
Transcriptome Analysis Identifies Key Candidate Genes Mediating Purple Ovary Coloration in Asiatic Hybrid Lilies

 ,
,
Abstract
:1. Introduction
2. Results
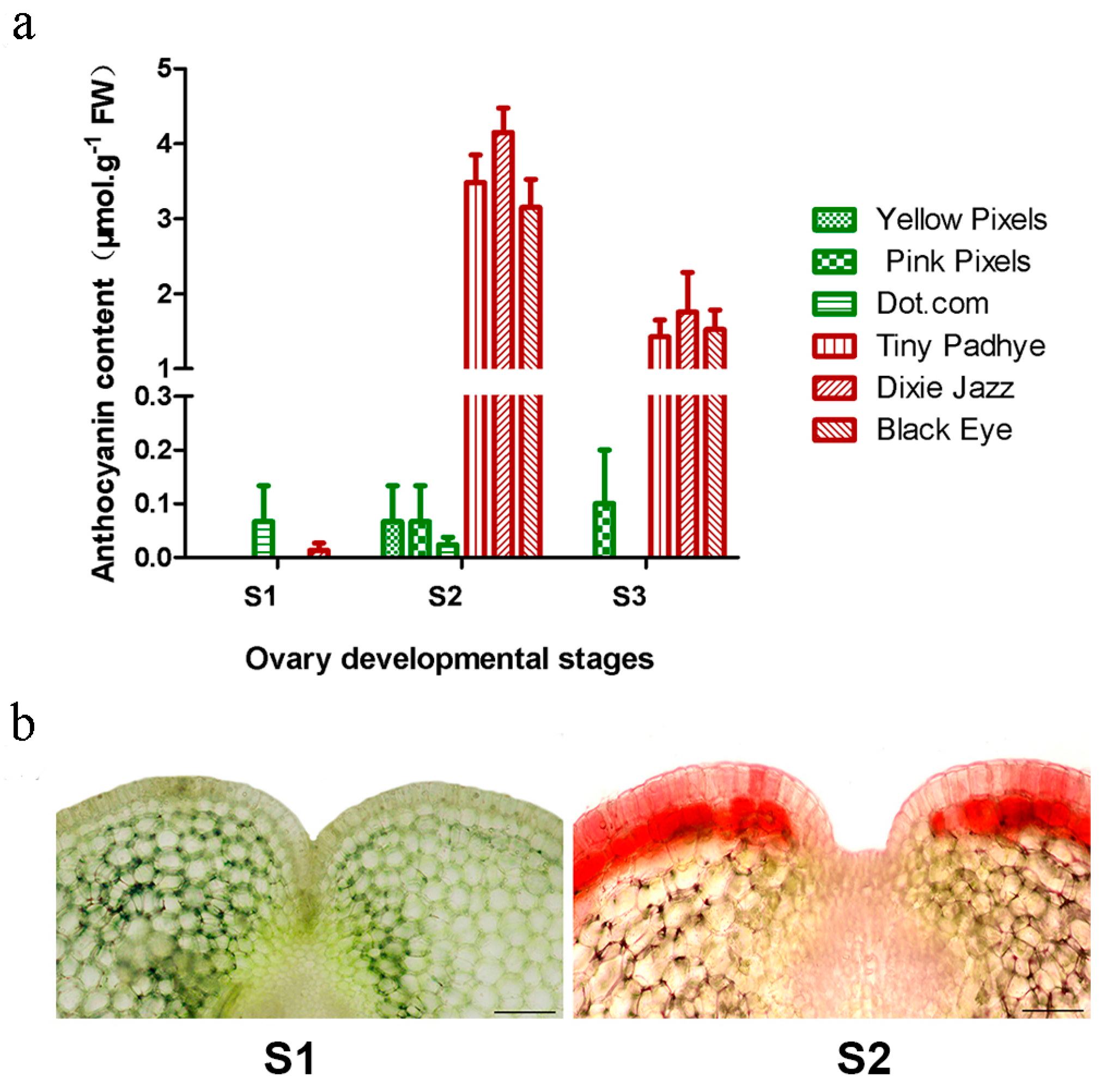
2.1. Measurement of Anthocyanin Content
2.2. Sequencing and Sequence Assembly
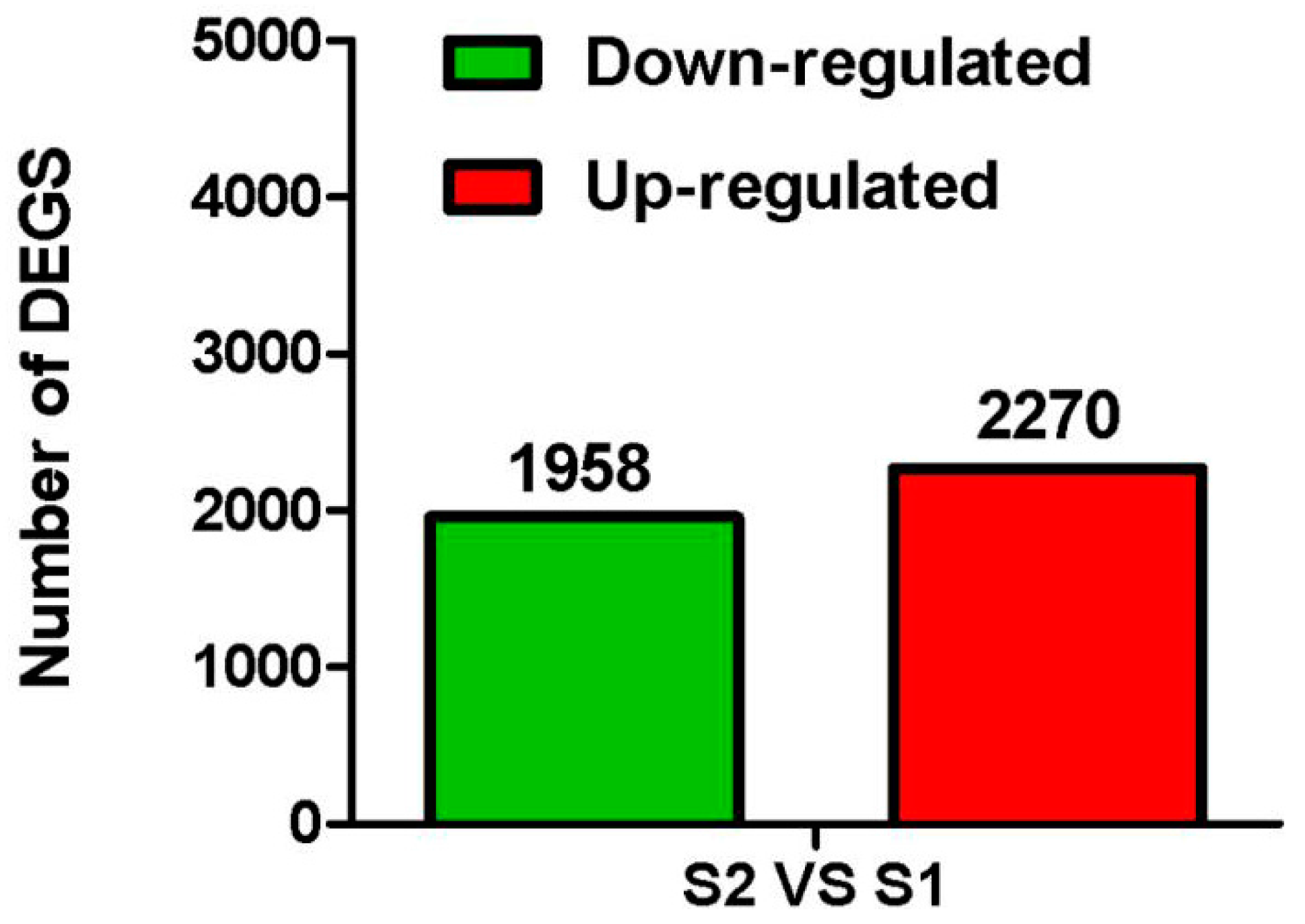
2.3. Identification of Differentially Expressed Genes
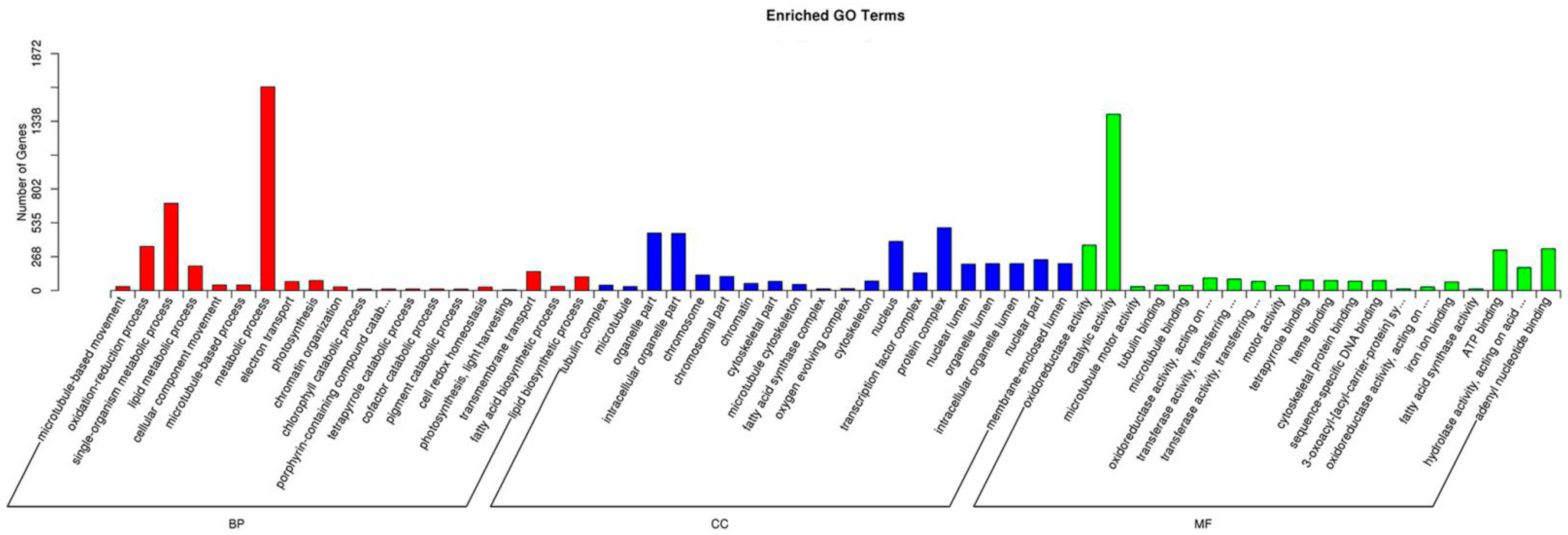
2.4. Functional Analysis of Differentially Expressed Genes
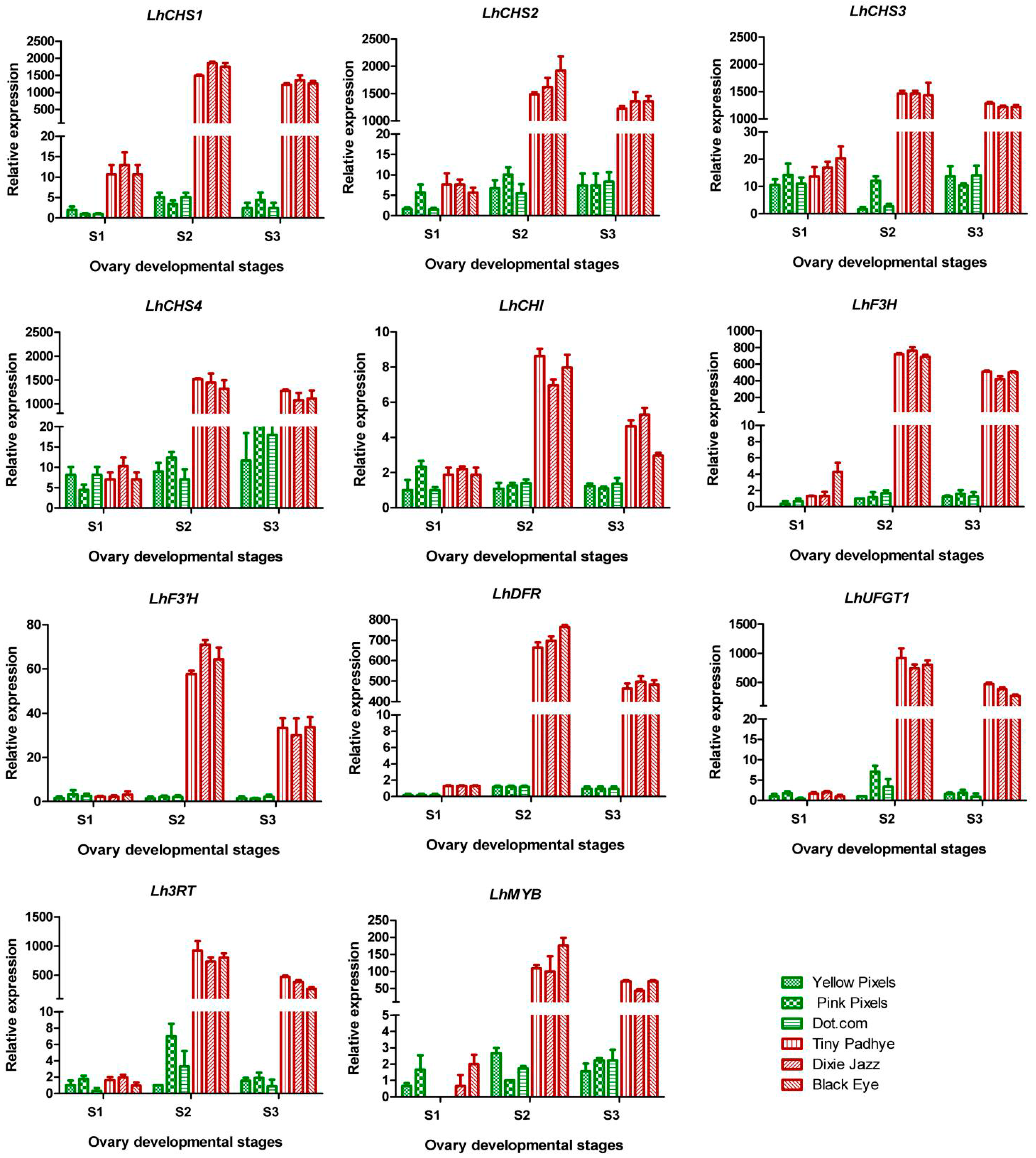
2.5. Expression Patterns of Genes Involved in Anthocyanin Biosynthesis Pathway
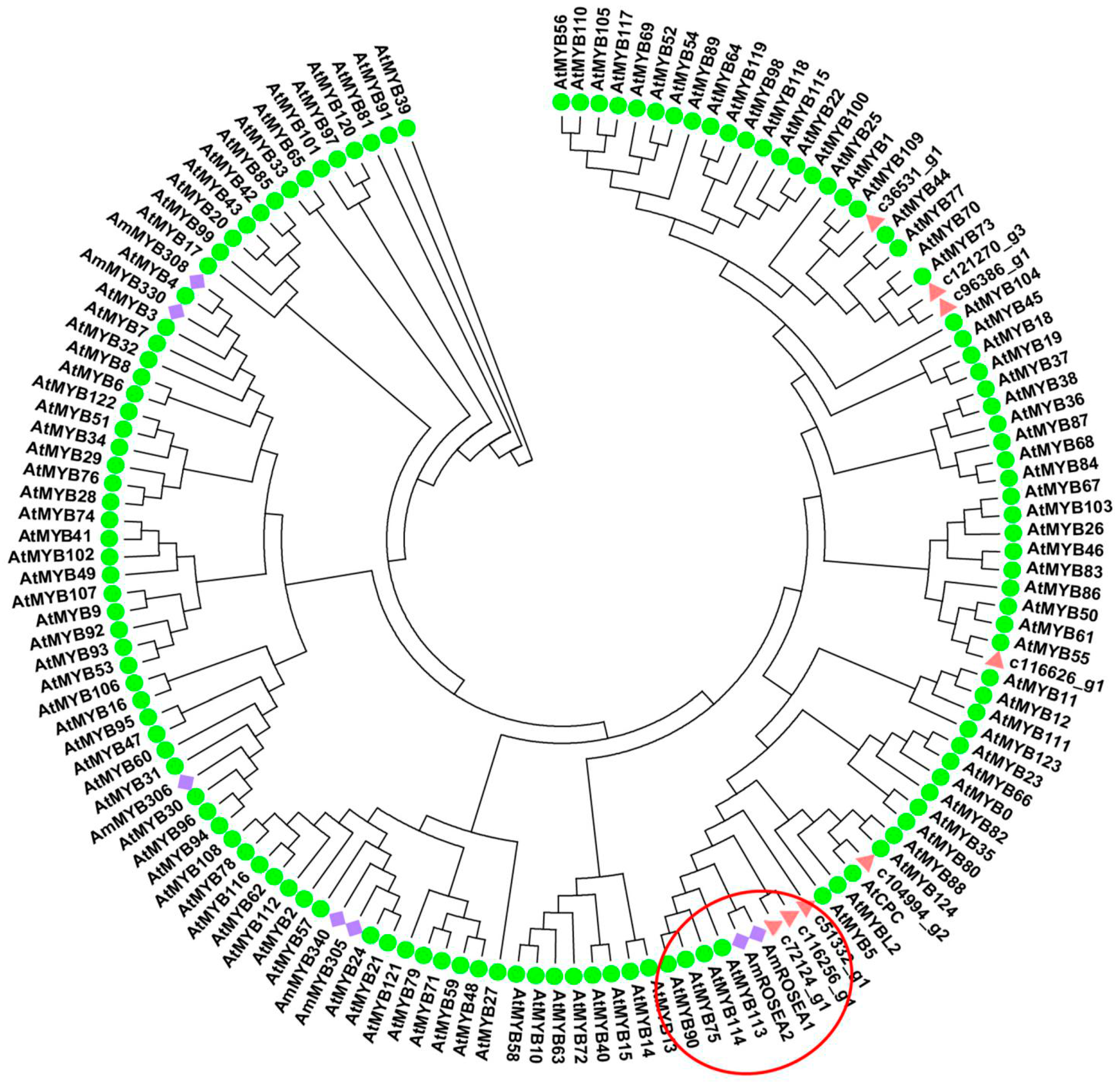
2.6. Phylogenetic Analysis of R2R3-MYB Transcription Factors
2.7. qPCR Validation of RNA-Seq Data
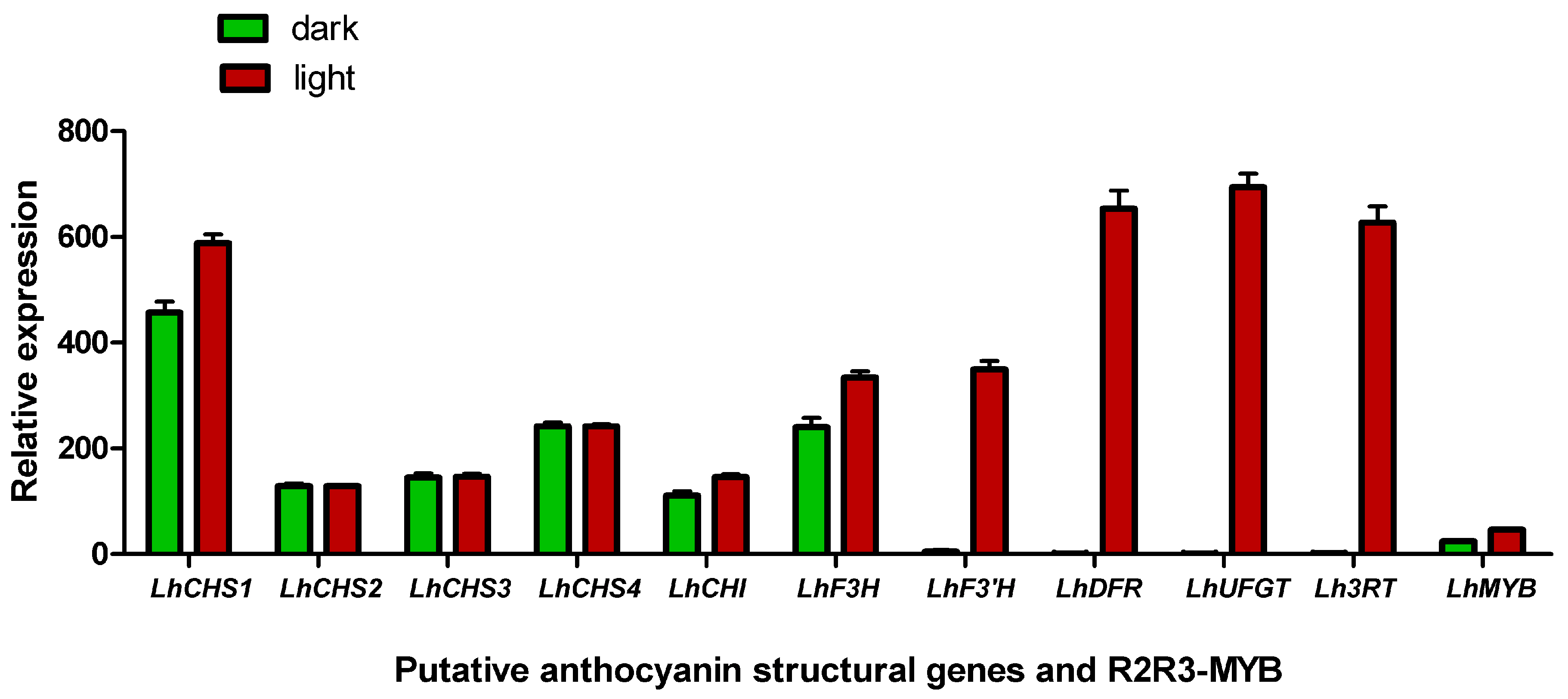
2.8. Effect of Shading Treatment on Pigment Accumulation and Gene Expression
2.9. Simple Sequence Repeat (SSR) Marker Identification and Characterization
3. Discussion
3.1. Co-Expression of Genes Involved in Anthocyanin Biosynthesis Determines Anthocyanin Accumulation in Purple Ovaries
3.2. LhMYB12-Lat Is Responsible for Purple Ovaries
3.3. Shading Treatment Decreased Pigment Accumulation
3.4. SSR Marker Identification and Characterization
4. Materials and methods
4.1. Plant Materials
4.2. Measurement of Anthocyanin Content
4.3. cDNA Library Construction, Sequencing, and Transcriptome Assembly
4.4. Functional Annotation and Classification
4.5. Differentially Expressed Genes Analysis
4.6. Phylogenetic Analysis
4.7. Quantitative Real-Time PCR Analyses
4.8. Effect of Shading Treatment on Pigment Accumulation and Gene Expression
4.9. Identification SSRs in ESTs and Primer Design
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Davies, K.M.; Albert, N.W.; Schwinn, K.E. From landing lights to mimicry: The molecular regulation of flower colouration and mechanisms for pigmentation patterning. Funct. Plant Biol. 2012, 39, 619–638. [Google Scholar] [CrossRef]
- Yamagishi, M.; Toda, S.; Tasaki, K. The novel allele of the LhMYB12 gene is involved in splatter-type spot formation on the flower tepals of Asiatic hybrid lilies (Lilium spp.). New Phytol. 2014, 201, 1009–1020. [Google Scholar] [CrossRef] [PubMed]
- Winkel-Shirley, B. Flavonoid biosynthesis. A colorful model for genetics, biochemistry, cell biology, and biotechnology. Plant Physiol. 2001, 126, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Grotewold, E. The genetics and biochemistry of floral pigments. Annu. Rev. Plant Biol. 2006, 57, 761–780. [Google Scholar] [CrossRef] [PubMed]
- Koes, R.; Verweij, W.; Quattrocchio, F. Flavonoids: A colorful model for the regulation and evolution of biochemical pathways. Trends Plant Sci. 2005, 10, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Ferreyra, M.L.F.; Rius, S.P.; Casati, P. Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front. Plant Sci. 2012. [Google Scholar] [CrossRef]
- Nakatsuka, A.; Izumi, Y.; Yamagishi, M. Spatial and temporal expression of chalcone synthase and dihydroflavonol 4-reductase genes in the Asiatic hybrid lily. Plant Sci. 2003, 165, 759–767. [Google Scholar] [CrossRef]
- Lai, Y.S.; Shimoyamada, Y.; Nakayama, M.; Yamagishi, M. Pigment accumulation and transcription of LhMYB12 and anthocyanin biosynthesis genes during flower development in the Asiatic hybrid lily (Lilium spp.). Plant Sci. 2012, 193, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, M.; Shimoyamada, Y.; Nakatsuka, T.; Masuda, K. Two R2R3-MYB Genes, Homologs of petunia AN2, regulate anthocyanin biosyntheses in flower tepals, tepal spots and leaves of Asiatic hybrid lily. Plant Cell Physiol. 2010, 51, 463–474. [Google Scholar] [CrossRef] [PubMed]
- Berardini, T.Z.; Mundodi, S.; Reiser, L.; Huala, E.; Garcia-Hernandez, M.; Zhang, P.F.; Mueller, L.A.; Yoon, J.; Doyle, A.; Lander, G.; et al. Functional annotation of the Arabidopsis genome using controlled vocabularies. Plant Physiol. 2004, 135, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.Q.; Jiang, Y.; Ning, C.L.; Meng, J.S.; Lin, S.S.; Ding, W.; Tao, J. Transcriptome sequencing of a chimaera reveals coordinated expression of anthocyanin biosynthetic genes mediating yellow formation in herbaceous peony (Paeonia lactiflora Pall.). BMC Genom. 2014. [Google Scholar] [CrossRef] [PubMed]
- Takos, A.M.; Jaffe, F.W.; Jacob, S.R.; Bogs, J.; Robinson, S.P.; Walker, A.R. Light-induced expression of a MYB gene regulates anthocyanin biosynthesis in red apples. Plant Physiol. 2006, 142, 1216–1232. [Google Scholar] [CrossRef] [PubMed]
- Ban, Y.; Honda, C.; Hatsuyama, Y.; Igarashi, M.; Bessho, H.; Moriguchi, T. Isolation and functional analysis of a MYB transcription factor gene that is a key regulator for the development of red coloration in apple skin. Plant Cell Physiol. 2007, 48, 958–970. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.R.; Lee, E.; Bogs, J.; McDavid, D.A.J.; Thomas, M.R.; Robinson, S.P. White grapes arose through the mutation of two similar and adjacent regulatory genes. Plant J. 2007, 49, 772–785. [Google Scholar] [CrossRef] [PubMed]
- Palapol, Y.; Ketsa, S.; Kui, L.W.; Ferguson, I.B.; Allan, A.C. A MYB transcription factor regulates anthocyanin biosynthesis in mangosteen (Garcinia mangostana L.) fruit during ripening. Planta 2009, 229, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Niu, S.S.; Xu, C.J.; Zhang, W.S.; Zhang, B.; Li, X.; Lin-Wang, K.; Ferguson, I.B.; Allan, A.C.; Chen, K.S. Coordinated regulation of anthocyanin biosynthesis in Chinese bayberry (Myrica rubra) fruit by a R2R3 MYB transcription factor. Planta 2010, 231, 887–899. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.Q.; Wang, Y.L.; Yang, S.; Xu, Y.T.; Chen, X.S. Anthocyanin biosynthesis in pears is regulated by a R2R3-MYB transcription factor PyMYB10. Planta 2010, 232, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Lai, B.; Li, X.J.; Hu, B.; Qin, Y.H.; Huang, X.M.; Wang, H.C.; Hu, G.B. LcMYB1 Is a Key Determinant of Differential Anthocyanin Accumulation among Genotypes, Tissues, Developmental Phases and ABA and Light Stimuli in Litchi chinensis. PLoS ONE 2014. [Google Scholar] [CrossRef] [PubMed]
- Cone, K.C.; Burr, F.A.; Burr, B. Molecular analysis of the maize anthocyanin regulatory locus C1. Proc. Natl. Acad. Sci. USA 1986, 83, 9631–9635. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.Q.; Tao, J. Recent advances on the development and regulation of flower color in ornamental plants. Front. Plant Sci. 2015. [Google Scholar] [CrossRef] [PubMed]
- Azuma, A.; Yakushiji, H.; Koshita, Y.; Kobayashi, S. Flavonoid biosynthesis-related genes in grape skin are differentially regulated by temperature and light conditions. Planta 2012, 236, 1067–1080. [Google Scholar] [CrossRef] [PubMed]
- Shinomiya, R.; Fujishima, H.; Muramoto, K.; Shiraishi, M. Impact of temperature and sunlight on the skin coloration of the “Kyoho” table grape. Sci. Hortic. 2015, 193, 77–83. [Google Scholar] [CrossRef]
- Ubi, B.E.; Honda, C.; Bessho, H.; Kondo, S.; Wada, M.; Kobayashi, S.; Moriguchi, T. Expression analysis of anthocyanin biosynthetic genes in apple skin: Effect of UV-B and temperature. Plant Sci. 2006, 170, 571–578. [Google Scholar] [CrossRef]
- Li, Y.Y.; Mao, K.; Zhao, C.; Zhao, X.Y.; Zhang, H.L.; Shu, H.R.; Hao, Y.J. MdCOP1 ubiquitin E3 ligases interact with MdMYB1 to regulate light-induced anthocyanin biosynthesis and red fruit coloration in apple. Plant Physiol. 2012, 160, 1011–1022. [Google Scholar] [CrossRef] [PubMed]
- Peng, T.; Saito, T.; Honda, C.; Ban, Y.; Kondo, S.; Liu, J.H.; Hatsuyama, Y.; Moriguchi, T. Screening of UV-B-induced genes from apple peels by SSH: Possible involvement of MdCOP1-mediated signaling cascade genes in anthocyanin accumulation. Physiol. Plant. 2013, 148, 432–444. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Qian, M.J.; Yu, B.; Teng, Y.W. Effect of fruit maturity on UV-B-induced post-harvest anthocyanin accumulation in red Chinese sand pear. Acta Physiol. Plant. 2013, 35, 2857–2866. [Google Scholar] [CrossRef]
- Li, Y.Y.; Mao, K.; Zhao, C.; Zhao, X.Y.; Zhang, R.F.; Zhang, H.L.; Shu, H.R.; Hao, Y.J. Molecular cloning and functional analysis of a blue light receptor gene MdCRY2 from apple (Malus domestica). Plant Cell Rep. 2013, 32, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Kadomura-Ishikawa, Y.; Miyawaki, K.; Noji, S.; Takahashi, A. Phototropin 2 is involved in blue light-induced anthocyanin accumulation in Fragaria x ananassa fruits. J. Plant Res. 2013, 126, 847–857. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.Y.; Cao, S.F.; Chen, W.; Yang, Z.F. Blue light induced anthocyanin accumulation and expression of associated genes in Chinese bayberry fruit. Sci. Hortic. 2014, 179, 98–102. [Google Scholar] [CrossRef]
- Lee, S.I.; Park, K.C.; Song, Y.S.; Son, J.H.; Kwon, S.J.; Na, J.K.; Kim, J.H.; Kim, N.S. Development of expressed sequence tag derived-simple sequence repeats in the genus Lilium. Genes Genom. 2011, 33, 727–733. [Google Scholar] [CrossRef]
- Shahin, A.; van Kaauwen, M.; Esselink, D.; Bargsten, J.W.; van Tuyl, J.M.; Visser, R.G.F.; Arens, P. Generation and analysis of expressed sequence tags in the extreme large genomes Lilium and Tulipa. BMC Genom. 2012. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.X.; Ge, L.; Liu, C.; Ming, J. The development of EST-SSR markers in Lilium regale and their cross-amplification in related species. Euphytica 2013, 189, 393–419. [Google Scholar] [CrossRef]
- Du, F.; Jiang, J.; Jia, H.M.; Zhao, X.Y.; Wang, W.H.; Gao, Q.K.; Mao, W.H.; Wu, Y.; Zhang, L.; Grierson, D.; et al. Selection of generally applicable SSR markers for evaluation of genetic diversity and identity in Lilium. Biochem. Syst. Ecol. 2015, 61, 278–285. [Google Scholar] [CrossRef]
- Cunja, V.; Mikulic-Petkovsek, M.; Stampar, F.; Schmitzer, V. Compound identification of selected rose species and cultivars: An insight to petal and leaf phenolic profiles. J. Am. Soc. Hortic. Sci. 2014, 139, 157–166. [Google Scholar]
- Liu, S.S.; Chen, J.; Li, S.C.; Zeng, X.; Meng, Z.X.; Guo, S.X. Comparative Transcriptome analysis of genes involved in GA-GID1-DELLA regulatory module in symbiotic and asymbiotic seed germination of Anoectochilus roxburghii (Wall.) Lindl. (Orchidaceae). Int. J. Mol. Sci. 2015, 16, 30190–30203. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.D.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.K.; Wang, Y.B.; Liu, Z.X.; Cheng, H.; Xue, Y. HemI: A toolkit for illustrating heatmaps. PLoS ONE 2014. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Protein | No. All a | No. Up b | No. Down c |
|---|---|---|---|---|
| CHS | Chalcone synthase | 38 | 11 | 0 |
| CHI | Chalcone isomerase | 1 | 1 | 0 |
| F3H | Flavanone 3-hydroxylase | 2 | 1 | 0 |
| F3′H | Flavonoid 3′-hydroxylase | 4 | 1 | 0 |
| DFR | Dihydroflavonol 4-reductase | 3 | 1 | 0 |
| ANS | Anthocyanidin synthase | 3 | 0 | 0 |
| UFGT | Anthocyanidin 3-O-glucosyltransfersae | 5 | 3 | 0 |
| 3RT | anthocyanidin-3-glucoside rhamnosyltransferase | 1 | 1 | 0 |
| R2R3-MYB | R2R3-MYB transcription factors | 28 | 8 | 0 |
| bHLH | Basic helix–loop–helix transcription factors | 19 | 8 | 11 |
| WD | WD-repeat proteins | 19 | 3 | 6 |
| Annotation | Unigene ID | S1 FPKM | S2 FPKM |
|---|---|---|---|
| CHS | c124520_g1 | 1.07 | 1255.60 |
| c114453_g1 | 1.91 | 1495.41 | |
| c122851_g2 | 2.16 | 1996.29 | |
| c124520_g2 | 2.15 | 2926.54 | |
| c201597_g1 | 1.17 | 106.51 | |
| c115882_g3 | 0.96 | 614.86 | |
| c122851_g1 | 0.00 | 342.93 | |
| c115882_g2 | 0.95 | 375.32 | |
| c81738_g1 | 0.00 | 12.03 | |
| c114453_g2 | 0.06 | 3.35 | |
| c124865_g1 | 0.07 | 1.22 | |
| CHI | c123012_g2 | 93.54 | 743.77 |
| F3H | c121630_g1 | 25.79 | 1134.48 |
| F3'H | c113932_g1 | 25.64 | 1099.95 |
| DFR | c124316_g1 | 8.43 | 5147.89 |
| UFGT | c113956_g1 | 5.67 | 3286.62 |
| c114752_g1 | 8.03 | 171.41 | |
| c92724_g2 | 13.57 | 49.55 | |
| 3RT | c117809_g1 | 1.20 | 584.93 |
| R2R3-MYB | c51332_g1 | 2.81 | 264.36 |
| c72124_g1 | 0.00 | 1.75 | |
| c116256_g1 | 2.26 | 10.81 | |
| c36531_g1 | 2.47 | 7.92 | |
| c121270_g3 | 42.98 | 256.59 | |
| c96386_g1 | 65.52 | 182.05 | |
| c116626_g1 | 2.21 | 5.60 | |
| c104994_g2 | 0.00 | 16.16 |
| Parameter | Value | Parameter | Value |
|---|---|---|---|
| SSR Identification | Distribution to Different Repeat Type Classes | ||
| Total number of sequences examined | 295,787 | Mononucleotide | 8163 (54.2%) |
| Total size of examined sequences (bp) | 160,816,025 | Dinucleotide | 4446 (29.5%) |
| Total number of identified SSRs | 15,048 | Trinucleotide | 2309 (15.3%) |
| Number of SSR containing sequences | 13,710 | Tetranucleotide | 96 (0.6%) |
| Number of sequences containing more than one SSR | 1201 | Pentanucleotide | 15 (0.1%) |
| Number of SSRs present in compound formation | 444 | Hexanucleotide | 19 (0.1%) |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, L.; Yang, P.; Yuan, S.; Feng, Y.; Xu, H.; Cao, Y.; Ming, J. Transcriptome Analysis Identifies Key Candidate Genes Mediating Purple Ovary Coloration in Asiatic Hybrid Lilies. Int. J. Mol. Sci. 2016, 17, 1881. https://doi.org/10.3390/ijms17111881
Xu L, Yang P, Yuan S, Feng Y, Xu H, Cao Y, Ming J. Transcriptome Analysis Identifies Key Candidate Genes Mediating Purple Ovary Coloration in Asiatic Hybrid Lilies. International Journal of Molecular Sciences. 2016; 17(11):1881. https://doi.org/10.3390/ijms17111881
Chicago/Turabian StyleXu, Leifeng, Panpan Yang, Suxia Yuan, Yayan Feng, Hua Xu, Yuwei Cao, and Jun Ming. 2016. "Transcriptome Analysis Identifies Key Candidate Genes Mediating Purple Ovary Coloration in Asiatic Hybrid Lilies" International Journal of Molecular Sciences 17, no. 11: 1881. https://doi.org/10.3390/ijms17111881
APA StyleXu, L., Yang, P., Yuan, S., Feng, Y., Xu, H., Cao, Y., & Ming, J. (2016). Transcriptome Analysis Identifies Key Candidate Genes Mediating Purple Ovary Coloration in Asiatic Hybrid Lilies. International Journal of Molecular Sciences, 17(11), 1881. https://doi.org/10.3390/ijms17111881