
Low Concentration of Exogenous Carbon Monoxide Modulates Radiation-Induced Bystander Effect in Mammalian Cell Cluster Model

_Kwan_Ngok_Yu.png)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. CO (CORM-2) Decreased DSB Formation in the Bystander but Not Irradiated Cell Population
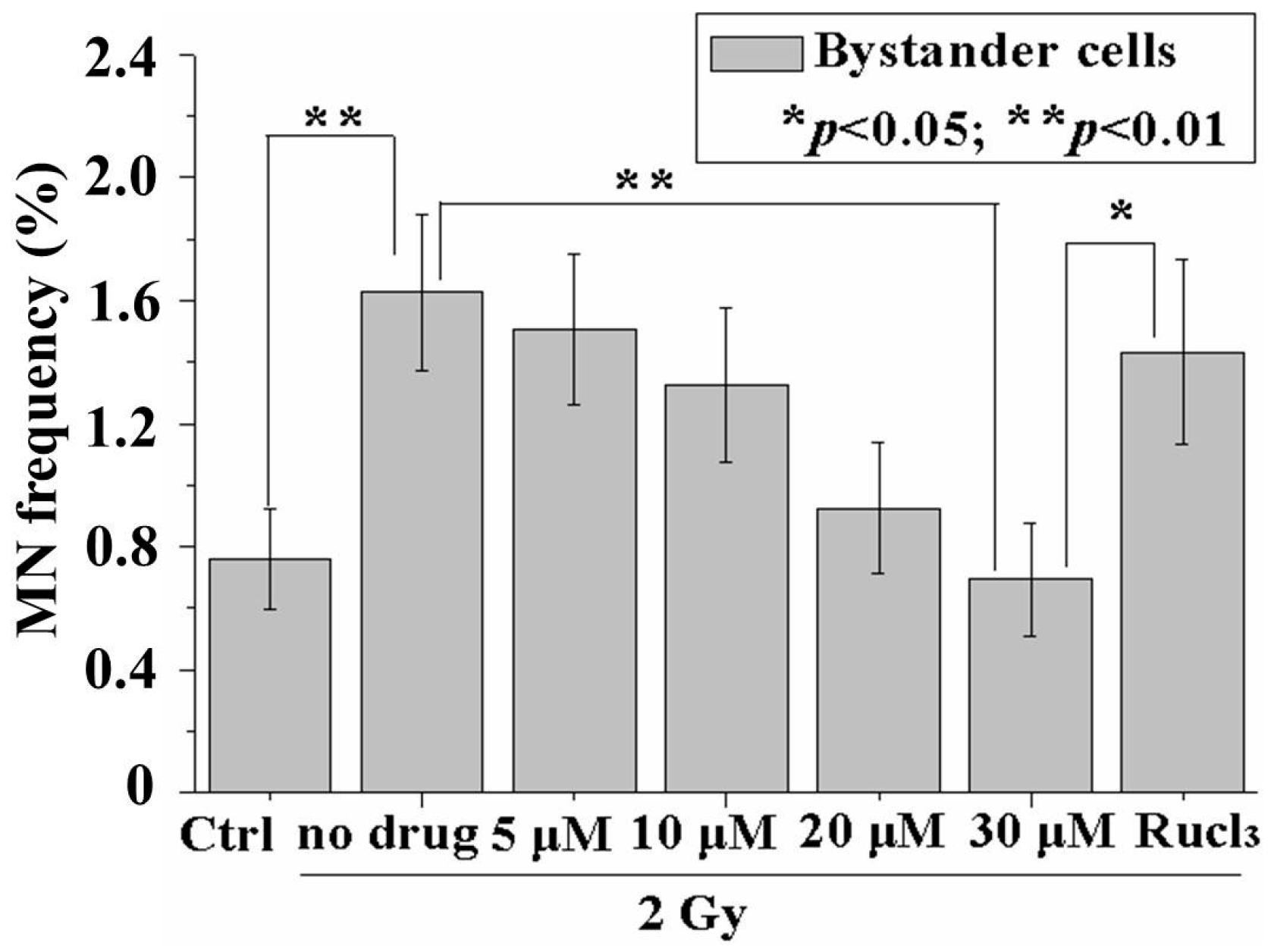
2.2. CO (CORM-2) Decreased MN Formation in the Bystander Cell Population
2.3. CO (CORM-2) Did Not Affect p53BP1 Formation and MN Frequency in Irradiated Cells
2.4. CO (CORM-2) Inhibited RIBE-Induced Cell Proliferation but Did Not Affect Irradiated Cells
2.5. Inhibition of iNOS or COX-2 Decreased RIBE-Induced p53BP1 Formation and MN Frequency
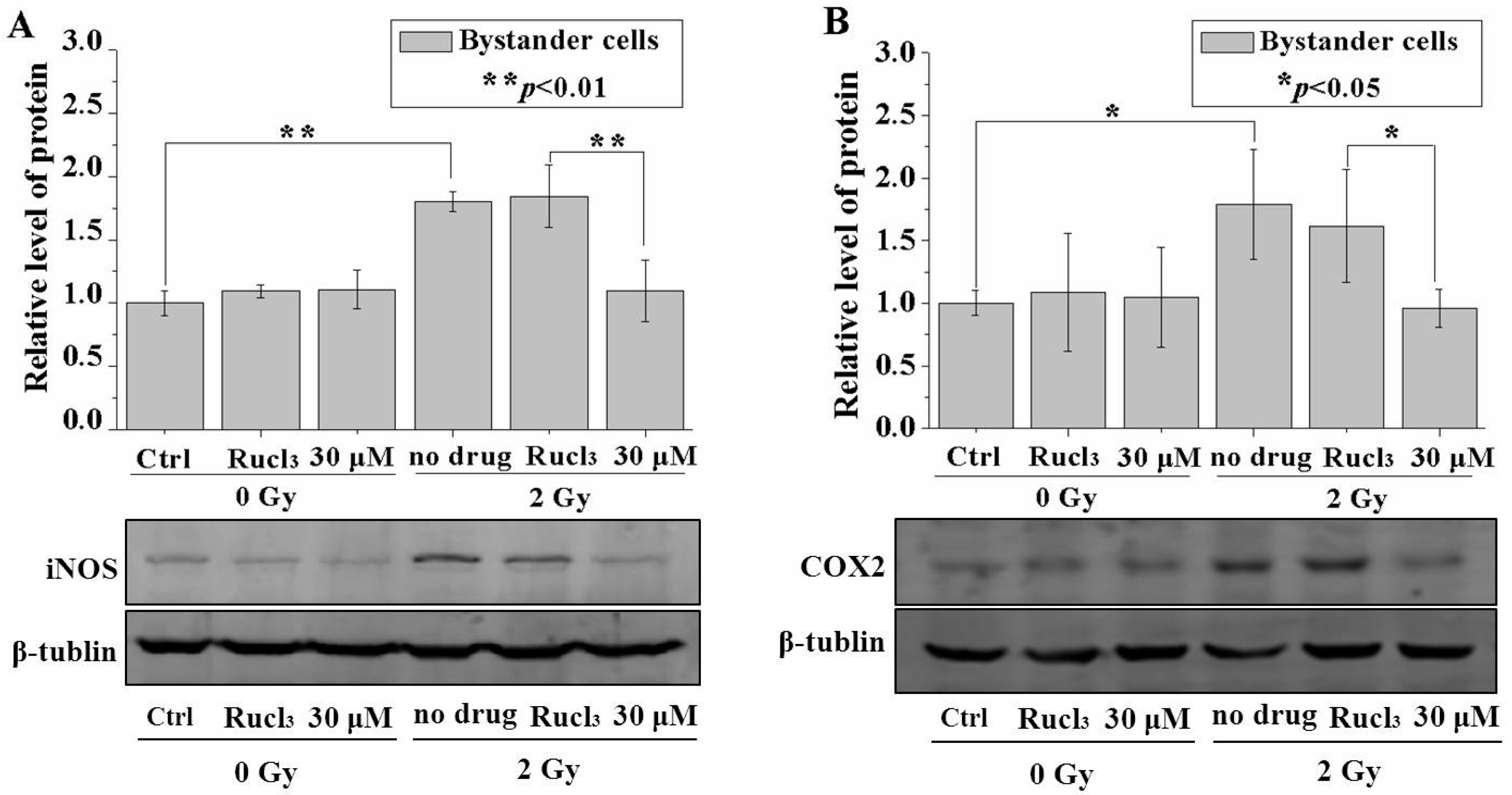
2.6. CO (CORM-2) Decreased Expression of iNOS and COX-2
3. Discussion
4. Materials and Methods
4.1. Cell Culture and X-ray Irradiation
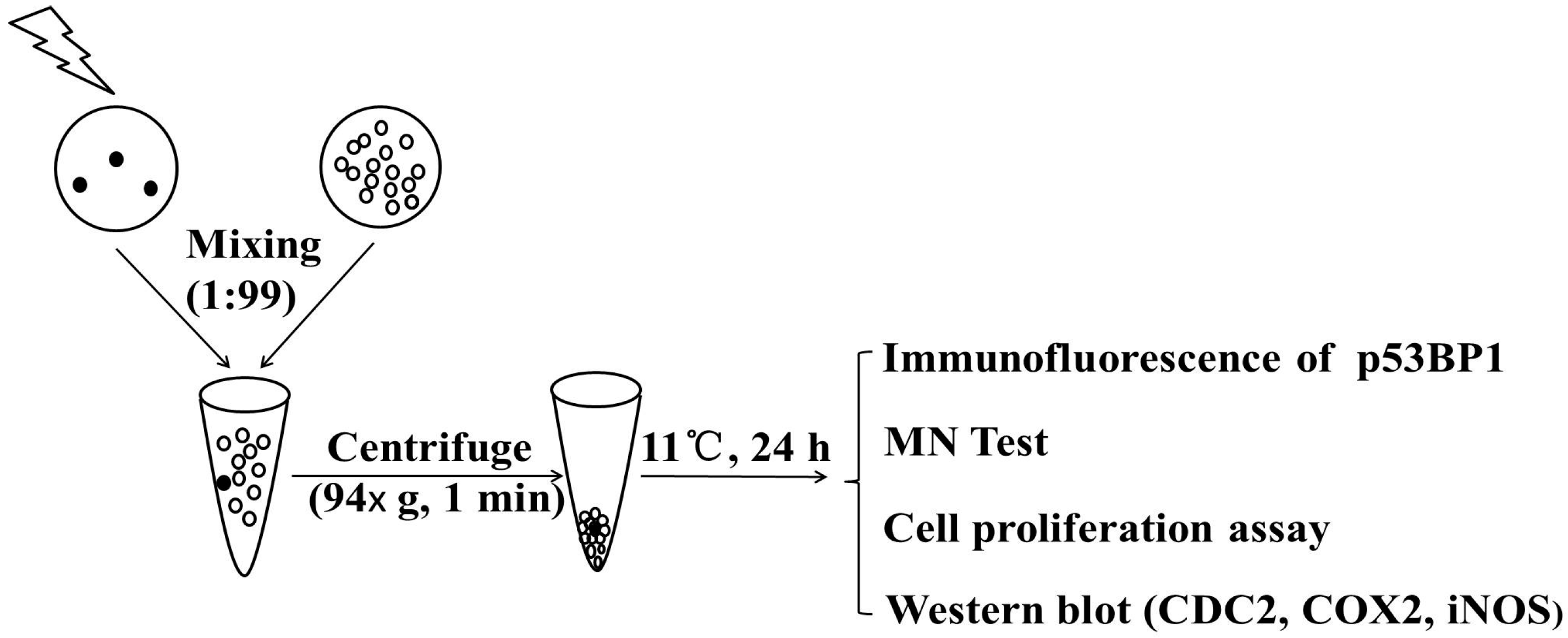
4.2. Preparation of Cell Cluster
4.3. CO (CORM-2) Treatment
4.4. Immunofluorescence of p53 Binding Protein 1 (p53BP1)
4.5. MN Assay
4.6. Cell Proliferation Assay
4.7. Western Blot Analyses
4.8. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| RIBE | radiation-induced bystander effect |
| CO | carbon monoxide |
| CORM-2 | CO releasing molecule, tricarbonyldichlororuthenium |
| DSB | DNA double-strand breaks |
| MN | micronucleus |
| iNOS | inducible nitric oxide synthase |
| COX-2 | cyclooxygenase-2 |
| DMSO | dimethyl sulfoxide |
| p53BP1 | p53 binding protein 1 |
| RuCl3 | ruthenium trichloride |
| CDC2 | cell division cycle 2 |
| NS 398 | N-[2-(cyclohexyloxy)-4-nitrophenyl]-methanesulfonamide |
| AG | aminoguanidine |
| MAPK | mitogen activated protein kinase |
References
- Prise, K.M.; O’Sullivan, J.M. Radiation-induced bystander signalling in cancer therapy. Nat. Rev. Cancer 2009, 9, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Baskar, R. Emerging role of radiation induced bystander effects: Cell communications and carcinogenesis. Genome Integr. 2010, 1, 13. [Google Scholar] [CrossRef] [PubMed]
- Morgan, W.F.; Sowa, M.B. Non-targeted effects induced by ionizing radiation: Mechanisms and potential impact on radiation induced health effects. Cancer Lett. 2015, 356, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Randers-Pehrson, G.; Waldren, C.A.; Vannais, D.; Hall, E.J.; Hei, T.K. Induction of a bystander mutagenic effect of α particles in mammalian cells. Proc. Natl. Acad. Sci. USA 2000, 97, 2099–2104. [Google Scholar] [CrossRef] [PubMed]
- Persaud, R.; Zhou, H.; Baker, S.E.; Hei, T.K.; Hall, E.J. Assessment of low linear energy transfer radiation-induced bystander mutagenesis in a three-dimensional culture model. Cancer Res. 2005, 65, 9876–9882. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Wu, L.; Han, W.; Zhang, L.; Chen, S.; Xu, A.; Hei, T.K.; Yu, Z. The time and spatial effects of bystander response in mammalian cells induced by low dose radiation. Carcinogenesis 2006, 27, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Koturbash, I.; Rugo, R.E.; Hendricks, C.A.; Loree, J.; Thibault, B.; Kutanzi, K.; Pogribny, I.; Yanch, J.C.; Engelward, B.P.; Kovalchuk, O. Irradiation induces DNA damage and modulates epigenetic effectors in distant bystander tissue in vivo. Oncogene 2006, 25, 4267–4275. [Google Scholar] [CrossRef] [PubMed]
- Iyer, R.; Lehnert, B.E.; Svensson, R. Factors underlying the cell growth-related bystander responses to α particles. Cancer Res. 2000, 60, 1290–1298. [Google Scholar] [PubMed]
- Shao, C.; Folkard, M.; Michael, B.D.; Prise, K.M. Targeted cytoplasmic irradiation induces bystander responses. Proc. Natl. Acad. Sci. USA 2004, 101, 13495–13500. [Google Scholar] [CrossRef] [PubMed]
- Sigg, M.; Crompton, N.E.; Burkart, W. Enhanced neoplastic transformation in an inhomogeneous radiation field: An effect of the presence of heavily damaged cells. Radiat. Res. 1997, 148, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, M.; Pasquali, E.; Leonardi, S.; Tanori, M.; Rebessi, S.; di Majo, V.; Pazzaglia, S.; Toni, M.P.; Pimpinella, M.; Covelli, V. Oncogenic bystander radiation effects in patched heterozygous mouse cerebellum. Proc. Natl. Acad. Sci. USA 2008, 105, 12445–12450. [Google Scholar] [CrossRef] [PubMed]
- Bishayee, A.; Hill, H.Z.; Stein, D.; Rao, D.V.; Howell, R.W. Free radical-initiated and gap junction-mediated bystander effect due to nonuniform distribution of incorporated radioactivity in a three-dimensional tissue culture model. Radiat. Res. 2001, 155, 335–344. [Google Scholar] [CrossRef]
- Azzam, E.I.; de Toledo, S.M.; Raaphorst, G.P.; Mitchel, R.E. Low-dose ionizing radiation decreases the frequency of neoplastic transformation to a level below the spontaneous rate in C3H 10T1/2 cells. Radiat. Res. 1996, 146, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.W.; Wang, J.; Bucking, C.P.; Mothersill, C.E.; Seymour, C.B. Evidence for a protective response by the gill proteome of rainbow trout exposed to X-ray induced bystander signals. Proteomics 2007, 7, 4171–4180. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.W.; Wang, J.; Schültke, E.; Seymour, C.B.; Bräuer-Krisch, E.; Laissue, J.A.; Mothersill, C.E. Proteomic changes in the rat brain induced by homogenous irradiation and by the bystander effect resulting from high energy synchrotron X-ray microbeams. Int. J. Radiat. Biol. 2013, 89, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, H.; Takahashi, A.; Ohnishi, T. Radiation-induced adaptive responses and bystander effects. Biol. Sci. Space 2004, 18, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Mothersill, C.; Seymour, C. Radiation-induced bystander effects: Evidence for an adaptive response to low dose exposures? Dose-Response 2006, 4, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Mothersill, C.; Seymour, C. Radiation-induced bystander effects: Are they good, bad or both? Med. Confl. Surviv. 2010, 21, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Wang, R. Carbon monoxide: Endogenous production, physiological functions, and pharmacological applications. Pharmacol. Rev. 2005, 57, 585–630. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Wu, L.; Chen, S.; Yu, K.N. Exogenous carbon monoxide protects the bystander chinese hamster ovary cells in mixed coculture system after alpha-particle irradiation. Carcinogenesis 2010, 31, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Yu, K.N.; Wu, L.J.; Wu, Y.C.; Wang, H.Z. Mechanism of protection of bystander cells by exogenous carbon monoxide: Impaired response to damage signal of radiation-induced bystander effect. Mutat. Res. 2011, 709–710, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Tong, L.; Yu, K.N.; Bao, L.; Wu, W.; Wang, H.; Han, W. Low concentration of exogenous carbon monoxide protects mammalian cells against proliferation induced by radiation-induced bystander effect. Mutat. Res. 2014, 759, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Gerashchenko, B.I.; Yamagata, A.; Oofusa, K.; Yoshizato, K.; de Toledo, S.M.; Howell, R.W. Proteome analysis of proliferative response of bystander cells adjacent to cells exposed to ionizing radiation. Proteomics 2007, 7, 2000–2008. [Google Scholar] [CrossRef] [PubMed]
- Shao, C.; Stewart, V.; Folkard, M.; Michael, B.D.; Prise, K.M. Nitric Oxide-Mediated Signaling in the Bystander Response of Individually Targeted Glioma Cells. Cancer Res. 2003, 63, 8437–8442. [Google Scholar] [PubMed]
- Zhou, H.; Ivanov, V.N.; Gillespie, J.; Geard, C.R.; Amundson, S.A.; Brenner, D.J.; Yu, Z.; Lieberman, H.B.; Hei, T.K. Mechanism of radiation-induced bystander effect: Role of the cyclooxygenase-2 signaling pathway. Proc. Natl. Acad. Sci. USA 2005, 102, 14641–14646. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, H.; Little, J.B. Induction of Sister Chromatid Exchanges by Extremely Low Doses of α-Particles. Cancer Res. 1992, 52, 6394–6396. [Google Scholar] [PubMed]
- Gerashchenko, B.I.; Howell, R.W. Flow cytometry as a strategy to study radiation-induced bystander effects in co-culture systems. Cytometry A 2003, 54, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Mothersill, C.; Seymour, R.J.; Seymour, C.B. Increased radiosensitivity in cells of two human cell lines treated with bystander medium from irradiated repair deficient cells. Radiat. Res. 2006, 165, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Gerashchenko, B.I.; Howell, R.W. Cell proximity is a prerequisite for the proliferative response of bystander cells co-cultured with cells irradiated with γ-rays. Cytometry A 2003, 56, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Smythe, C.; Newport, J.W. Coupling of mitosis to the completion of S phase in Xenopus occurs via modulation of the tyrosine kinase that phosphorylates p34cdc2. Cell 1992, 68, 787–797. [Google Scholar] [CrossRef]
- Azzam, E.I.; de Toledo, S.M.; Pykett, M.J.; Nagasawa, H.; Little, J.B. CDC2 is down-regulated by ionizing radiation in a p53-dependent manner. Cell Growth Differ. 1997, 8, 1161–1169. [Google Scholar] [PubMed]
- Belyakov, O.V.; Mitchell, S.A.; Parikh, D.; Randers-Pehrson, G.; Marino, S.A.; Amundson, S.A.; Geard, C.R.; Brenner, D.J. Biological effects in unirradiated human tissue induced by radiation damage up to 1 mm away. Proc. Natl. Acad. Sci. USA 2005, 102, 14203–14208. [Google Scholar] [CrossRef] [PubMed]
- Sedelnikova, O.A.; Nakamura, A.; Kovalchuk, O.; Koturbash, I.; Mitchell, S.A.; Marino, S.A.; Brenner, D.J.; Bonner, W.M. DNA double-strand breaks form in bystander cells after microbeam irradiation of three-dimensional human tissue models. Cancer Res. 2007, 67, 4295–4302. [Google Scholar] [CrossRef] [PubMed]
- Persaud, R.; Zhou, H.; Hei, T.K.; Hall, E.J. Demonstration of a radiation-induced bystander effect for low dose low let beta-particles. Radiat. Environ. Biophys. 2007, 46, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Hei, T.K.; Zhou, H.; Ivanov, V.N.; Hong, M.; Lieberman, H.B.; Brenner, D.J.; Amundson, S.A.; Geard, C.R. Mechanism of radiation-induced bystander effects: A unifying model. J. Pharm. Pharmacol. 2008, 60, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Shao, C.; Folkard, M.; Prise, K.M. Role of tgf-beta1 and nitric oxide in the bystander response of irradiated glioma cells. Oncogene 2008, 27, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Azzam, E.I.; de Toledo, S.M.; Spitz, D.R.; Little, J.B. Oxidative metabolism modulates signal transduction and micronucleus formation in bystander cells from α-particle-irradiated normal human fibroblast cultures. Cancer Res. 2002, 62, 5436–5442. [Google Scholar] [PubMed]
- Zhou, H.; Ivanov, V.N.; Lien, Y.C.; Davidson, M.; Hei, T.K. Mitochondrial function and nuclear factor-kappab-mediated signaling in radiation-induced bystander effects. Cancer Res. 2008, 68, 2233–2240. [Google Scholar] [CrossRef] [PubMed]
- Cavicchi, M.; Gibbs, L.; Whittle, B.J. Inhibition of inducible nitric oxide synthase in the human intestinal epithelial cell line, DLD-1, by the inducers of hemeoxygenase 1, bismuth salts, heme, and nitric oxide donors. Gut 2000, 47, 771–778. [Google Scholar] [CrossRef] [PubMed]
- Zuckerbraun, B.S.; Otterbein, L.E.; Boyle, P.; Jaffe, R.; Upperman, J.; Zamora, R.; Ford, H.R. Carbon monoxide protects against the development of experimental necrotizing enterocolitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 289, G607–G613. [Google Scholar] [CrossRef] [PubMed]
- Chien, P.T.; Lin, C.C.; Hsiao, L.D.; Yang, C.M. Induction of ho-1 by carbon monoxide releasing molecule-2 attenuates thrombin-induced cox-2 expression and hypertrophy in primary human cardiomyocytes. Toxicol. Appl. Pharmacol. 2015, 289, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Suh, G.Y.; Jin, Y.; Yi, A.K.; Wang, X.M.; Choi, A.M. CCAAT/enhancer-binding protein mediates carbon monoxide-induced suppression of cyclooxygenase-2. Am. J. Respir. Cell Mol. Biol. 2006, 35, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.J.; Jeong, S.O.; Zheng, M.; Chen, Y.; Cho, G.J.; Chung, H.T.; Joe, Y. Carbon monoxide attenuates dextran sulfate sodium-induced colitis via inhibition of GSK-3β signaling. Oxidative Med. Cell. Longev. 2013, 2013, 210563. [Google Scholar] [CrossRef] [PubMed]
- Motterlini, R.; Clark, J.E.; Foresti, R.; Sarathchandra, P.; Mann, B.E.; Green, C.J. Carbon monoxide-releasing molecules: Characterization of biochemical and vascular activities. Circ. Res. 2002, 90, E17–E24. [Google Scholar] [CrossRef] [PubMed]
- Tartier, L.; Gilchrist, S.; Burdak-Rothkamm, S.; Folkard, M.; Prise, K.M. Cytoplasmic irradiation induces mitochondrial-dependent 53bp1 protein relocalization in irradiated and bystander cells. Cancer Res. 2007, 67, 5872–5879. [Google Scholar] [CrossRef] [PubMed]






© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, W.; Nie, L.; Yu, K.N.; Wu, L.; Kong, P.; Bao, L.; Chen, G.; Yang, H.; Han, W. Low Concentration of Exogenous Carbon Monoxide Modulates Radiation-Induced Bystander Effect in Mammalian Cell Cluster Model. Int. J. Mol. Sci. 2016, 17, 2051. https://doi.org/10.3390/ijms17122051
Wu W, Nie L, Yu KN, Wu L, Kong P, Bao L, Chen G, Yang H, Han W. Low Concentration of Exogenous Carbon Monoxide Modulates Radiation-Induced Bystander Effect in Mammalian Cell Cluster Model. International Journal of Molecular Sciences. 2016; 17(12):2051. https://doi.org/10.3390/ijms17122051
Chicago/Turabian StyleWu, Wenqing, Lili Nie, K. N. Yu, Lijun Wu, Peizhong Kong, Lingzhi Bao, Guodong Chen, Haoran Yang, and Wei Han. 2016. "Low Concentration of Exogenous Carbon Monoxide Modulates Radiation-Induced Bystander Effect in Mammalian Cell Cluster Model" International Journal of Molecular Sciences 17, no. 12: 2051. https://doi.org/10.3390/ijms17122051
APA StyleWu, W., Nie, L., Yu, K. N., Wu, L., Kong, P., Bao, L., Chen, G., Yang, H., & Han, W. (2016). Low Concentration of Exogenous Carbon Monoxide Modulates Radiation-Induced Bystander Effect in Mammalian Cell Cluster Model. International Journal of Molecular Sciences, 17(12), 2051. https://doi.org/10.3390/ijms17122051