
Lead Poisoning Disturbs Oligodendrocytes Differentiation Involved in Decreased Expression of NCX3 Inducing Intracellular Calcium Overload


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
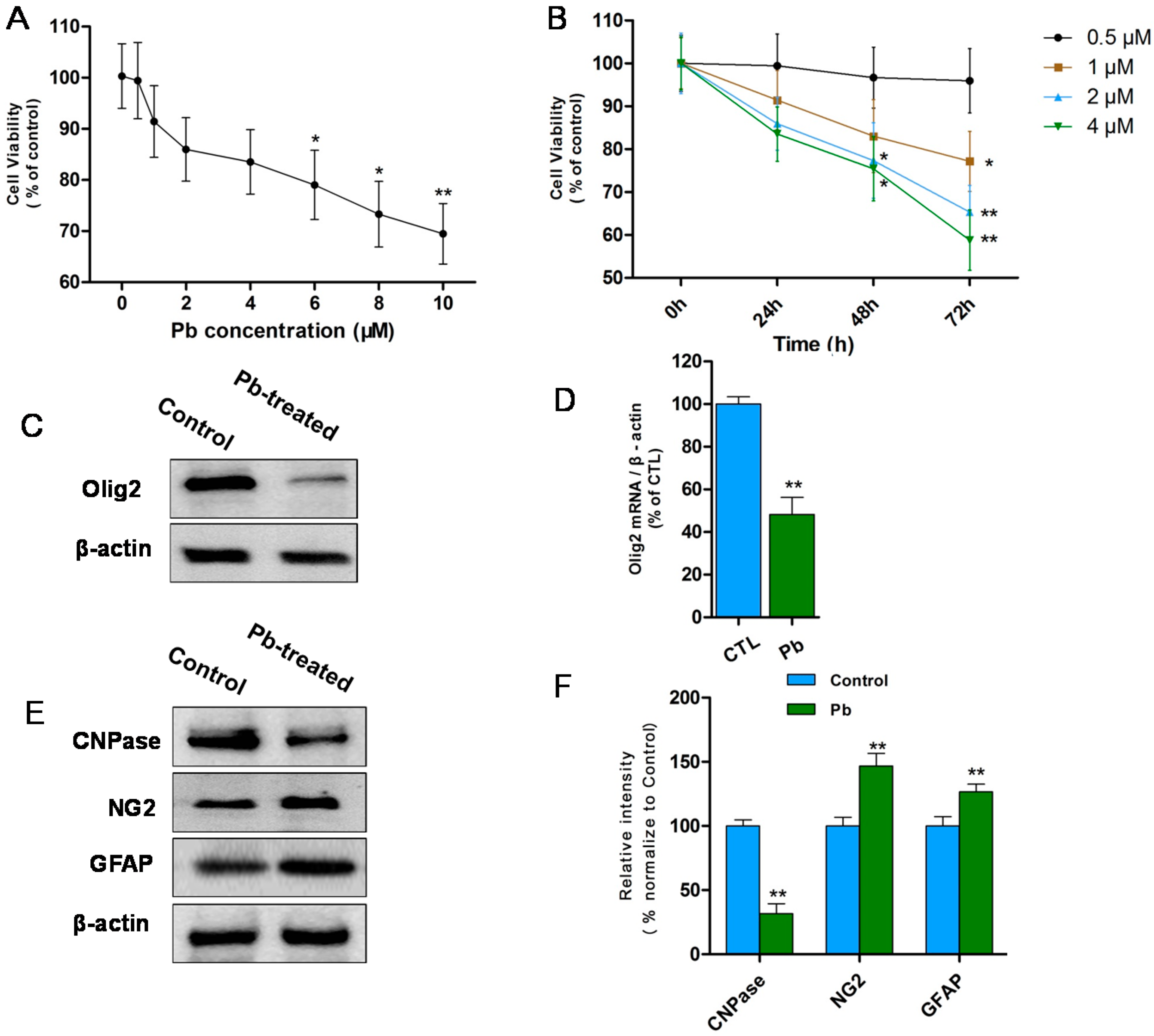
2.1. Discrepancies in Differentiation of OLs Precursor Cells (OPCs) at Low Concentrations of Pb in Vitro

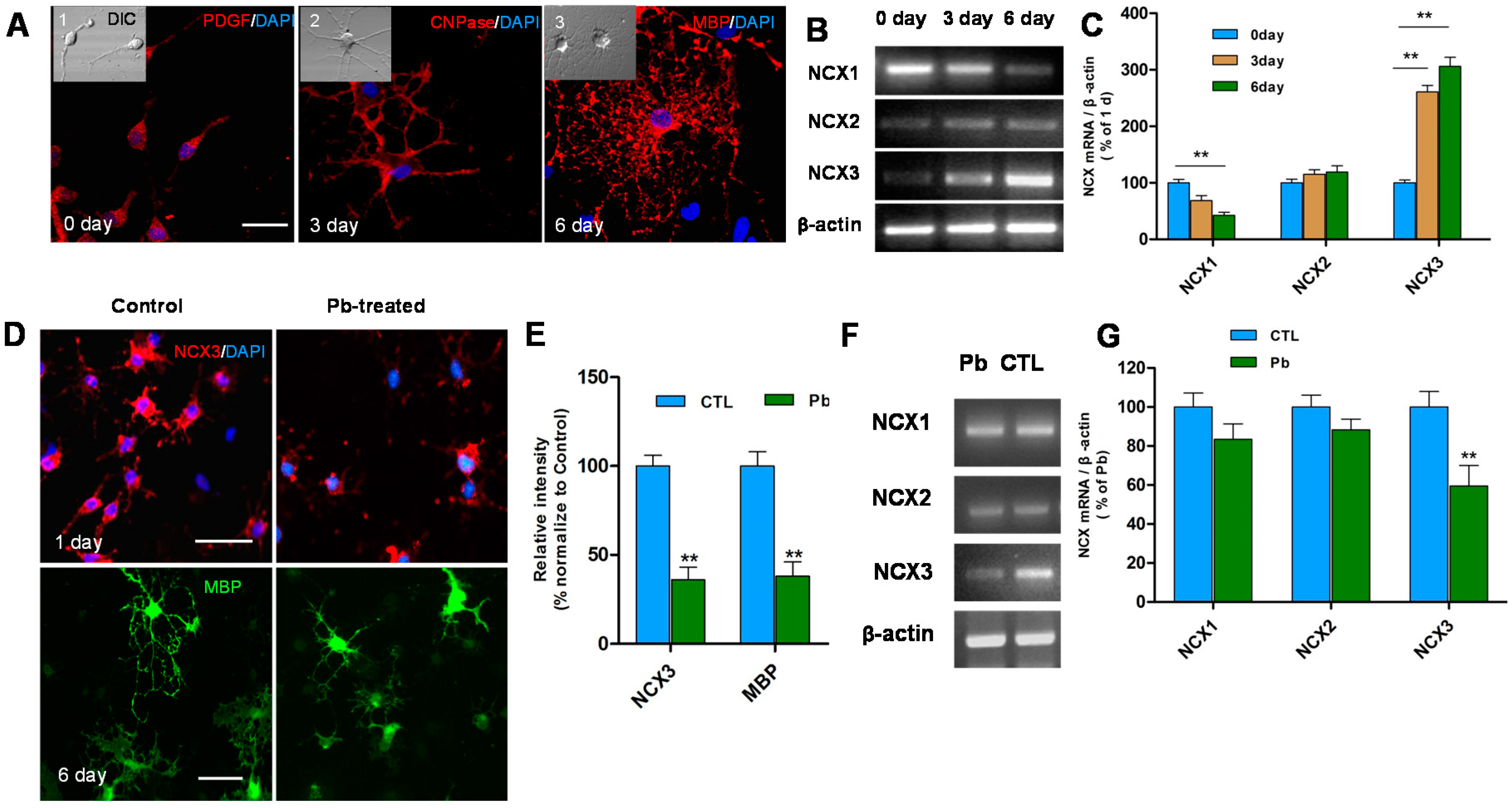
2.2. Pb Exposure Decreases the Expression of NCX3 and MBP in OLs Differentiation Model


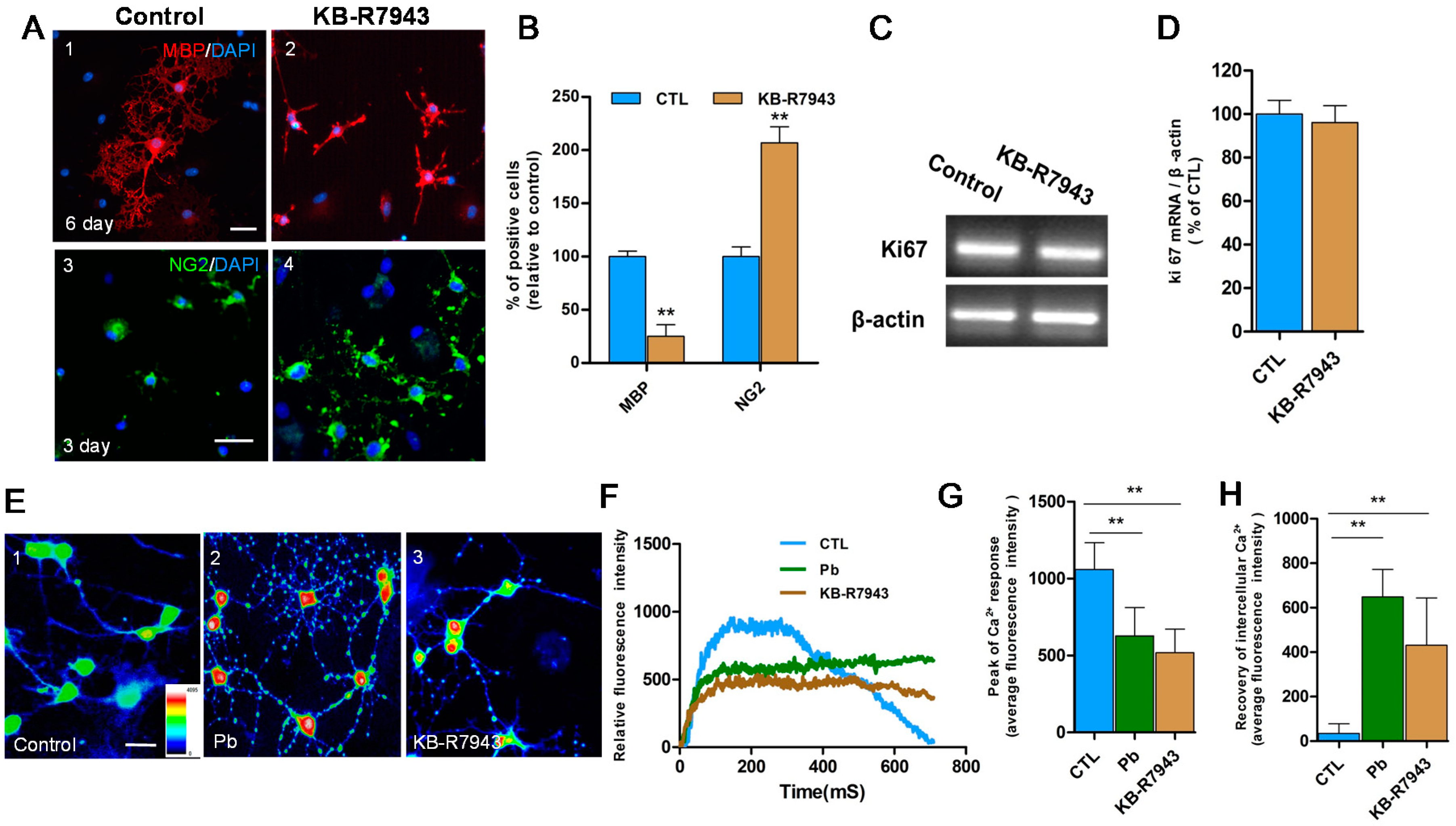
2.3. Pb Induced Inhibition of NCX3 and Increased Intracellular Calcium Interfering with OPCs Differentiation
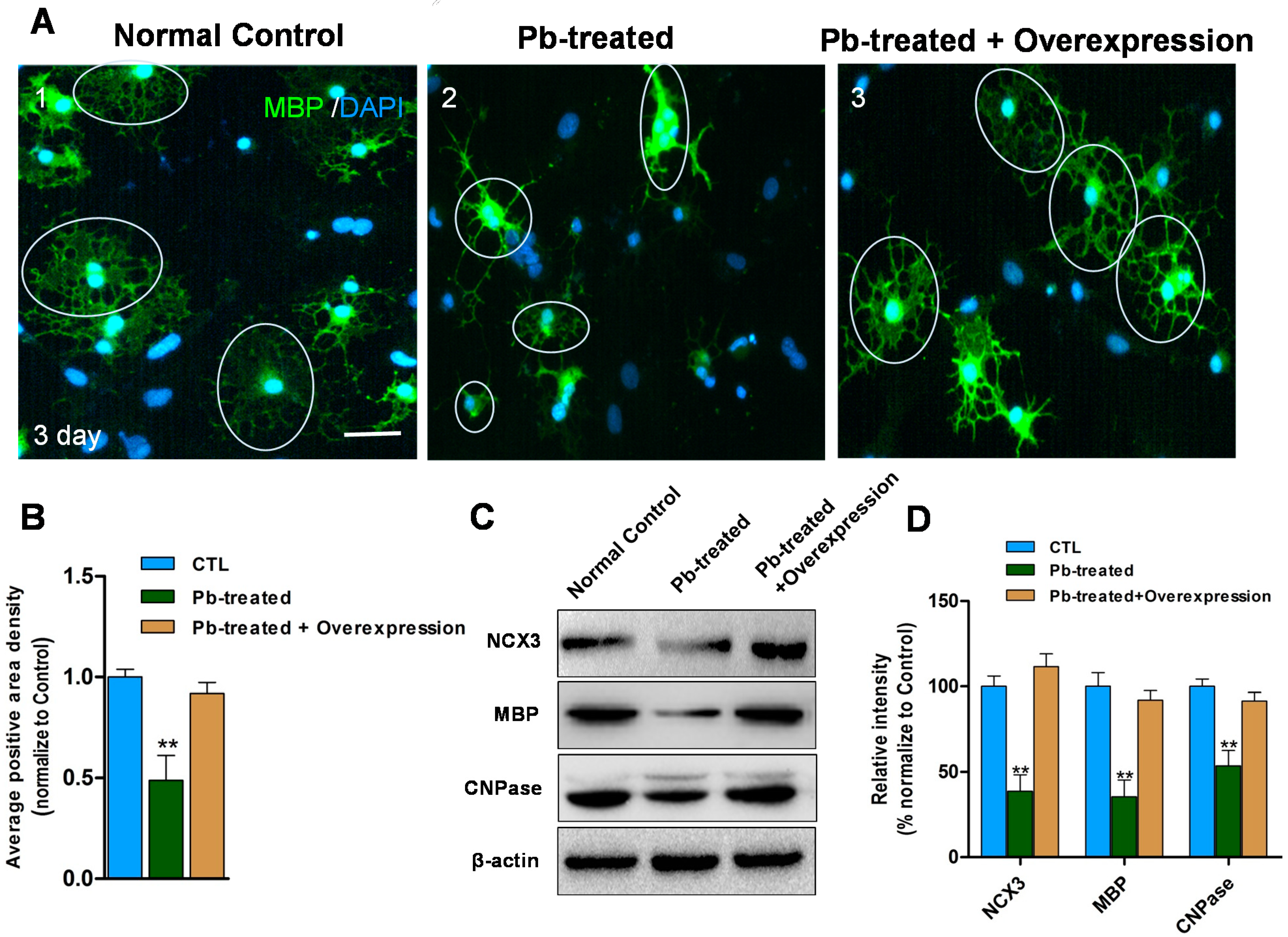
2.4. Overexpression of NCX3 Reverse Pb Induced Disturbances of OLs Differentiation

3. Discussion
4. Experimental Section
4.1. OPCs Culture and Differentiation
4.2. MTT Assay
4.3. Confocal Ca2+ Imaging
4.4. Immunohistochemistry and Immunofluorescent Staining
4.5. RT-PCR
4.6. Western Blot Analysis
4.7. OPCs Transfection
4.8. Quantitive Image Analysis
4.9. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Potash, E.; Walsh, J.; Brew, J.; Loewi, A.; Majumdar, S.; Reece, A.; Rozier, E.; Jorgensen, E.; Mansour, R.; Ghani, R. Predictive modeling for public health: Preventing childhood lead poisoning. In Proceedings of the 21th ACM SIGKDD International Conference on Knowledge Discovery and Data Mining, Sydney, NSW, Australia, 10–13 August 2015; pp. 2039–2047.
- Benoff, S.; Jacob, A.; Hurley, I.R. Male infertility and environmental exposure to lead and cadmium. Hum. Reprod. Update 2000, 6, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Burdette, L.J.; Goldstein, R. Long-term behavioral and electrophysiological changes associated with lead exposure at different stages of brain development in the rat. Dev. Brain Res. 1986, 29, 101–110. [Google Scholar] [CrossRef]
- Needleman, H. Lead poisoning. Annu. Rev. Med. 2004, 55, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, A.M.; Baniasadi, S.; Jorjani, M.; Rahimi, F.; Bakhshayesh, M. Investigation of acute lead poisoning on apoptosis in rat hippocampus in vivo. Neurosci. Lett. 2002, 329, 45–48. [Google Scholar] [CrossRef]
- Driscoll, J.W.; Stegner, S.E. Behavioral effects of chronic lead ingestion on laboratory rats. Pharmacol. Biochem. Behav. 1976, 4, 411–417. [Google Scholar] [CrossRef]
- Bleecker, M.L.; Ford, D.P.; Vaughan, C.G.; Walsh, K.S.; Lindgren, K.N. The association of lead exposure and motor performance mediated by cerebral white matter change. Neurotoxicology 2007, 28, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Lidsky, T.I.; Schneider, J.S. Lead neurotoxicity in children: Basic mechanisms and clinical correlates. Brain 2003, 126, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Tiffany-Castiglioni, E. Cell culture models for lead toxicity in neuronal and glial cells. Neurotoxicology 1993, 14, 513–536. [Google Scholar] [PubMed]
- Banik, N.L.; Shields, D.C. A putative role for calpain in demyelination associated with optic neuritis. Histol. Histopathol. 1999, 14, 649–656. [Google Scholar] [PubMed]
- Krishnan, C.; Kaplin, A.I.; Pardo, C.A.; Kerr, D.A.; Keswani, S.C. Demyelinating disorders: Update on transverse myelitis. Curr. Neurol. Neurosci. Rep. 2006, 6, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, M. Effectors of demyelination and remyelination in the CNS: Implications for multiple sclerosis. Brain Pathol. 2007, 17, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Soliven, B. Calcium signalling in cells of oligodendroglial lineage. Microsc. Res. Tech. 2001, 52, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Gurer, H.; Ercal, N. Can antioxidants be beneficial in the treatment of lead poisoning? Free Radic. Biol. Med. 2000, 29, 927–945. [Google Scholar] [CrossRef]
- Kip, S.N.; Strehler, E.E. Rapid downregulation of NCX and PMCA in hippocampal neurons following H2O2 oxidative stress. Ann. N. Y. Acad. Sci. 2007, 1099, 436–439. [Google Scholar] [CrossRef] [PubMed]
- Quednau, B.D.; Nicoll, D.A.; Philipson, K.D. Tissue specificity and alternative splicing of the Na+/Ca2+ exchanger isoforms NCX1, NCX2, and NCX3 in rat. Am. J. Physiol. 1997, 272, C1250–C1261. [Google Scholar] [PubMed]
- Nicoll, D.A.; Longoni, S.; Philipson, K.D. Molecular cloning and functional expression of the cardiac sarcolemmal Na+–Ca2+ exchanger. Science 1990, 250, 562–565. [Google Scholar] [CrossRef] [PubMed]
- Thurneysen, T.; Nicoll, D.A.; Philipson, K.D.; Porzig, H. Sodium/calcium exchanger subtypes NCX1, NCX2 and NCX3 show cell-specific expression in rat hippocampus cultures. Mol. Brain Res. 2002, 107, 145–156. [Google Scholar] [CrossRef]
- Molinaro, P.; Viggiano, D.; Nistico, R.; Sirabella, R.; Secondo, A.; Boscia, F.; Pannaccione, A.; Scorziello, A.; Mehdawy, B.; Sokolow, S.; et al. Na+–Ca2+ exchanger (NCX3) knock-out mice display an impairment in hippocampal long-term potentiation and spatial learning and memory. J. Neurosci. 2011, 31, 7312–7321. [Google Scholar] [CrossRef] [PubMed]
- Boscia, F.; D’Avanzo, C.; Pannaccione, A.; Secondo, A.; Casamassa, A.; Formisano, L.; Guida, N.; Sokolow, S.; Herchuelz, A.; Annunziato, L. Silencing or knocking out the Na+/Ca2+ exchanger-3 (NCX3) impairs oligodendrocyte differentiation. Cell Death Differ. 2012, 19, 562–572. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; McKinnon, R.D.; Poretz, R.D. Lead exposure delays the differentiation of oligodendroglial progenitors in vitro. Toxicol. Appl. Pharmacol. 2001, 174, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S. Heavy metal pollution in China: Origin, pattern and control. Environ. Sci. Pollut. Res. Int. 2003, 10, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Leggett, R.W. An age-specific kinetic model of lead metabolism in humans. Environ. Health Perspect. 1993, 101, 598–616. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Hu, Y. Lead (Pb) isotopic fingerprinting and its applications in lead pollution studies in China: A review. Environ. Pollut. 2010, 158, 1134–1146. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.-M.; Rosen, J.F.; Guo, D.; Wu, S.-M. Childhood lead poisoning in China. Sci. Total Environ. 1996, 181, 101–109. [Google Scholar] [CrossRef]
- Meyer, P.A.; Brown, M.J.; Falk, H. Global approach to reducing lead exposure and poisoning. Mutat. Res. 2008, 659, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Coria, F.; Berciano, M.T.; Berciano, J.; Lafarga, M. Axon membrane remodeling in the lead-induced demyelinating neuropathy of the rat. Brain Res. 1984, 291, 369–372. [Google Scholar] [CrossRef]
- Martinez, Y.; N’Gouemo, P. Blockade of the sodium calcium exchanger exhibits anticonvulsant activity in a pilocarpine model of acute seizures in rats. Brain Res. 2010, 1366, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Bano, D.; Young, K.W.; Guerin, C.J.; Lefeuvre, R.; Rothwell, N.J.; Naldini, L.; Rizzuto, R.; Carafoli, E.; Nicotera, P. Cleavage of the plasma membrane Na+/Ca2+ exchanger in excitotoxicity. Cell 2005, 120, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Orrenius, S.; Zhivotovsky, B.; Nicotera, P. Regulation of cell death: The calcium-apoptosis link. Nat. Rev. Mol. Cell Biol. 2003, 4, 552–565. [Google Scholar] [CrossRef] [PubMed]
- Verkhratsky, A.; Rodriguez, J.J.; Parpura, V. Calcium signalling in astroglia. Mol. Cell. Endocrinol. 2012, 353, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Gokhan, S.; Marin-Husstege, M.; Yung, S.Y.; Fontanez, D.; Casaccia-Bonnefil, P.; Mehler, M.F. Combinatorial profiles of oligodendrocyte-selective classes of transcriptional regulators differentially modulate myelin basic protein gene expression. J. Neurosci. 2005, 25, 8311–8321. [Google Scholar] [CrossRef] [PubMed]
- Marshall, C.A.; Novitch, B.G.; Goldman, J.E. Olig2 directs astrocyte and oligodendrocyte formation in postnatal subventricular zone cells. J. Neurosci. 2005, 25, 7289–7298. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.R.; Sun, T.; Zhu, Z.; Ma, N.; Garcia, M.; Stiles, C.D.; Rowitch, D.H. Common developmental requirement for Olig function indicates a motor neuron/oligodendrocyte connection. Cell 2002, 109, 75–86. [Google Scholar] [CrossRef]
- Ligon, K.L.; Fancy, S.P.; Franklin, R.J.; Rowitch, D.H. Olig gene function in CNS development and disease. Glia 2006, 54, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ligon, K.L.; Kesari, S.; Kitada, M.; Sun, T.; Arnett, H.A.; Alberta, J.A.; Anderson, D.J.; Stiles, C.D.; Rowitch, D.H. Development of NG2 neural progenitor cells requires Olig gene function. Proc. Natl. Acad. Sci. USA 2006, 103, 7853–7858. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.S.; Tatsumi, K.; Okuda, H.; Shiosaka, S.; Wanaka, A. Olig2-expressing progenitor cells preferentially differentiate into oligodendrocytes in cuprizone-induced demyelinated lesions. Neurochem. Int. 2009, 54, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Wang, S.; Anderson, D.J. Identification of a novel family of oligodendrocyte lineage-specific basic helix-loop-helix transcription factors. Neuron 2000, 25, 331–343. [Google Scholar] [CrossRef]
- Niu, J.; Wang, L.; Liu, S.; Li, C.; Kong, J.; Shen, H.Y.; Xiao, L. An efficient and economical culture approach for the enrichment of purified oligodendrocyte progenitor cells. J. Neurosci. Methods 2012, 209, 241–249. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, T.; Wu, X.; Cai, Q.; Wang, Y.; Xiao, L.; Tian, Y.; Li, H. Lead Poisoning Disturbs Oligodendrocytes Differentiation Involved in Decreased Expression of NCX3 Inducing Intracellular Calcium Overload. Int. J. Mol. Sci. 2015, 16, 19096-19110. https://doi.org/10.3390/ijms160819096
Ma T, Wu X, Cai Q, Wang Y, Xiao L, Tian Y, Li H. Lead Poisoning Disturbs Oligodendrocytes Differentiation Involved in Decreased Expression of NCX3 Inducing Intracellular Calcium Overload. International Journal of Molecular Sciences. 2015; 16(8):19096-19110. https://doi.org/10.3390/ijms160819096
Chicago/Turabian StyleMa, Teng, Xiyan Wu, Qiyan Cai, Yun Wang, Lan Xiao, Yanping Tian, and Hongli Li. 2015. "Lead Poisoning Disturbs Oligodendrocytes Differentiation Involved in Decreased Expression of NCX3 Inducing Intracellular Calcium Overload" International Journal of Molecular Sciences 16, no. 8: 19096-19110. https://doi.org/10.3390/ijms160819096
APA StyleMa, T., Wu, X., Cai, Q., Wang, Y., Xiao, L., Tian, Y., & Li, H. (2015). Lead Poisoning Disturbs Oligodendrocytes Differentiation Involved in Decreased Expression of NCX3 Inducing Intracellular Calcium Overload. International Journal of Molecular Sciences, 16(8), 19096-19110. https://doi.org/10.3390/ijms160819096