
Maternal PUFA ω-3 Supplementation Prevents Neonatal Lung Injuries Induced by Hyperoxia in Newborn Rats

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
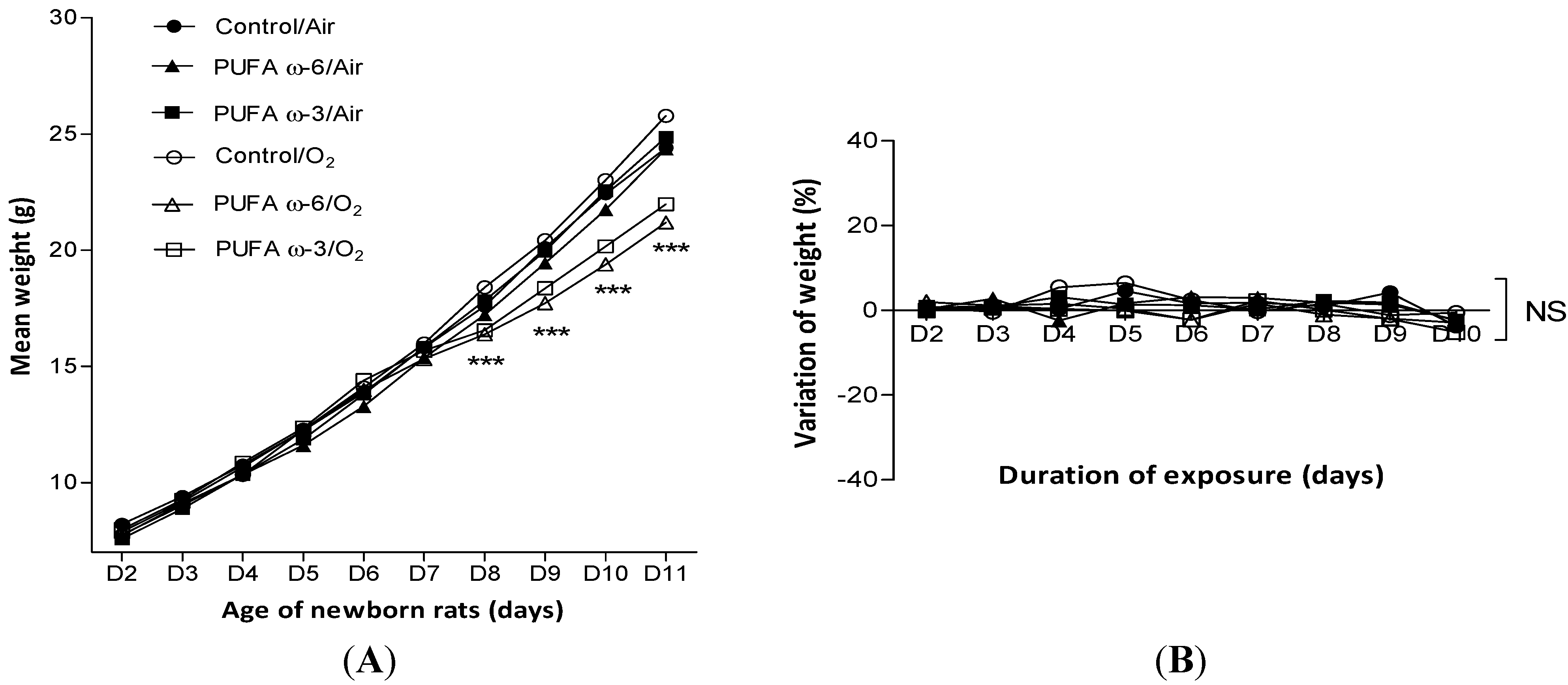
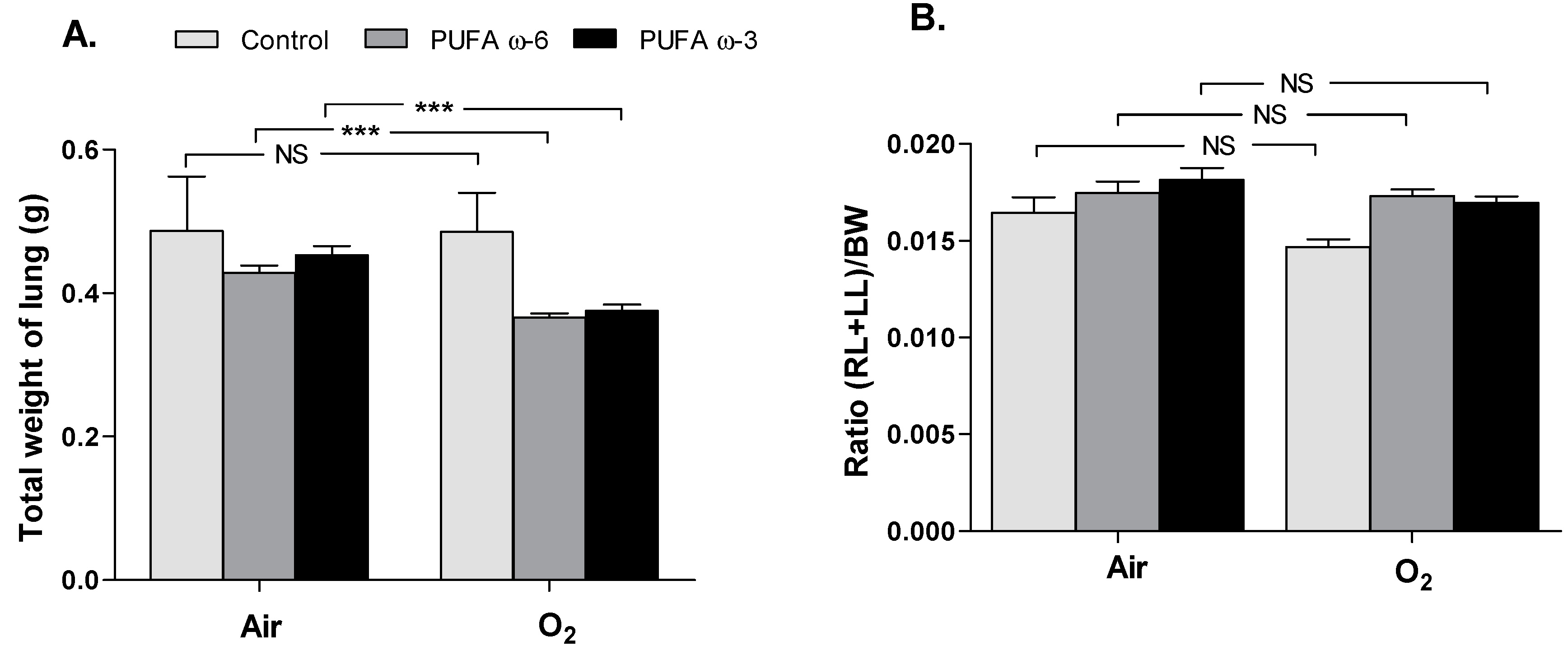
2.1. Effects of Hyperoxia on Survival and Growth of Newborns Rats and of Nursing Adult Rats

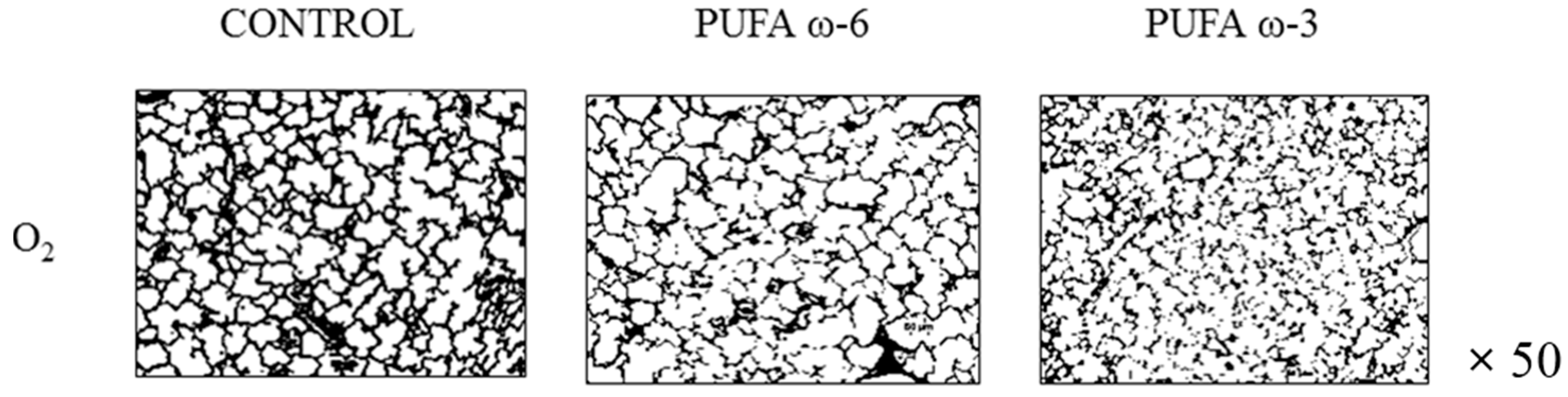
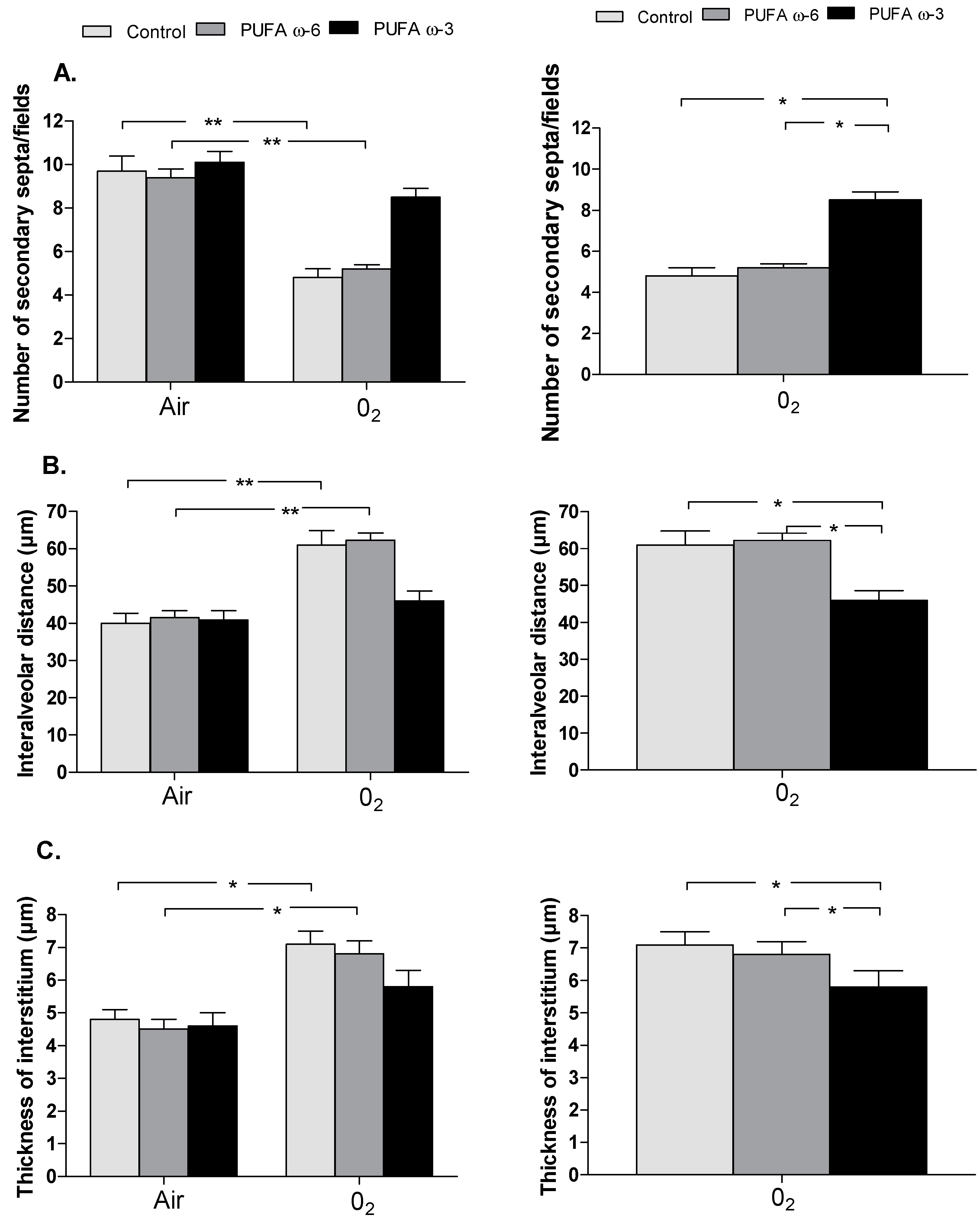
2.2. Effects of PUFA ω-3 on Lung Airway Development



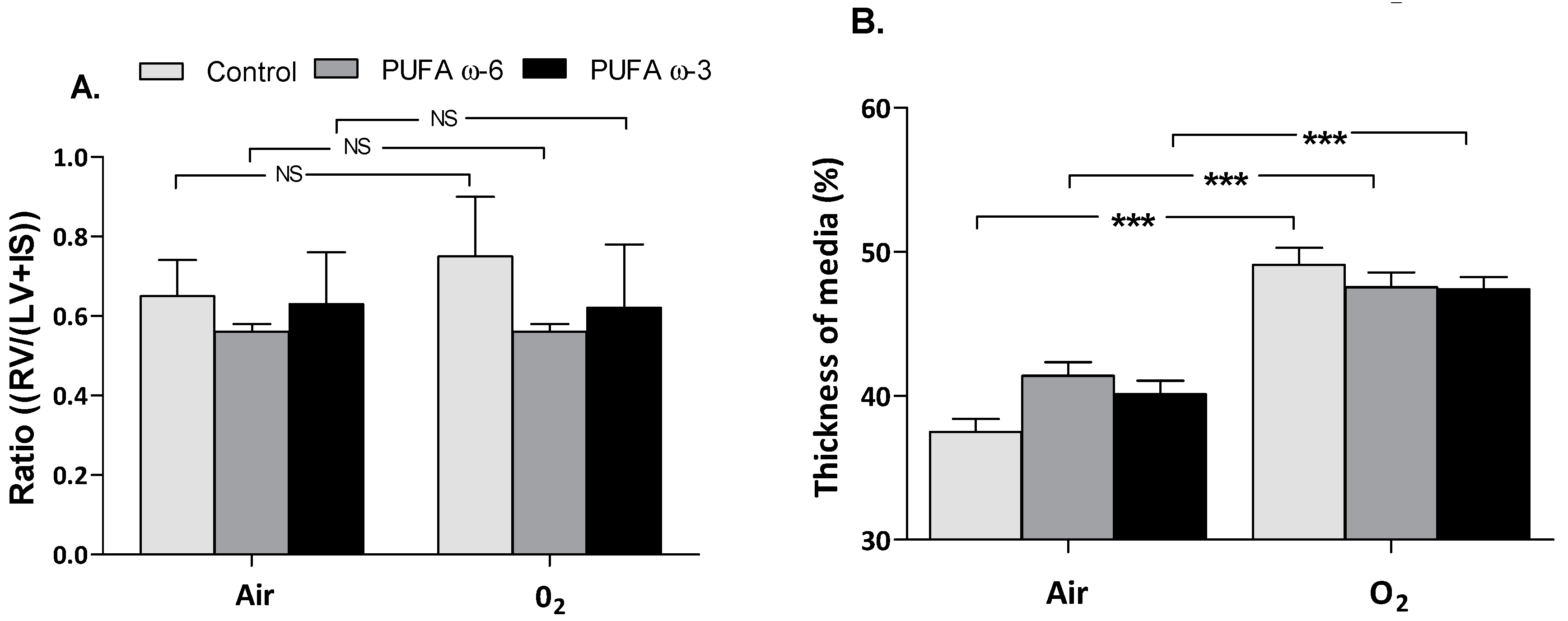
2.3. Effects of PUFA ω-3 on Cardiovascular Development


3. Discussion
4. Experimental Section
4.1. Animals
4.2. Experimental Design
4.3. Tissue Preparation and Histomorphometric Analysis
4.4. Main Parameters of Study
4.5. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hilgendorff, A.; Reiss, I.; Ehrhardt, H.; Eickelberg, O.; Alvira, C.M. Chronic lung disease in the preterm infant. Am. J. Respir. Cell Mol. Biol. 2014, 50, 233–245. [Google Scholar] [PubMed]
- Speer, C.P. Inflammation and bronchopulmonary dysplasia: A continuing story. Semin. Fetal Neonatal Med. 2006, 11, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Bose, C.L.; Damman, C.E.; Laughon, M.M. Bronchopulmonary dysplasia and inflammatory biomarkers in the premature neonate. Arch. Dis. Child. Fetal Neonatal Ed. 2008, 93, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Schneibel, K.R.; Fitzpatrick, A.M.; Ping, X.D.; Brown, L.-A.S.; Gauthier, T.W. Inflammatory mediator patterns in tracheal aspirate and their association with bronchopulmonary dysplasia in very low birth weight neonates. J. Perinatol. 2013, 33, 383–387. [Google Scholar] [CrossRef] [PubMed]
- Vom Hove, M.; Prenzel, F.; Uhlig, H.H.; Robel-Tillig, E. Pulmonary outcome in former preterm, very low birth weight children with bronchopulmonary dysplasia: A case-control follow-up at school age. J. Pediatr. 2014, 164, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, J.A.; Bell, S.J.; van Ausdal, W. Omega-3 fatty acid supplementation during pregnancy. Rev. Obstet. Gynecol. 2008, 1, 162–169. [Google Scholar] [PubMed]
- Schwartz, J.; Dube, K.; Alexy, U.; Kalhoff, H.; Kersting, M. PUFA and LC-PUFA intake during the first year of life: Can dietary practice achieve a guideline diet? Eur. J. Clin. Nutr. 2010, 64, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Carlson, S.E.; Werkman, S.H.; Tolley, E.A. Effect of long-chain n-3 fatty acids supplementation on visual acuity and growth of preterm infants with and without bronchopulmonary dysplasia. Am. J. Clin. Nutr. 1996, 63, 687–697. [Google Scholar] [PubMed]
- Hanebutt, F.L.; Demmelmair, H.; Schiessl, B.; Larqué, E.; Koletzko, B. Long-chain polyunsaturated fatty acid (LC-PUFA) transfer across the placenta. Clin. Nutr. ESPEN 2008, 27, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Agget, P.J.; Agostoni, C.; Axelsson, I.; de Curtis, M.; Goulet, O.; Hernell, O.; Koletzko, B.; Lafeber, H.N.; Michaelsen, K.F.; Puntis, J.W.L.; et al. Feeding preterm infants after hospital discharge: A commentary by the ESPGHAN committee on nutrition. J. Pediatr. Gastroenterol. Nutr. 2006, 42, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Lavoie, P.M.; Lacase-Masmonteil, T.; Rhainds, M.; Marc, I. Omega-3 long-chain polyunsaturated fatty acids for extremely preterm infants: A systematic review. Pediatrics 2014, 134, 120–134. [Google Scholar] [CrossRef] [PubMed]
- Larsen, B.M.K.; Goonewardene, L.A.; Joffre, A.R.; van Aerde, J.E.; Field, C.J.; Olstad, D.L.; Clandinin, M.T. Pre-treatment with an intravenous lipid emulsion containing fish oil (eicosapentaenoic and docosahexaenoic acid) decreases inflammatory markers after open-heart surgery in infants: A randomized, controlled trial. Clin. Nutr. 2012, 31, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Larsen, B.M.K.; Field, C.J.; Leong, A.Y.; Goonewardene, L.A.; van Aerde, J.E.; Joffre, A.R.; Olstad, D.L.; Clandinin, M.T. Pretreatment with an intravenous lipid emulsion increases plasma eicosapentaenoic acid and downregulates leukotriene B4, procalcitonin, and lymphocte concentrations after open heart surgery in infants. J. Parenter. Enter. Nutr. 2015, 39, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Rieger-Fackeldey, E.; Hentschel, R. Bronchopulmonary dysplasia and early prophylactic inhaled nitric oxide in preterm infants: current concepts and future research strategies in animal models. J. Perinat. Med. 2008, 36, 442–447. [Google Scholar] [CrossRef] [PubMed]
- Berger, J.; Bhandari, V. Animal models of bronchopulmonary dysplasia. The term mouse models. Am. J. Physiol. Lung Cell Mol. Physiol. 2014, 307, 936–947. [Google Scholar] [CrossRef] [PubMed]
- De Visser, Y.P.; Walther, F.J.; el Laghmani, H.; van der Laarse, A.; Wagenaar, G.T.M. Apelin attenuates hyperoxic lung and heart injury in neonatal rats. Am. J. Respir. Crit. Care Med. 2010, 15, 1239–1250. [Google Scholar] [CrossRef] [PubMed]
- De Visser, Y.P.; Walther, F.J.; el Laghmani, H.; Steendijk, P.; Middeldorp, M.; van der Laarse, A.; Wagenaar, G.T. Phosphodiesterase 4 inhibition attenuates persistent heart and lung injury by neonatal hyperoxia in rats. Am. J. Physiol. Lung Cell Mol. Physiol. 2012, 1, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Houeijeh, A.; Aubry, E.; Coridon, H.; Montaigne, K.; Sfeir, R.; Deruelle, P.; Storme, L. Effects of n-3 polyunsaturated fatty acids in the fetal pulmonary circulation. Crit. Care Med. 2011, 39, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.U.; Kathirvel, K.; Choudhury, S.; Garg, S.K.; Mishra, S.K. Eicosapentaenoic acid-induced endothelium-dependent and -independent relaxation of sheep pulmonary artery. Eur. J. Pharmacol. 2010, 636, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Omura, M.; Kobayashi, S.; Mizukami, Y.; Mogami, K.; Todoroki-Ikeda, N.; Miyake, T.; Matsuzaki, M. Eicosapentaenoic acid (EPA) induces Ca2+-independent activation and translocation of endothelial nitric oxide synthase and endothelium-dependent vasorelaxation. FEBS Lett. 2001, 487, 361–366. [Google Scholar] [CrossRef]
- Alphonse, R.S.; Vadivel, A.; Coltan, L.; Eaton, F.; Barr, A.J.; Dyck, J.R.B.; Thebaud, B. Activation of Akt protects alveoli from neonatal oxygen-induced lung injury. Am. J. Respir. Cell Mol. Biol. 2011, 44, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Porzionato, A.; Sfriso, M.M.; Mazzatenta, A.; Macchi, V.; de Caro, R.; di Giulio, C. Effects of hyperoxic exposure on signal transduction pathways in the lung. Respir. Physiol. Neurobiol. 2015, 209, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Morin, C.; Hiram, R.; Rousseau, E.; Blier, P.U.; Fortin, S. Docosapentaenoic acid monoacylglyceride reduces inflammation and vascular remodeling in experimental pulmonary hypertension. Am. J. Physiol. Heart Circ. Physiol. 2014, 307, 574–586. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.R.; Zaman, M.M.; Gilkey, C.; Salguero, M.V.; Hasturk, H.; Kantarci, A.; van Dyke, T.E.; Freedman, S.D. Resolvin D1 and lipoxin A4 improve alveolarization and normalize septal wall thickness in a neonatal murine model of hyperoxia-induced lung injury. PLoS ONE 2014, 9, e98773. [Google Scholar] [CrossRef] [PubMed]
- Béghin, L.; Storme, L.; Coopman, S.; Razka, T.; Gottrand, F. Parenteral nutrition with fish oil supplements is safe and seems to be effective in severe preterm neonates with respiratory distress syndrome. Acta Paediatr. 2015. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.J.; Markham, N.E.; Balasubramaniam, V.; Tang, J.R.; Maxey, A.; Kinsella, J.P.; Abman, S.H. Inhaled nitric oxide enhances distal lung growth after exposure to hyperoxia in neonatal rats. Pediatr. Res. 2005, 58, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.R.; Seedorf, G.J.; Muehlethaler, V.; Walker, D.L.; Markham, N.E.; Balasubramaniam, V.; Abman, S.H. Moderate postnatal hyperoxia accelerates lung growth and attenuates pulmonary hypertension in infant rats after exposure to intra-amniotic endotoxin. Am. J. Physiol. Lung Cell Mol. Physiol. 2010, 6, 735–748. [Google Scholar] [CrossRef] [PubMed]
- Deruelle, P.; Balasubramaniam, V.; Kunig, A.M.; Seedorf, G.J.; Markham, N.E.; Abman, S.H. BAY 41–2272, a direct activator of soluble guanylate cyclase, reduces right ventricular hypertrophy and prevents pulmonary vascular remodeling during chronic hypoxia in neonatal rats. Biol. Neonate 2006, 90, 135–144. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, D.; Nkembi, A.S.; Aubry, E.; Houeijeh, A.; Butruille, L.; Houfflin-Debarge, V.; Besson, R.; Deruelle, P.; Storme, L. Maternal PUFA ω-3 Supplementation Prevents Neonatal Lung Injuries Induced by Hyperoxia in Newborn Rats. Int. J. Mol. Sci. 2015, 16, 22081-22093. https://doi.org/10.3390/ijms160922081
Sharma D, Nkembi AS, Aubry E, Houeijeh A, Butruille L, Houfflin-Debarge V, Besson R, Deruelle P, Storme L. Maternal PUFA ω-3 Supplementation Prevents Neonatal Lung Injuries Induced by Hyperoxia in Newborn Rats. International Journal of Molecular Sciences. 2015; 16(9):22081-22093. https://doi.org/10.3390/ijms160922081
Chicago/Turabian StyleSharma, Dyuti, Armande Subayi Nkembi, Estelle Aubry, Ali Houeijeh, Laura Butruille, Véronique Houfflin-Debarge, Rémi Besson, Philippe Deruelle, and Laurent Storme. 2015. "Maternal PUFA ω-3 Supplementation Prevents Neonatal Lung Injuries Induced by Hyperoxia in Newborn Rats" International Journal of Molecular Sciences 16, no. 9: 22081-22093. https://doi.org/10.3390/ijms160922081
APA StyleSharma, D., Nkembi, A. S., Aubry, E., Houeijeh, A., Butruille, L., Houfflin-Debarge, V., Besson, R., Deruelle, P., & Storme, L. (2015). Maternal PUFA ω-3 Supplementation Prevents Neonatal Lung Injuries Induced by Hyperoxia in Newborn Rats. International Journal of Molecular Sciences, 16(9), 22081-22093. https://doi.org/10.3390/ijms160922081