Lactis CV56, a probiotic strain isolated from the vaginas of healthy women. J. Bacteriol. 2011, 193, 2886–2887. [Google Scholar]
- Górecki, R.K.; Koryszewska-Bagińska, A.; Gołębiewski, M.; Żylińska, J.; Grynberg, M.; Bardowski, J.K. Adaptative potential of the Lactococcus lactis IL594 strain encoded in its 7 plasmids. PLoS ONE 2011, 6, e22238. [Google Scholar] [CrossRef] [PubMed]
- Wegmann, U.; Overweg, K.; Jeanson, S.; Gasson, M.; Shearman, C. Molecular characterization and structural instability of the industrially important composite metabolic plasmid pLP712. Microbiology 2012, 158, 2936–2945. [Google Scholar] [CrossRef] [PubMed]
- Siezen, R.J.; Renckens, B.; van Swam, I.; Peters, S.; van Kranenburg, R.; Kleerebezem, M.; de Vos, W.M. Complete sequences of four plasmids of Lactococcus lactis subsp. cremoris SK11 reveal extensive adaptation to the dairy environment. Appl. Environ. Microbiol. 2005, 71, 8371–8382. [Google Scholar]
- Tanous, C.; Chambellon, E.; Yvon, M. Sequence analysis of the mobilizable lactococcal plasmid pGdh442 encoding glutamate dehydrogenase activity. Microbiology 2007, 153, 1664–1675. [Google Scholar] [CrossRef] [PubMed]
- Defoor, E.; Kryger, M.B.; Martinussen, J. The orotate transporter encoded by oroP from Lactococcus lactis is required for orotate utilization and has utility as a food-grade selectable marker. Microbiology 2007, 153, 3645–3659. [Google Scholar] [CrossRef] [PubMed]
- Siezen, R.J.; Bayjanov, J.; Renckens, B.; Wels, M.; van Hijum, S.A.; Molenaar, D.; van Hylckama Vlieg, J.E. Complete genome sequence of Lactococcus lactis subsp. lactis KF147, a plant-associated lactic acid bacterium. J. Bacteriol. 2010, 192, 2649–2650. [Google Scholar]
- Fallico, V.; McAuliffe, O.; Fitzgerald, G.F.; Ross, R.P. Plasmids of raw milk cheese isolate Lactococcus lactis subsp. lactis biovar diacetylactis DPC3901 suggest a plant-based origin for the strain. Appl. Environ. Microbiol. 2011, 77, 6451–6462. [Google Scholar]
- Van Kranenburg, R.; Kleerebezem, M.; de Vos, W.M. Nucleotide sequence analysis of the lactococcal EPS plasmid pNZ4000. Plasmid 2000, 43, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Madsen, A.; Josephsen, J. Characterization of LlaCI, a new restriction-modification system from Lactococcus lactis subsp. cremoris W15. Biol. Chem. 1998, 379, 443–449. [Google Scholar] [CrossRef]
- O’Sullivan, D.; Twomey, D.P.; Coffey, A.; Hill, C.; Fitzgerald, G.F.; Ross, R.P. Novel type I restriction specificities through domain shuffling of HsdS subunits in Lactococcus lactis. Mol. Microbiol. 2000, 36, 866–875. [Google Scholar] [CrossRef] [PubMed]
- O’Driscoll, J.; Glynn, F.; Fitzgerald, G.F.; van Sinderen, D. Sequence analysis of the lactococcal plasmid pNP40, a mobile replicon for coping with environmental hazards. J. Bacteriol. 2006, 188, 6629–6639. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, S.; Mahony, J.; van Sinderen, D. The plasmid complement of Lactococcus lactis UC509.9 encodes multiple bacteriophage resistance systems. Appl. Environ. Microbiol. 2014, 80, 4341–4349. [Google Scholar] [CrossRef] [PubMed]
- Christensson, C.; Pillidge, C.J.; Ward, L.J.; O’Toole, P.W. Nucleotide sequence and characterization of the cell envelope proteinase plasmid in Lactococcus lactis subsp. cremoris HP. J. Appl. Microbiol. 2001, 91, 334–343. [Google Scholar] [CrossRef]
- Strahinic, I.; Kojic, M.; Tolinacki, M.; Fira, D.; Topisirovic, L. Molecular characterization of plasmids pS7a and pS7b from Lactococcus lactis subsp. lactis bv. diacetylactis S50 as a base for the construction of mobilizable cloning vectors. J. Appl. Microbiol. 2009, 106, 78–88. [Google Scholar]
- Duan, K.; Liu, C.Q.; Liu, Y.J.; Ren, J.; Dunn, N.W. Nucleotide sequence and thermostability of pND324, a 3.6-kb plasmid from Lactococcus lactis. Appl. Microbiol. Biotechnol. 1999, 53, 36–42. [Google Scholar] [CrossRef] [PubMed]
- O’Driscoll, J.; Glynn, F.; Cahalane, O.; O’Connell-Motherway, M.; Fitzgerald, G.F.; van Sinderen, D. Lactococcal plasmid pNP40 encodes a novel, temperature-sensitive restriction-modification system. Appl. Environ. Microbiol. 2004, 70, 5546–5556. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, P.H.; Mairhofer, J. Marker-free plasmids for biotechnological applications—Implications and perspectives. Trends Biotechnol. 2013, 31, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, K.I.; Larsen, R.; Kibenich, A.; Junge, M.P.; Johansen, E. A food-gradecloning system for industrial strains of Lactococcus lactis. Appl. Environ. Microbiol. 2000, 66, 1253–1258. [Google Scholar] [CrossRef] [PubMed]
- Solem, C.; Defoor, E.; Jensen, P.R.; Martinussen, J. Plasmid pCS1966, a new selection/counterselection tool for lactic acid bacterium strain construction based on the oroP gene, encoding an orotate transporter from Lactococcus lactis. Appl. Environ. Microbiol. 2008, 74, 4772–4775. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Kong, J.; Kong, W. Construction and application of a food-grade expression system for Lactococcus lactis. Mol. Biotechnol. 2013, 54, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Takala, T.M.; Saris, P.E. A food-grade cloning vector for lactic acid bacteriabased on the nisin immunity gene nisI. Appl. Microbiol. Biotechnol. 2002, 59, 467–471. [Google Scholar] [PubMed]
- Mills, S.; Coffey, A.; O’Sullivan, L.; Stokes, D.; Hill, C.; Fitzgerald, G.F.; Ross, R.P. Use of lacticin 481 to facilitate delivery of the bacteriophage resistance plasmid, pCBG104 to cheese starters. J. Appl. Microbiol. 2002, 9, 238–246. [Google Scholar] [CrossRef]
- Coakley, M.; Fitzgerald, G.; Ros, R.P. Application and evaluation of the phageresistance- and bacteriocin-encoding plasmid pMRC01 for the improvementof dairy starter cultures. Appl. Environ. Microbiol. 1997, 6, 1434–1440. [Google Scholar]
- Hayes, F.; Vos, P.; Fitzgerald, G.F.; de Vos, W.M.; Daly, C. Molecular organization of the minimal replicon of novel, narrow-host-range, lactococcal plasmid pCI305. Plasmid 1991, 25, 16–26. [Google Scholar] [CrossRef]
- Kiewiet, R.; Bron, S.; de Jonge, K.; Venema, G.; Seegers, J.F. Theta replication of the lactococcal plasmid pWVO2. Mol. Biol. 1993, 10, 319–327. [Google Scholar] [CrossRef]
- Gravesen, A.; Josephsen, J.; von Wright, A.; Vogensen, F.K. Characterization of the replicon from the lactococcal theta-replicating plasmid pJW563. Plasmid 1995, 34, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Émond, E.; Lavallée, R.; Drolet, G.; Moineau, S.; LaPointe, G. Molecular characterization of a theta replication plasmid and its use for development of a two-component food-grade cloning system for Lactococcus lactis. Appl. Environ. Microbiol. 2001, 67, 1700–1709. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Ogura, M. A novel Bacillus natto plasmid pLS32 capable of replication in Bacillus subtilis. FEBS Lett. 1998, 422, 243–246. [Google Scholar] [CrossRef]
- Kearney, K.; Fitzgerald, G.F.; Seegers, J.F. Identification and characterization of an active plasmid partition mechanism for the novel Lactococcus lactis plasmid pCI2000. J. Bacteriol. 2000, 182, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Del Solar, G.; Giraldo, R.; Ruiz-Echevarría, M.J.; Espinosa, M.; Díaz-Orejas, R. Replication and control of circular bacterial plasmids. Microbiol. Mol. Biol. Rev. 1998, 62, 434–464. [Google Scholar] [PubMed]
- Frère, J.; Benachour, A.; Novel, M.; Novel, G. Identification of the θ-type minimal replicon of the Lactococcus lactis subsp. lactis CNRZ270 lactose protease plasmid pUCL22. Curr. Microbiol. 1993, 27, 97–102. [Google Scholar]
- Frère, J.; Novel, M.; Novel, G. Molecular analysis of the L. lactis subspecies lactis CNRZ270 bidirectional theta replicating lactose plasmid pUCL22. Mol. Microbiol. 1993, 10, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Bolotin, A.; Quinquis, B.; Ehrlich, S.D.; Sorokin, A. Complete genome sequence of Lactococcus lactis subsp. cremoris A76. J. Bacteriol. 2012, 194, 1241–1242. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, L.C.; Saraiva, T.D.L.; Soares, S.C.; Ramos, R.T.J.; Sá, P.H.C.G.; Carneiro, A.R.; Miranda, F.; Freire, M.; Renan, W.; Júnior, A.F.O.; et al. Genome sequence of Lactococcus lactis subsp. lactis NCDO 2118, a GABA-producing strain. Genome Announc. 2014, 2, e00980-14. [Google Scholar]
- De Vos, W.M. Gene cloning and expression in lactic streptococci. FEMS Microbiol. Rev. 1987, 46, 281–295. [Google Scholar] [CrossRef]
- Xu, F.; Pearce, L.E.; Yu, P.L. Molecular cloning and expression of a proteinase gene from Lactococcus lactis subsp. crernoris H2 and construction of a new lactococcal vector pFX1. Arch. Microbiol. 1990, 154, 99–104. [Google Scholar]
- Leenhouts, K.J.; Tolner, B.; Bron, S.; Kok, J.; Venema, G.; Seegers, J.F. Nucleotide sequence and characterization of the broad-host-range lactococcal plasmid pWVO1. Plasmid 1991, 26, 55–66. [Google Scholar] [CrossRef]
- Chang, H.C.; Choi, Y.D.; Lee, H.J. Nucleotide sequence of a plasmid pCL2.1 from Lactococcus lactis ssp. lactis ML8. Plasmid 1995, 34, 234–235. [Google Scholar] [CrossRef] [PubMed]
- Pillidge, C.J.; Cambourn, W.M.; Pearce, L.E. Nucleotide sequence and analysis of pWC1, A pC194-type rolling circle replicon in Lactococcus lactis. Plasmid 1996, 35, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, C.; Mayo, B. Sequence and analysis of pBM02, a novel RCR cryptic plasmid from Lactococcus lactis subsp. cremoris P8-2-47. Plasmid 2003, 49, 118–129. [Google Scholar]
- Raha, A.R.; Hooi, W.Y.; Mariana, N.S.; Radu, S.; Varma, N.R.; Yusoff, K. DNA sequence analysis of a small cryptic plasmid from Lactococcus lactis subsp. lactis M14. Plasmid 2006, 56, 53–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.; Pearce, L.E.; Yu, P.L. Construction of a family of Lactococcal vectors for gene cloning and translational fusion. FEMS Microbiol. Lett. 1991, 77, 55–60. [Google Scholar] [CrossRef]
- De Vos, W.M.; Simons, G.F.M. Gene cloning and expression systems in Lactococci. In Genetics and Biotechnology of Lactic Acid Bacteria, 1st ed.; Gasson, M.J., de Vos, W., Eds.; Springer: Dordrecht, The Netherlands, 1994; pp. 52–105. [Google Scholar]
- Kiewiet, R.; Kok, J.; Seegers, J.F.M.L.; Venema, G.; Bron, S. The mode of replication is a major factor in segregational plasmid instability in Lactococcus lactis. Appl. Environ. Microbiol. 1993, 59, 358–364. [Google Scholar] [PubMed]
- Seegers, J.F.M.L.; Bron, S.; Francke, C.M.; Venema, G.; Kiewiet, R. The majority of lactococcal plasmids carry a highly related replicon. Microbiology 1994, 140, 1291–1300. [Google Scholar] [CrossRef] [PubMed]
- Aguado-Urda, M.; Gibello, A.; Blanco, M.M.; López-Campos, G.H.; Cutuli, M.T.; Fernández-Garayzábal, J.F. Characterization of plasmids in a human clinical strain of Lactococcus garvieae. PLoS ONE 2012, 7, e40119. [Google Scholar] [CrossRef] [PubMed]
- Raccach, M. Pediococci and Biotechnology. Crit. Rev. Microbiol. 1987, 14, 291–309. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, C.F.; Kunka, B.S. Evidence for plasmid linkage of raffinose utilization and associated α-Galactosidase and sucrose hydrolase activity in Pediococcus pentosaceus. Appl. Environ. Microbiol. 1986, 51, 105–109. [Google Scholar] [PubMed]
- Gonzalez, C.F.; Kunka, B.S. Plasmid-associated bacteriocin production and sucrose fermentation in Pediococcus acidilactici. Appl. Environ. Microbiol. 1987, 53, 2534–2538. [Google Scholar] [PubMed]
- Torriani, S.; Vesvovo, M.; Dellaglio, F. Tracing Pediococcus acidilactici in ensiled maize by plasmid-encoded erythromycin resistance. J. Appl. Bacteriol. 1987, 63, 305–309. [Google Scholar] [CrossRef]
- Tankovic, J.; Leclercq, R.; Duval, J. Antimicrobial susceptibility of Pediococcus spp. and genetic basis of macrolide resistance in Pediococcus acidilactici HM3020. Antimicrob. Agents Chemother. 1993, 37, 789–792. [Google Scholar] [CrossRef] [PubMed]
- Daeschel, M.A.; Klaenhammer, T.R. Association of a 13.6-Megadalton plasmid in Pediococcus pentosaceus with bacteriocin activity. Appl. Environ. Microbiol. 1985, 50, 1538–1541. [Google Scholar] [PubMed]
- Marugg, J.D.; Gonzalez, C.F.; Kunka, B.S.; Ledeboe, A.M.; Pucci, M.J.; Toonen, M.Y.; Walker, S.A.; Zoetmulder, L.C.; Vandenburgh, P.A. Cloning, expression, and nucleotide sequence of genes involved in production of pediocin PA-1, and bacteriocin from Pediococcus acidilactici PAC1.0. Appl. Environ. Microbiol. 1992, 58, 2360–2367. [Google Scholar] [PubMed]
- Motlagh, A.; Bukhtiyarova, M.; Ray, B. Complete nucleotide sequence of pSMB 74, a plasmid encoding the production of pediocin AcH in Pediococcus acidilactici. Lett. Appl. Microbiol. 1994, 18, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Kantor, A.; Montville, T.J.; Mett, A.; Shapira, R. Molecular characterization of the replicon of the Pediococcus pentosaceus 43200 pediocin A plasmid pMD136. FEMS Microbiol. Lett. 1997, 151, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Giacomini, A.; Squartini, A.; Nuti, M.P. Nucleotide sequence and analysis of plasmid pMD136 from Pediococcus pentosaceus FBB61 (ATCC43200) involved in pediocin A production. Plasmid 2000, 43, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.H.; Zhang, C.; Wang, Y.F.; Shi, J.; Zhang, L.W.; Ding, Z.Q.; Qu, X.J.; Cui, H.Y. Class IIa bacteriocins, diversity and new developments. Int. J. Mol. Sci. 2012, 13, 16668–16707. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, E.B.; O’Sullivan, O.; Stanton, C.; Danielsen, M.; Simpson, P.J.; Callanan, M.J.; Ross, R.P.; Hill, C. pEOC01, A plasmid from Pediococcus acidilactici which encodes an identical streptomycin resistance (aadE) gene to that found in Campylobacter jejuni. Plasmid 2007, 58, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Alegre, M.T.; Rodríguez, M.C.; Mesas, J.M. Nucleotide sequence, structural organization and host range of pRS4, a small cryptic Pediococcus entosaceus plasmid that contains two cassettes commonly found in other lactic acid bacteria. FEMS Microbiol. Lett. 2005, 250, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Alegre, M.T.; Rodríguez, M.C.; Mesas, J.M. Characterization of pRS5, a theta-type plasmid found in a strain of Pediococcus pentosaceus isolated from wine that can be used to generate cloning vectors for lactic acid bacteria. Plasmid 2009, 61, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Ray, B.; Motlagh, A.M.; Johnson, M.C.; Bozoglu, F. Mapping of pSMB74, a plasmid encoding bacteriocin AcH production (Pap+) trait in Pediococcus acidilactici H. Lett. Appl. Microbiol. 1992, 15, 35–37. [Google Scholar] [CrossRef]
- Motlagh, A.M.; Bhunia, A.K.; Szostek, F.; Hansen, T.R.; Johnson, M.C.; Ray, B. Nucleotide and amino acid sequence of pap-gene (pediocin AcH production) in Pediococcus acidilactici H. Lett. Appl. Microbiol. 1992, 15, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Schved, F.; Lalazar, A.; Henis, Y.; Juven, B.J. Purification, partial characterization and plasmid linkage of pediocin SJ-1, a bacteriocin produced by Pediococcus acidilactici. J. Appl. Bacteriol. 1993, 74, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.M.; Cintas, L.M.; Casaus, P.; Martínez, M.I.; Suárez, A.; Hernández, P.E. Detection of pediocin PA-1 producing pediococci by rapid molecular producing by rapid molecular biology techniques. Food Microbiol. 1997, 14, 363–371. [Google Scholar] [CrossRef]
- Miller, K.W.; Ray, P.; Steinmetz, T.; Hanekamp, T.; Ray, B. Gene organization and sequences of pediocin AcH/PA-1 production operons in Pediococcus and Lactobacillus plasmids. Lett. Appl. Microbiol. 2005, 40, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Ennahar, S.; Aoude-Werner, D.; Sorokine, O.; van Dorsselaer, A.; Bringel, F.; Hubert, J.C.; Hasselmann, C. Production of pediocin AcH by Lactobacillus plantarum WHE92 isolated from cheese. Appl. Environ. Microbiol. 1996, 62, 4381–4387. [Google Scholar] [PubMed]
- Bernbom, N.; Licht, T.R.; Saadbye, P.; Vogensen, F.K.; Norrung, B. Lactobacillus plantarum inhibits growth of Listeria monocytogenes in an in vitro continuous flow gut model, but promotes invasion of L. monocytogenes in the gut of gnotobiotic rats. Int. J. Food Microbiol. 2006, 108, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Devi, S.M.; Halami, P.M. Detection of mobile genetic elements in pediocin PA-1 like producing lactic acid bacteria. J. Basic Microbiol. 2013, 53, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Devi, S.M.; Ramaswamy, A.M.; Halami, P.M. In situ production of pediocin PA-1 like bacteriocin by different genera of lactic acid bacteria in soymilk fermentation and evaluation of sensory properties of the fermented soy curd. J. Food Sci. Technol. 2014, 51, 3325–3332. [Google Scholar]
- Le Marrec, C.; Hyronimus, B.; Bressollier, P.; Verneuil, B.; Urdaci, M.C. Biochemical and genetic characterization of coagulin, a new antilisterial bacteriocin in the pediocin family of bacteriocins, produced by Bacillus coagulans I4. Appl. Environ. Microbiol. 2000, 66, 5213–5220. [Google Scholar] [CrossRef] [PubMed]
- Ray, S.K.; Johnson, M.C.; Ray, B. Bacteriocin plasmids of Pediococcus acidilactici. J. Ind. Microbiol. 1989, 4, 163–171. [Google Scholar] [CrossRef]
- Jager, K.; Harlander, S. Characterization of a bacteriocin from Pediococcus acidilactici PC and comparison of bacteriocin-producing strains using molecular typing procedures. Appl. Environ. Microbiol. 1992, 37, 631–637. [Google Scholar] [CrossRef]
- Bhunia, A.K.; Bhowmik, T.K.; Johnson, M.G. Determination of bacteriocin-encoding plasmids of Pediococcus acidilactici strains by southern hybridization. Lett. Appl. Microbiol. 1994, 18, 168–170. [Google Scholar] [CrossRef]
- Pittet, V.; Abegunde, T.; Marfleet, T.; Haakensen, M.; Morrow, K.; Jayaprakash, T.; Schroeder, K.; Trost, B.; Byrns, S.; Bergsveinson, J.; et al. Complete genome sequence of the beer spoilage organism ATCC BAA-344T. J. Bacteriol. 2012, 194, 1271–1272. [Google Scholar] [CrossRef] [PubMed]
- Klein, G. Taxonomy, ecology and antibiotic resistance of enterococci from food and gastro-intestinal tract. Review. Int. J. Food Microbiol. 2003, 88, 123–131. [Google Scholar] [CrossRef]
- Foulquié Moreno, M.R.; Sarantinopoulos, P.; Tsakalidou, E.; de Vuyst, L. The role and application of enterococci in food and health. Int. J. Food Microbiol. 2006, 106, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Poeta, P.; Costa, D.; Rodrigues, J.; Torres, C. Antimicrobial resistance and the mechanisms implicated in faecalis enterococci from healthy humans, poultry and pets in Portugal. Int. J. Antimicrob. Agents 2006, 27, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Clewell, D.B.; Yagi, Y.; Dunny, G.M.; Schultz, S.K. Characterization of three plasmid deoxyribonucleic acid molecules in a strain of Streptococcus faecalis, identification of a plasmid determining erythromycin resistance. J. Bacteriol. 1974, 117, 283–289. [Google Scholar] [PubMed]
- Francia, M.V.; Clewell, D.B. Amplification of the tetracycline resistance determinant of pAMalpha1 in Enterococcus faecalis requires a site-specific recombination event involving relaxase. J. Bacteriol. 2002, 184, 5187–5193. [Google Scholar] [CrossRef] [PubMed]
- Tanimoto, K.; Ike, Y. Complete nucleotide sequencing and analysis of the 65-kb highly conjugative Enterococcus faecium plasmid pMG1, identification of the transfer-related region and the minimum region required for replication. FEMS Microbiol. Lett. 2008, 288, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Sletvold, H.; Johnsen, P.J.; Hamre, I.; Simonsen, G.S.; Sundsfjord, A.; Nielsen, K.M. Complete sequence of Enterococcus faecium pVEF3 and the detection of an omega-epsilon-zeta toxin-antitoxin module and an ABC transporter. Plasmid 2008, 60, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Zischka, M.; Kuenne, C.; Blom, J.; Dabrowski, P.W.; Linke, B.; Hain, T.; Nitsche, A.; Goesmann, A.; Larsen, J.; Jensen, L.B.; et al. Complete genome sequence of the porcine isolate Enterococcus faecalis D32. J. Bacteriol. 2012, 194, 5490–5491. [Google Scholar] [CrossRef] [PubMed]
- Balla, E.; Dicks, L.M. Molecular analysis of the gene cluster involved in the production and secretion of enterocins 1071A and 1071B and of the genes responsible for the replication and transfer of plasmid pEF1071. Int. J. Food Microbiol. 2005, 99, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Barba, J.L.; Floriano, B.; Maldonado-Barragan, A.; Jimenez-Diaz, R. Molecular analysis of the 21-kb bacteriocin-encoding plasmid pEF1from Enterococcus faecium 6T1a. Plasmid 2007, 57, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Criado, R.; Diep, D.B.; Aakra, Å.; Gutiérrez, J.; Nes, I.F.; Hernández, P.E.; Cintas, L.M. Complete sequence of the enterocin Q-encoding plasmid pCIZ2 from the multiple bacteriocin producer Enterococcus faecium L50 and genetic characterization of enterocin Q production and immunity. Appl. Environ. Microbiol. 2006, 72, 6653–6666. [Google Scholar] [PubMed]
- Yamashita, H.; Tomita, H.; Inoue, T.; Ike, Y. Genetic organization and mode of action of a novel bacteriocin, bacteriocin 51, determinant of VanA-type vancomycin-resistant Enterococcus faecium. Antimicrob. Agents Chemother. 2011, 55, 4352–4360. [Google Scholar] [CrossRef] [PubMed]
- Lam, M.M.; Seemann, T.; Bulach, M.; Gladman, S.L.; Chen, H.; Haring, V.; Moore, R.J.; Ballard, S.; Grayson, M.L.; Johnson, P.D.; et al. Comparative analysis of the first complete Enterococcus faecium genome. J. Bacteriol. 2012, 194, 2334–2341. [Google Scholar] [CrossRef] [PubMed]
- De Boever, E.H.; Clewell, D.B.; Fraser, C.M. Enterococcus faecalis conjugative plasmid pAM373, complete nucleotide sequence and genetic analyses of sex pheromone response. Mol. Microbiol. 2002, 37, 1327–1341. [Google Scholar] [CrossRef]
- Paulsen, I.; Banerjei, L.; Myers, G.; Nelson, K.; Seshadri, R.; Read, T.D.; Fouts, D.E.; Eisen, J.A.; Gill, S.R.; Heidelberg, J.F.; et al. Role of mobile DNA in the evolution of vancomycin-resistant Enterococcus faecalis. Science 2003, 299, 2071–2074. [Google Scholar] [CrossRef] [PubMed]
- Tendolkar, P.M.; Baghdayan, A.S.; Shankar, N. Putative surface proteins encoded within a novel transferable locus confer a high-biofilm phenotype to Enterococcus faecalis. J. Bacteriol. 2006, 188, 2063–2072. [Google Scholar] [CrossRef] [PubMed]
- Brede, D.A.; Snipen, L.G.; Ussery, D.W.; Nederbragt, A.J.; Nes, I.F. Complete genome sequence of the commensal Enterococcus faecalis 62, isolated from a healthy Norwegian infant. J. Bacteriol. 2011, 193, 2377–2378. [Google Scholar] [CrossRef] [PubMed]
- Hirt, H.; Manias, D.A.; Bryan, E.M.; Klein, J.R.; Marklund, J.K.; Staddon, J.H.; Paustian, M.L.; Kapur, V.; Dunny, G.M. Characterization of the pheromone response of the Enterococcus faecalis conjugative plasmid pCF10, complete sequence and comparative analysis of the transcriptional and phenotypic responses of pCF10-containing cells to pheromone induction. J. Bacteriol. 2005, 187, 1044–1054. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Foulquié-Moreno, M.R.; Revets, H. Screening for enterocins and detection of hemolysin and vancomycin resistance in enterococci of different origins. Int. J. Food Microbiol. 2003, 84, 299–318. [Google Scholar] [CrossRef]
- Maidin, M.S.T.; Song, A.A.L.; Jalilsood, T.; Sieo, C.C.; Yusoff, K.; Rahim, R.A. Construction of a novel inducible expression vector for Lactococcus lactis M4 and Lactobacillus plantarum Pa21. Plasmid 2014, 74, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Mercenier, A. Molecular genetics of Streptococcus thermophilus. FEMS Microbiol. Rev. 1990, 87, 61–78. [Google Scholar] [CrossRef]
- Somkuti, G.A.; Solaiman, D.K.Y.; Steinberg, D.H. Structural and functional properties of the hsp16.4-bearing plasmid pER341 in Streptococcus thermophilus. Plasmid 1998, 40, 61–72. [Google Scholar]
- O’Sullivan, T.; van Sinderen, D.; Fitzgerald, G. Structural and functional analysis of pCI65st, a 6.5 kb plasmid from Streptococcus thermophilus NDI-6. Microbiology 1999, 145, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Su, P.; Jury, K.; Allison, G.E.; Wong, W.Y.; Kim, W.S.; Liu, C.Q.; Vancov, T.; Dunn, N.W. Cloning vectors for Streptococcus thermophilus derived from a native plasmid. FEMS Microbiol. Lett. 2002, 216, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Geis, A.E.; Demerdash, H.A.M.; Heller, K.J. Sequence analysis and characterization of plasmids from Streptococcus thermophilus. Plasmid 2003, 50, 53–69. [Google Scholar] [CrossRef]
- Petrova, P.; Miteva, V.; Ruiz-Maso, J.A.; del Solar, G. Structural and functional analysis of pt38, a 2.9 kb plasmid of Streptococcus thermophilus yoghurt strain. Plasmid 2003, 50, 176–189. [Google Scholar] [CrossRef]
- Somkuti, G.A.; Steinberg, D.H. Promoter activity of the pER341-borne STphsp in heterologous gene expression in E. coli and Streptococcus thermophilus. FEMS Microbiol. Lett. 1999, 179, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Solow, B.T.; Somkuti, G.A. Comparison of low-molecular-weight heat stress proteins encoded on plasmids in different strains of Streptococcus thermophilus. Curr. Microbiol. 2000, 41, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.; Slesarev, A.; Wolf, Y.; Sorokin, A.; Mirkin, B.; Koonin, E.; Pavlov, A.; Pavlova, N.; Karamychev, V.; Polouchine, N.; et al. Comparative genomics of the lactic acid bacteria. Proc. Natl. Acad. Sci. USA 2006, 103, 15611–15616. [Google Scholar] [CrossRef] [PubMed]
- Turgeon, N.; Moineau, S. Isolation and characterization of a Streptococcus thermophilus plasmid closely related to the pMV158 family. Plasmid 2001, 45, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Girard, S.L.; Moineau, S. Analysis of two theta-replicating plasmids of Streptococcus thermophilus. Plasmid 2007, 58, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Schleifer, K.H.; Ludwig, W. Phylogenetic relationships of lactic acid bacteria. In The Genera of Lactic Acid Bacteria, 1st ed.; Wood, B.J.B., Holzapfel, W.H., Eds.; Springer US: New York, NY, USA, 1995; Volume 2, pp. 7–18. [Google Scholar]
- Sgorbati, B.; Biavati, B.; Palenzona, D. The genus Bifidobacterium. In The Genera of Lactic Acid Bacteria, 1st ed.; Wood, B.J.B., Holzapfel, W.H., Eds.; Springer US: New York, NY, USA, 1995; Volume 2, pp. 279–306. [Google Scholar]
- Russell, D.A.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C. Metabolic activities and probiotic potential of bifidobacteria. Int. J. Food Microbiol. 2011, 149, 88–105. [Google Scholar] [CrossRef] [PubMed]
- Sgorbati, B.; Scardovi, V.; Leblanc, D.J. Related structures in the plasmid profiles of Bifidobacterium asteroides, B. indicum and B. globosum. Microbiologica 1986, 9, 443–454. [Google Scholar]
- Sgorbati, B.; Scardovi, V.; Leblanc, D.J. Related structures in the plasmid profiles of Bifidobacterium longum. Microbiologica 1986, 9, 415–422. [Google Scholar] [PubMed]
- Iwata, M.; Morishita, T. The presence of plasmids in Bifidobacterium breve. Lett. Appl. Microbiol. 2008, 9, 165–168. [Google Scholar] [CrossRef]
- Shkoporov, A.N.; Efimov, B.A.; Khokhlova, E.V.; Steele, J.L.; Kafarskaia, L.I.; Smeianov, V.V. Characterization of plasmids from human infant Bifidobacterium strains, sequence analysis and construction of E. coli-Bifidobacterium shuttle vectors. Plasmid 2008, 60, 136–148. [Google Scholar]
- Álvarez-Martín, P.; BelénFlórez, A.; Mayo, B. Screening for plasmids among human bifidobacteria species: Sequencing and analysis of pBC1 from Bifidobacterium catenulatum L48. Plasmid 2007, 57, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Martín, P.; O’Connell-Motherway, M.; van Sinderen, D.; Mayo, B. Functional analysis of the pBC1 replicon from Bifidobacterium catenulatum L48. Appl. Microbiol. Biotechnol. 2007, 76, 1395–1402. [Google Scholar] [CrossRef] [PubMed]
- Park, M.S.; Shin, D.W.; Lee, K.H.; Ji, G.E. Sequence analysis of plasmid pKJ50 from Bifidobacterium longum. Microbiology 1999, 145, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Schell, M.A.; Karmirantzou, M.; Snel, B.; Vilanova, D.; Berger, B.; Pessi, G.; Zwahlen, M.C.; Desiere, F.; Bork, P.; Delley, M.; et al. The genome sequence of Bifidobacterium Longum reflects its adaptation to the human gastrointestinal tract. Proc. Natl. Acad. Sci. USA 2002, 99, 14422–14427. [Google Scholar] [CrossRef]
- Corneau, N.E.; Emond, G.; LaPointe, G. Molecular characterizationof three plasmids from Bifidobacterium longum. Plasmid 2004, 51, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Samura, K.; Kano, Y. Structural and functional analysis of pTB6 from Bifidobacterium longum. Biosci. Biotechnol. Biochem. 2005, 69, 422–425. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; O’Sullivan, D.J. Sequence analysis of two cryptic plasmids from Bifidobacterium longum DJO10A and construction of a shuttle cloning vector. Appl. Environ. Microbiol. 2006, 72, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Moon, G.S.; Wegmann, U.; Gunning, A.P.; Gasson, M.J.; Narbad, A. Isolation and characterization of a theta-type cryptic plasmid from Bifidobacterium longum FI10564. J. Microbiol. Biotechnol. 2009, 19, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Ham, J.S.; Lee, T.; Byun, M.J.; Lee, K.T.; Kim, M.K.; Han, G.S.; Jeong, S.-G.; Oh, M.-H.; Kim, D.-H.; Kim, H. Complete genome sequence of Bifidobacterium longum subsp. longum KACC 91563. J. Bacteriol. 2011, 193, 5044. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, S.; Toh, H.; Hase, K.; Oshima, K.; Nakanishi, Y.; Yoshimura, K.; Tobe, T.; Clarke, J.M.; Topping, D.L.; Suzuki, T.; et al. Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature 2011, 469, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Martín, P.; Życka-Krzesińska, J.; Bardowski, J.; Mayo, B. Sequence analysis of plasmid pSP02 from Bifidobacterium longum M62 and construction of pSP02-derived cloning vectors. Plasmid 2013, 69, 119–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, M.; Svenson, C.; Bowling, D.; Glenn, D. Complete nucleotide sequence of a native plasmid from Brevibacterium linens. Plasmid 2003, 49, 160–168. [Google Scholar] [CrossRef]
- Sangrador-Vegas, A.; Stanton, C.; van Sinderen, D.; Fitzgerald, G.F.; Ross, R.P. Characterization of plasmid pASV479 from Bifidobacterium pseudolongum subsp. globosum and its use for expression vector construction. Plasmid 2007, 58, 140–147. [Google Scholar]
- Gibbs, M.J.; Smeianov, V.V.; Steele, J.L.; Upcroft, P.; Efimov, B.A. Two families of Rep-like genes that probably originated by interspecies recombination are represented in viral, plasmid, bacterial, and parasitic protozoan genomes. Mol. Biol. Evol. 2006, 23, 1097–1100. [Google Scholar] [CrossRef] [PubMed]
- Spano, G.; Massa, S. Environmental stress response in wine lactic acid bacteria: Beyond Bacillus subtilis. Crit. Rev. Microbiol. 2006, 32, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Capozzi, V.; Russo, P.; Beneduce, L.; Weidmann, S.; Grieco, F.; Guzzo, J.; Spano, G. Technological properties of Oenococcus oeni strains isolated from typical southern italian wines. Lett. Appl. Microbiol. 2010, 50, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Versari, A.; Parpinello, G.P.; Cattaneo, M. Leuconostoc oenos and malolactic fermentation in wine, a review. J. Ind. Microbiol. Biotechnol. 1999, 23, 447–455. [Google Scholar] [CrossRef]
- Fremaux, C.; Aigle, M.; Lonvaud-Funel, A. Sequence analysis of Leuconostoc oenos DNA, organization of pLo13, a cryptic plasmid. Plasmid 1993, 30, 212–223. [Google Scholar] [CrossRef] [PubMed]
- Prévost, H.; Cavin, J.F.; Lamoreux, M.; Diviès, C. Plasmid and chromosome characterization of Leuconostoc oenos strains. Am. J. Enol. Vitic. 1995, 46, 43–48. [Google Scholar]
- Brito, L.; Vieira, G.; Santos, M.A.; Paveia, H. Nucleotide sequence analysis of pOg32, a cryptic plasmid from Leuconostoc oenos. Plasmid 1996, 36, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Zúñiga, M.; Pardo, I.; Ferrer, S. Nucleotide sequence of plasmid p4028, a cryptic plasmid from Leuconostoc oenos. Plasmid 1996, 35, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Alegre, M.T.; Rodríguez, M.C.; Mesas, J.M. Nucleotide sequence analysis of pRS1, a cryptic plasmid from Oenococcus oeni. Plasmid 1999, 41, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Mesas, J.M.; Alegre, M.T. Plasmids from wine-related lactic acid bacteria. In Biology of Microorganisms on Grapes, in Must and in Wine, 1st ed.; König, H., Unden, G., Fröhlich, G., Eds.; Springer-Verlag Berlin Heidelberg: Heidelberg, Germany, 2009; pp. 415–428. [Google Scholar]
- Favier, M.; Bilhère, E.; Lonvaud-Funel, A.; Moine, V.; Lucas, P.M. Identification of pOENI-1 and related plasmids in Oenococcus oeni strains performing the malolactic fermentation in wine. PLoS ONE 2012, 7, e49082. [Google Scholar] [CrossRef] [PubMed]
- Beltramo, C.; Oraby, M.; Bourel, G.; Garmynb, D.; Guzzob, J. A new vector, pGID052, for genetic transfer in Oenococcus oeni. FEMS Microbiol. Lett. 2004, 236, 53–60. [Google Scholar] [PubMed]
- Mesas, J.M.; Rodríguez, M.C.; Alegre, M.T. Nucleotide sequence analysis of pRS2 and pRS3, two small cryptic plasmids from Oenococcus oeni. Plasmid 2001, 46, 149–151. [Google Scholar] [CrossRef] [PubMed]
- Borneman, A.R.; McCarthy, J.M.; Chambers, P.J.; Bartowsky, E. Comparative analysis of the Oenococcus oeni pan genome revealsgenetic diversity in industrially-relevant pathways. BMC Genomics 2012, 13, 373. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, M.C.; Alegre, M.T.; Martín, M.C.; Mesas, J.M. The use of the replication region of plasmid pRS7 from Oenococcus oeni as a putative tool to generate cloning vectorsfor lactic acid bacteria. Plasmid 2015, 77, 28–31. [Google Scholar] [CrossRef] [PubMed]
- Endo, A.; Okada, S. Oenococcus kitaharae sp. nov., a non-acidophilic and nonmalolactic-fermenting Oenococcus isolated from a composting distilled shochu residue. Int. J. Syst. Evol. Microbiol. 2006, 56, 2345–2348. [Google Scholar] [CrossRef] [PubMed]
- Borneman, A.R.; McCarthy, J.M.; Chambers, P.J.; Bartowsky, E.J. Functional divergence in the genus Oenococcus as predicted by genome sequencing of the newly-described species, Oenococcus kitaharae. PLoS ONE 2012, 7, e29626. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.A. Plasmid rolling-circle replication, highlights of two decades or research. Plasmid 2005, 53, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.A. Rolling-circle replication of bacterial plasmids. Microbiol. Mol. Biol. Rev. 1997, 61, 442–455. [Google Scholar] [PubMed]
- Kornberg, A.; Baker, T.A. DNA Replication, 2nd ed.; William, H., Ed.; Freeman and Company: New York, NY, USA, 1992; pp. 637–674. [Google Scholar]
- Bruand, C.; Le Chatelier, E.; Ehrlich, S.D.; Janniere, L. A fourth class of theta-replicating plasmids, the pAMβ1 family from Gram-positive bacteria. Proc. Natl. Acad. Sci. USA 1993, 90, 11668–11672. [Google Scholar] [CrossRef] [PubMed]
- Meijer, W.J.; de Boer, A.J.; van Tongeren, S.; Venema, G.; Bron, S. Characterization of the replication region of the Bacillus subtilis plasmid pLS20, a novel type of replicon. Nucleic Acids Res. 1995, 23, 3214–3223. [Google Scholar] [CrossRef] [PubMed]
- Brantl, S.D.; Behnke, D.; Alonso, J.C. Molecular analysis of the replication region of the conjugative Streptococcus agalactiae plasmid pIP501 in Bacillus subtilis. Comparison with plasmids pAMb1 and pSM19035. Nucleic Acids Res. 1990, 18, 4783–4790. [Google Scholar] [CrossRef] [PubMed]
- Takiguchi, R.; Hashiba, H.; Aoyama, K.; Ishii, S. Complete nucleotide sequence and characterization of a cryptic plasmid from Lactobacillus helveticus subsp. jugurti. Appl. Environ. Microbiol. 1989, 55, 1653–1655. [Google Scholar]
- Heath, D.G.; An, F.Y.; Weaver, K.E.; Clewell, D. Phase variation of Enterococcus faecalis pAD1 conjugation functions relates to changes in iteron sequence region. J. Bacteriol. 1995, 177, 5453–5459. [Google Scholar] [PubMed]
- Weaver, K.E.; Clewell, D.B.; An, F. Identification, characterization, and nucleotide sequence of a region of Enterococcus faecalis pheromone-responsive plasmid pAD1 capable of autonomous replication. J. Bacteriol. 1993, 175, 1900–1909. [Google Scholar] [PubMed]
- Hedberg, P.J.; Leonard, B.A.; Ruhfel, R.E.; Dunny, G.M. Identification and characterization of the genes of Enterococcus faecalis plasmid pCF10 involved in replication and in negative control of pheromone-inducible conjugation. Plasmid 1996, 35, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, S.; Tomita, H.; Wakamatsu, E.; Tanimoto, K.; Ike, Y. Physical mapping of the conjugative bacteriocin plasmid pPD1 of Enterococcus faecalis and identification of the determinant related to the pheromone response. J. Bacteriol. 1995, 177, 5574–5581. [Google Scholar] [PubMed]
- Francia, M.V.; Varsaki, A.; Garcillan-Barcia, M.P.; Latorre, A.; Drainas, C.; de la Cruz, F. A classification scheme for mobilization regions of bacterial plasmids. FEMS Microbiol. Rev. 2004, 28, 79–100. [Google Scholar] [CrossRef] [PubMed]
- Verraes, C.; van Boxstael, S.; van Meervenne, E.; van Coillie, E.; Butaye, P.; Catry, B.; de Schaetzen, M.A.; van Huffel, X.; Imberechts, H.; Dierick, K. Antimicrobial resistance in the food chain: A review. Int. J. Environ. Res. Public Health 2013, 10, 2643–2669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smillie, C.; Garcillán-Barcia, M.P.; Francia, M.V.; Rocha, E.P.C.; de la Cruz, F. Mobility of plasmids. Microbiol. Mol. Biol. Rev. 2010, 74, 434–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grohmann, E.; Muth, G.; Espinosa, M. Conjugative plasmid transfer in gram-positive bacteria. Microbiol. Mol. Biol. Rev. 2003, 67, 277–301. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo-Díaz, F.; Fernández-Lόpez, C.; Garcillán-Barcia, M.P.; Espinosa, M. Bringing them together: Plasmid pMV158 rolling circle replication and conjugation under an evolutionary perspective. Plasmid 2014, 74, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Fernández-López, C.; Bravo, A.; Ruiz-Cruz, S.; Solano-Collado, V.; Garsin, D.A.; Lorenzo-Díaz, F.; Espinosa, M. Mobilizable rolling-circle replicating plasmids from Gram-positive bacteria: Alow-cost conjugative transfer. Microbiol. Spectr. 2014, 2, 8. [Google Scholar] [CrossRef] [PubMed]
- Garcillán-Barcia, M.P.; Francia, M.V.; de la Cruz, F. Thediversity of conjugative relaxases and its application in plasmid classification. FEMS Microbiol. Rev. 2009, 33, 657–687. [Google Scholar] [CrossRef] [PubMed]
- Sano, K.; Otani, M.; Okada, Y.; Kawamura, R.; Umesaki, M.; Ohi, Y.; Umezawa, C.; Kanatani, K. Identification of the replication region of the Lactobacillus acidophilus plasmid pLA106. FEMS Microbiol. Lett. 1997, 148, 223–236. [Google Scholar] [CrossRef] [PubMed]
- De las Rivas, B.; Marcobal, A.; Muñoz, R. Complete nucleotide sequence and structural organization of pPB1, a small Lactobacillus plantarum cryptic plasmid that originated by modular exchange. Plasmid 2004, 52, 203–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudhamani, M.; Ismaiel, E.; Geis, A.; Batish, V.; Heller, K.J. Characterisation of pSMA23, a 3.5 kbp plasmid of Lactobacillus casei, and application for heterologous expression in Lactobacillus. Plasmid 2008, 59, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Z.; Hao, Y.; Yin, S.; Luan, C. Characterization of a novel rolling-circle replication plasmid pYSI8 from Lactobacillus sakei YSI8. Plasmid 2009, 62, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Vujcic, M.; Topisirovic, L. Molecular analysis of the rolling-circle replicating plasmid pA1 of Lactobacillus plantarum A112. Appl. Environ. Microbiol. 1993, 59, 274–280. [Google Scholar] [PubMed]
- Skaugen, M. The complete nucleotide sequence of a small cryptic plasmid from Lactobacillus plantarum. Plasmid 1989, 22, 175–179. [Google Scholar] [CrossRef]
- Malik, S.; Siezen, R.J.; Renckens, B.; Vaneechoutte, M.; Vanderleyden, J.; Lebeer, S. Draft genome sequence of Lactobacillus plantarum CMPG5300, a human vaginal isolate. Genome Announc. 2014, 2, e01149-14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Sun, Z.; Bilige, M.; Zhang, H. Complete genome sequence of probiotic Lactobacillus plantarum P-8 with antibacterial activity. J. Biotechnol. 2015, 193, 41–42. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.M.; Wang, Y.Y.; Wang, Z.L.; Cui, J.; Lan, H.K.; Zhou, J.Y. Complete DNA sequence and analysis of two cryptic plasmids isolated from Lactobacillus plantarum. Plasmid 2003, 50, 70–73. [Google Scholar]
- Li, X.; Gu, Q.; Lou, X.; Zhang, X.; Song, D.; Shen, L.; Zhao, Y. Complete genome sequence of the probiotic Lactobacillus plantarum strain ZJ316. Genome Announc. 2013, 1, e0009413. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhai, Z.; Yin, S.; Huang, Y.; Wang, Q.; Luo, Y.; Hao, Y. Characterization of a rolling-circle replication plasmid pLR1 from Lactobacillus plantarum LR1. Curr. Microbiol. 2009, 58, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Olympia, M.; Fukuda, H.; Ono, H.; Kaneko, Y.; Takano, M. Characterization of starch-hydrolyzing lactic acid bacteria isolated from a fermented fish and rice food, “BurongIsda”, and its amylolytic enzyme. J. Ferment. Bioeng. 1995, 80, 124–130. [Google Scholar] [CrossRef]
- Kaneko, Y.; Kobayashi, H.; Kiatpapan, P.; Nishimoto, T.; Napitupulu, R.; Ono, H.; Murooka, Y. Development of a host-vector system for Lactobacillus plantarum L137 isolated from a traditional fermented food produced in the Philippines. J. Biosci. Bioeng. 2000, 89, 62–67. [Google Scholar] [CrossRef]
- Van Reenen, C.A.; van Zyl, W.H.; Dicks, L.M. Expression of the immunity protein of plantaricin 423, produced by Lactobacillus plantarum 423, and analysis of the plasmid encoding the bacteriocin. Appl. Environ. Microbiol. 2006, 72, 7644–7651. [Google Scholar] [CrossRef] [PubMed]
- Bouia, A.; Bringel, F.; Frey, L.; Kammerer, B.; Belarbi, A.; Guyonvarch, A.; Hubert, J.C. Structural organization of pLP1, a cryptic plasmid from Lactobacillus plantarum CCM 1904. Plasmid 1989, 22, 185–192. [Google Scholar] [CrossRef]
- Bates, E.E.; Gilbert, H.J. Characterization of a cryptic plasmid from Lactobacillus plantarum. Gene 1989, 85, 253–258. [Google Scholar] [CrossRef]
- Leer, R.J.; van Luijk, N.; Posno, M.; Pouwels, P.H. Structural and functional analysis of two cryptic plasmids from Lactobacillus pentosus MD353 and Lactobacillus plantarum ATCC 8014. Mol. Gen. Genet. 1992, 234, 265–274. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, D.; Ross, R.P.; Twomey, D.P.; Fitzgerald, G.F.; Hill, C.; Coffey, A. Naturally occurring lactococcal plasmid pAH90 links bacteriophage resistance and mobility functions to a food-grade selectable marker. Appl. Environ. Microbiol. 2001, 67, 929–937. [Google Scholar] [CrossRef] [PubMed]
- Lucey, M.; Daly, C.; Fitzgerald, G. Identification and sequence analysis of the replication region of the phage resistance plasmid pCI528 from Lactococcus lactis subsp. cremoris UC503. FEMS Microbiol. Lett. 1993, 110, 249–256. [Google Scholar] [CrossRef]
- Kobayashi, M.; Nomura, M.; Kimoto, H. Manipulation for plasmid elimination by transforming synthetic competitors diversifies Lactococcus lactis starters applicable to food products. Biosci. Biotechnol. Biochem. 2007, 71, 2647–2654. [Google Scholar] [CrossRef] [PubMed]
- Gravesen, A.; von Wright, A.; Josephsen, J.; Vogensen, F.K. Replication regions of two pairs of incompatible lactococcal Theta plasmids. Plasmid 1997, 38, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Schouler, C.; Gautier, M.; Ehrlich, S.D.; Chopin, M.C. Combinational variation of restriction modification specificities in Lactococcus lactis. Mol. Microbiol. 1998, 28, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Anba, J.; Bidnenko, E.; Hillier, A.; Ehrlich, D.; Chopin, M.C. Characterization of the lactococcal abiD1 gene coding for phage abortive infection. J. Bacteriol. 1995, 177, 3818–3823. [Google Scholar] [PubMed]
- Schouler, C.; Clier, F.; Lerayer, A.L.; Ehrlich, S.D.; Chopin, M.C. A type IC restriction-modification system in Lactococcus lactis. J. Bacteriol. 1998, 180, 407–411. [Google Scholar] [PubMed]
- Perreten, V.; Schwarz, F.V.; Teuber, M.; Levy, S.B. Mdt(A), a new efflux protein conferring multiple antibiotic resistance in Lactococcus lactis and Escherichia coli. Antimicrob. Agents Chemother. 2001, 45, 1109–1114. [Google Scholar] [CrossRef] [PubMed]
- Kojic, M.; Jovcic, B.; Strahinic, I.; Begovic, J.; Lozo, J.; Veljovic, K.; Topisirovic, L. Cloning and expression of a novel lactococcal aggregation factor from Lactococcus lactis subsp. lactis BGKP1. BMC Microbiol. 2011, 19, 265. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.M.; Yan, T.R. DNA sequence analysis of a cryptic plasmid pL2 from Lactococcus lactis subsp. lactis. Biotechnol. Lett. 2007, 29, 1519–1527. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.Q.; Duan, K.M.; Dunn, N.W. Cloning and sequence analysis of a plasmid replicon from Lactococcus lactis subsp. cremoris FG2. J. Gen. Appl. Microbiol. 1997, 43, 75–80. [Google Scholar] [CrossRef]
- Jahns, A.; Schafer, A.; Geis, A.; Teuber, M. Identification, cloning and sequencing of the replication region of Lactococcus lactis ssp. lactis biovar. diacetylactis Bu2 citrate plasmid pSL2. FEMS Microbiol. Lett. 1991, 64, 253–258. [Google Scholar] [CrossRef]
- Boucher, I.; Emond, E.; Parrot, M.; Moineau, S. DNA sequence analysis of three Lactococcus lactis plasmids encoding phage resistance mechanisms. J. Dairy Sci. 2001, 84, 1610–1620. [Google Scholar] [CrossRef]
- Von Wright, A.; Raty, K. The nucleotide sequence for the replication region of pVS40, a lactococcal food grade cloning vector. Lett. Appl. Microbiol. 1993, 17, 25–28. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wang, Y.; Huo, G. Complete genome sequence of Lactococcus lactis subsp. lactis 4.0325. Genome Announc. 2013, 1, e00962-13. [Google Scholar]
- Seegers, J.F.; van Sinderen, D.; Fitzgerald, G.F. Molecular characterization of the lactococcal plasmid pCIS3: Natural stacking of specificity subunits of a type I restriction/modification system in a single lactococcal strain. Microbiology 2000, 146, 435–443. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, Y.; Hu, T.; Qu, X.; Zhang, L.; Ding, Z.; Dong, A. Plasmids from Food Lactic Acid Bacteria: Diversity, Similarity, and New Developments. Int. J. Mol. Sci. 2015, 16, 13172-13202. https://doi.org/10.3390/ijms160613172
Cui Y, Hu T, Qu X, Zhang L, Ding Z, Dong A. Plasmids from Food Lactic Acid Bacteria: Diversity, Similarity, and New Developments. International Journal of Molecular Sciences. 2015; 16(6):13172-13202. https://doi.org/10.3390/ijms160613172
Chicago/Turabian StyleCui, Yanhua, Tong Hu, Xiaojun Qu, Lanwei Zhang, Zhongqing Ding, and Aijun Dong. 2015. "Plasmids from Food Lactic Acid Bacteria: Diversity, Similarity, and New Developments" International Journal of Molecular Sciences 16, no. 6: 13172-13202. https://doi.org/10.3390/ijms160613172
APA StyleCui, Y., Hu, T., Qu, X., Zhang, L., Ding, Z., & Dong, A. (2015). Plasmids from Food Lactic Acid Bacteria: Diversity, Similarity, and New Developments. International Journal of Molecular Sciences, 16(6), 13172-13202. https://doi.org/10.3390/ijms160613172