
Crystallization of Galectin-8 Linker Reveals Intricate Relationship between the N-terminal Tail and the Linker


Abstract
:1. Introduction
2. Results
2.1. Gal-8_1-186 Induces Erythrocyte Agglutination
2.2. SDS-PAGE of Protein Crystals
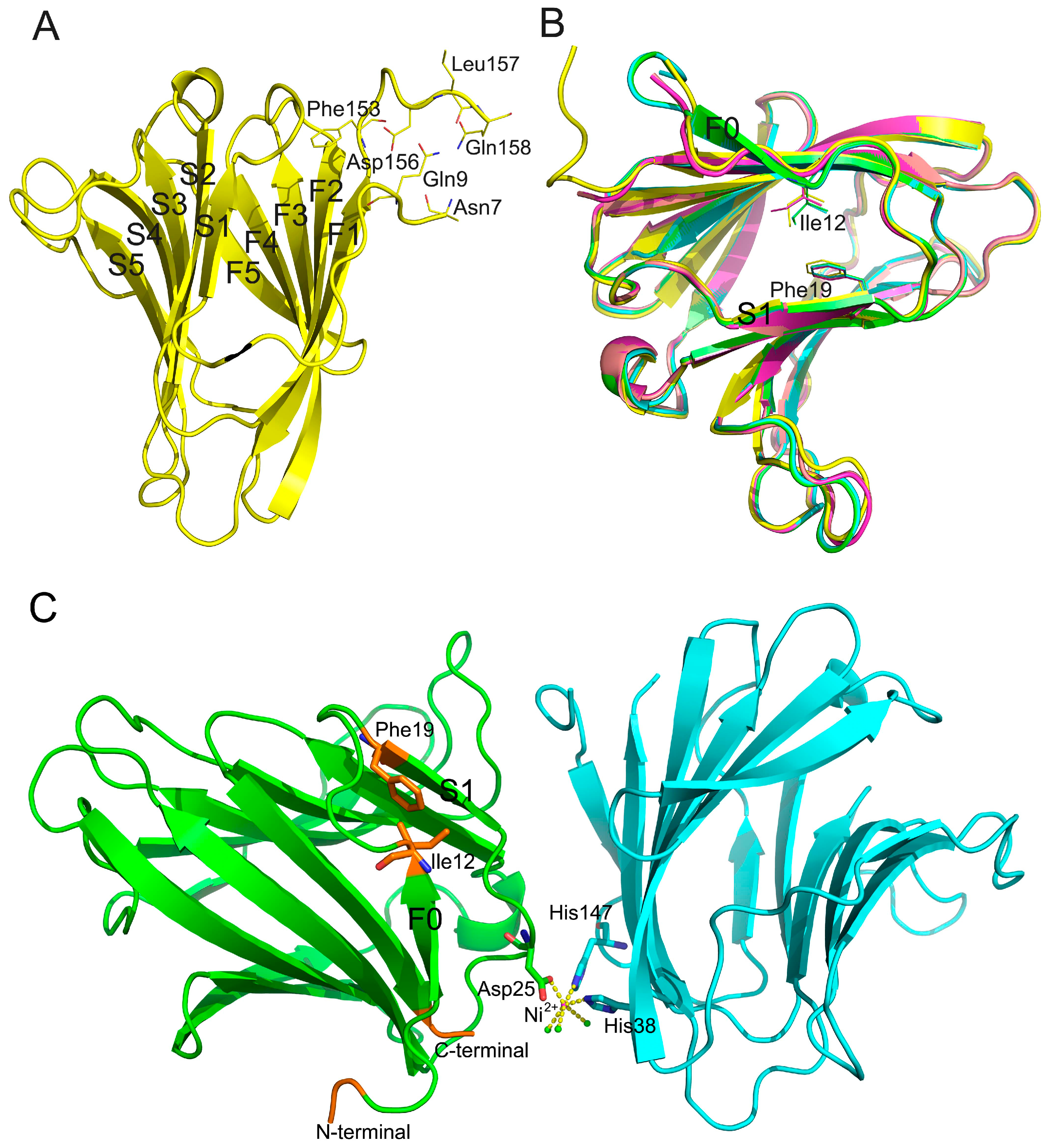
2.3. Description of the Gal-8_1-186 Structures
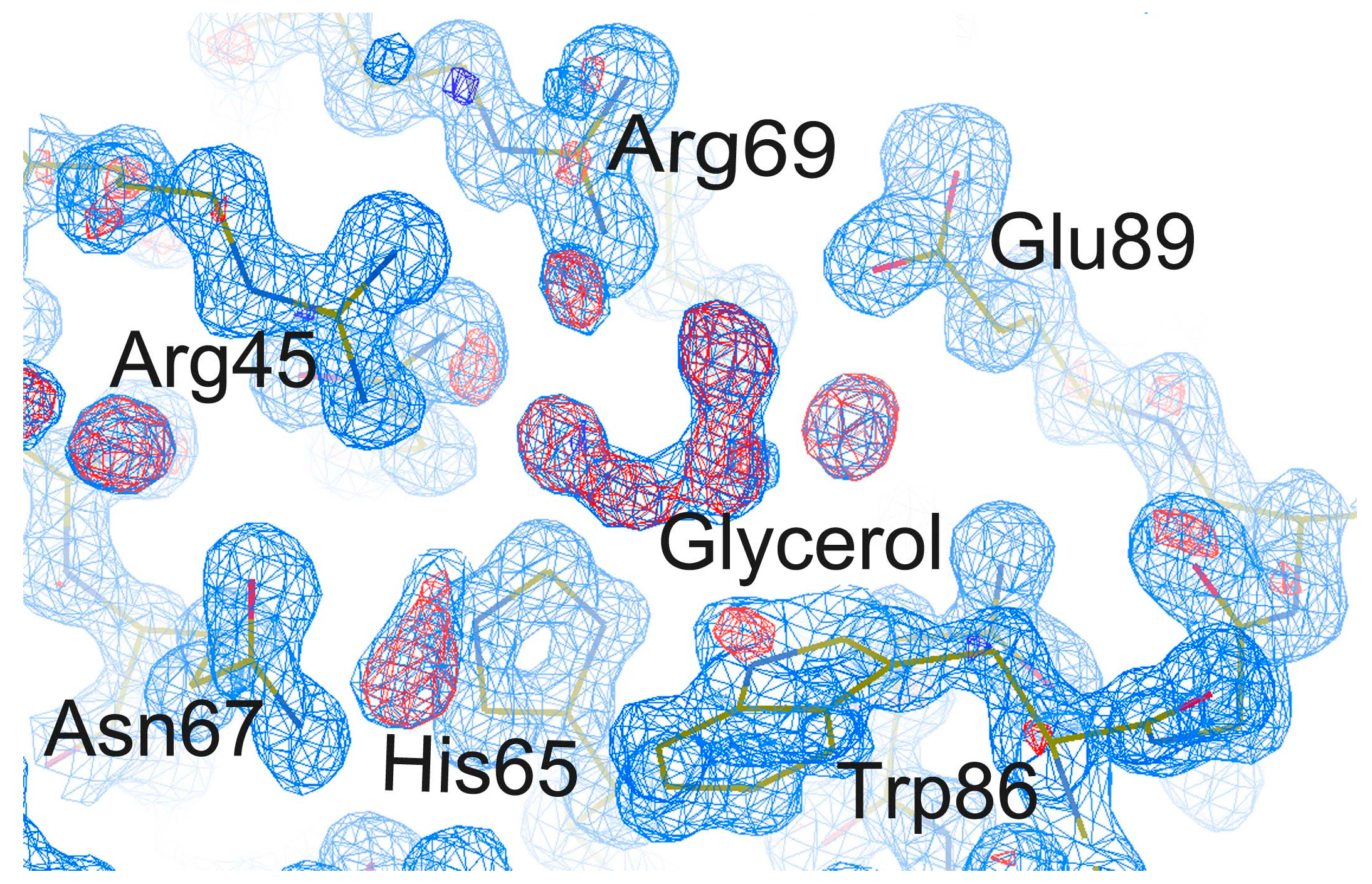
2.4. Effect of Cryoprotectants on Gal-8_1-186 Crystals
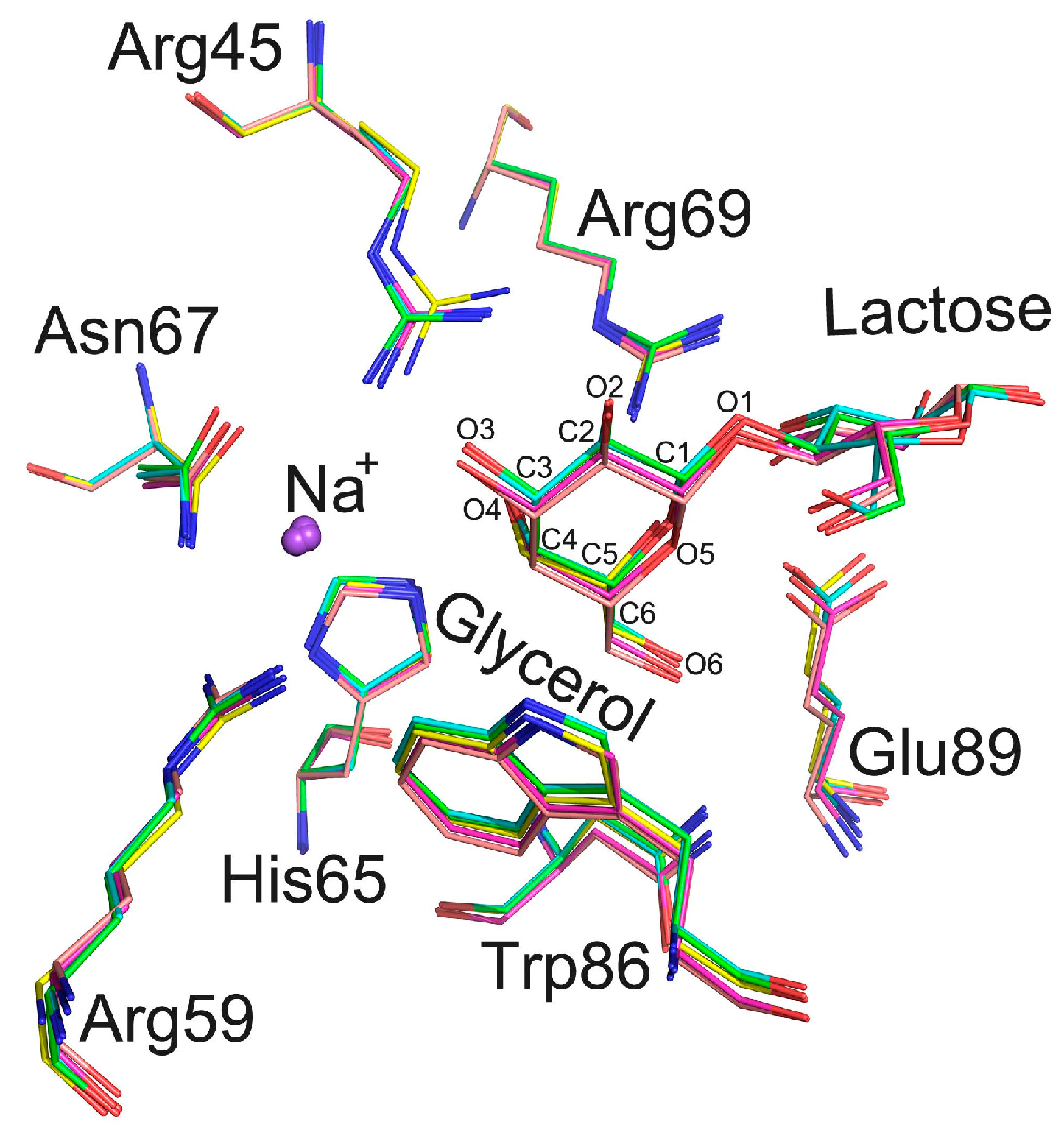
2.5. The Carbohydrate Binding Site
3. Discussion
4. Materials and Methods
4.1. Cloning, Expression, and Purification of Gal-8_1-186
4.2. Crystallization, Data Collection, and Structure Determination
4.3. Hemagglutination Assay
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| MAC | Minimum Agglutination Concentration |
| MIC | Minimum Inhibition Concentration |
| PCR | Polymerase Chain Reaction |
| CD | Circular Dichroism |
| CRD | Carbohydrate-Recognition Domain |
| Gal-8 | Galectin-8 |
| Gal-8_1-186 | The truncated protein of Galectin-8 contain 1 to 186 amino acid |
| Gal-8 N | N-terminal CRD of Galectin-8 |
| Gal-8 L | “long” Gal-8,with the longest linker peptide |
| Gal-8 M | “medium” Gal-8, with an intermediate linker region |
| SDS-PAGE | Sodium Dodecyl Sulfate Polyacrylamide Gel Electropheresis |
References
- Barondes, S.H.; Castronovo, V.; Cooper, D.N.; Cummings, R.D.; Drickamer, K.; Feizi, T.; Gitt, M.A.; Hirabayashi, J.; Hughes, C.; Kasai, K. Galectins: A family of animal β-galactoside-binding lectins. Cell 1994, 76, 597–598. [Google Scholar] [CrossRef]
- Lahm, H.; Andre, S.; Hoeflich, A.; Kaltner, H.; Siebert, H.C.; Sordat, B.; von der Lieth, C.W.; Wolf, E. Tumor galectinology: Insights into the complex network of a family of endogenous lectins. Glycoconj. J. 2004, 20, 227–228. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.T.; Rabinovich, G.A. Galectins as modulators of tumour progression. Nat. Rev. Cancer 2005, 5, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Massa, S.M.; Cooper, D.N.; Leffler, H.; Barondes, S.H. L-29, an endogenous lectin, binds to glycoconjugate ligands with positive cooperativity. Biochemistry 1993, 32, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.Y.; Rabinovich, G.A.; Liu, F.T. Galectins: Structure, function and therapeutic potential. Expert Rev. Mol. Med. 2008, 10, e17. [Google Scholar] [CrossRef] [PubMed]
- Kasai, K.; Hirabayashi, J. Galectins: A family of animal lectins that decipher glycocodes. J. Biochem. 1996, 119, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Cooper, D.N. Galectinomics: Finding themes in complexity. Biochim. Biophys. Acta 2002, 1572, 209–231. [Google Scholar] [CrossRef]
- Tasumi, S.; Vasta, G.R. A galectin of unique domain organization from hemocytes of the Eastern oyster (Crassostrea virginica) is a receptor for the protistan parasite Perkinsus marinus. J. Immunol. 2007, 179, 3086–3098. [Google Scholar] [CrossRef] [PubMed]
- Knirel, Y.A.; Gabius, H.J.; Blixt, O.; Rapoport, E.M.; Khasbiullina, N.R.; Shilova, N.V.; Bovin, N.V. Human tandem-repeat-type galectins bind bacterial nonβGal polysaccharides. Glycoconj. J. 2014, 31, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Hadari, Y.R.; Paz, K.; Dekel, R.; Mestrovic, T.; Accili, D.; Zick, Y. Galectin-8. A new rat lectin, related to galectin-4. J. Biol. Chem. 1995, 270, 3447–3453. [Google Scholar] [PubMed]
- Hadari, Y.R.; Arbel-Goren, R.; Levy, Y.; Amsterdam, A.; Alon, R.; Zakut, R.; Zick, Y. Galectin-8 binding to integrins inhibits cell adhesion and induces apoptosis. J. Cell Sci. 2000, 113, 2385–2397. [Google Scholar] [PubMed]
- Levy, Y.; Arbel-Goren, R.; Hadari, Y.R.; Eshhar, S.; Ronen, D.; Elhanany, E.; Geiger, B.; Zick, Y. Galectin-8 functions as a matricellular modulator of cell adhesion. J. Biol. Chem. 2001, 276, 31285–31295. [Google Scholar] [CrossRef] [PubMed]
- Pal, Z.; Antal, P.; Srivastava, S.K.; Hullam, G.; Semsei, A.F.; Gal, J.; Svebis, M.; Soos, G.; Szalai, C.; Andre, S.; et al. Non-synonymous single nucleotide polymorphisms in genes for immunoregulatory galectins: Association of galectin-8 (F19Y) occurrence with autoimmune diseases in a Caucasian population. Biochim. Biophys. Acta 2012, 1820, 1512–1518. [Google Scholar] [CrossRef] [PubMed]
- Gopalkrishnan, R.V.; Roberts, T.; Tuli, S.; Kang, D.; Christiansen, K.A.; Fisher, P.B. Molecular characterization of prostate carcinoma tumor antigen-1, PCTA-1, a human galectin-8 related gene. Oncogene 2000, 19, 4405–4416. [Google Scholar] [CrossRef] [PubMed]
- Bidon-Wagner, N.; le Pennec, J.P. Human galectin-8 isoforms and cancer. Glycoconj. J. 2004, 19, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Elola, M.T.; Ferragut, F.; Cardenas Delgado, V.M.; Nugnes, L.G.; Gentilini, L.; Laderach, D.; Troncoso, M.F.; Compagno, D.; Wolfenstein-Todel, C.; Rabinovich, G.A. Expression, localization and function of galectin-8, a tandem-repeat lectin, in human tumors. Histol. Histopathol. 2014, 29, 1093–1105. [Google Scholar] [PubMed]
- Levy, Y.; Auslender, S.; Eisenstein, M.; Vidavski, R.R.; Ronen, D.; Bershadsky, A.D.; Zick, Y. It depends on the hinge: A structure-functional analysis of galectin-8, a tandem-repeat type lectin. Glycobiology 2006, 16, 463–476. [Google Scholar] [CrossRef] [PubMed]
- Nishi, N.; Itoh, A.; Fujiyama, A.; Yoshida, N.; Araya, S.; Hirashima, M.; Shoji, H.; Nakamura, T. Development of highly stable galectins: Truncation of the linker peptide confers protease-resistance on tandem-repeat type galectins. FEBS Lett. 2005, 579, 2058–2064. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Nishi, N.; Nakakita, S.; Kamitori, S. Crystallization and preliminary X-ray diffraction analysis of a protease-resistant mutant form of human galectin-8. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2009, 65, 512–514. [Google Scholar] [CrossRef] [PubMed]
- Hirabayashi, J.; Hashidate, T.; Arata, Y.; Nishi, N.; Nakamura, T.; Hirashima, M.; Urashima, T.; Oka, T.; Futai, M.; Muller, W.E.; et al. Oligosaccharide specificity of galectins: A search by frontal affinity chromatography. Biochim. Biophys. Acta 2002, 1572, 232–254. [Google Scholar] [CrossRef]
- Ideo, H.; Seko, A.; Ishizuka, I.; Yamashita, K. The N-terminal carbohydrate recognition domain of galectin-8 recognizes specific glycosphingolipids with high affinity. Glycobiology 2003, 13, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Ideo, H.; Matsuzaka, T.; Nonaka, T.; Seko, A.; Yamashita, K. Galectin-8-N-domain recognition mechanism for sialylated and sulfated glycans. J. Biol. Chem. 2011, 286, 11346–11355. [Google Scholar] [CrossRef] [PubMed]
- Collins, P.M.; Hidari, K.I.; Blanchard, H. Slow diffusion of lactose out of galectin-3 crystals monitored by X-ray crystallography: Possible implications for ligand-exchange protocols. Acta Cryst. Sect. D Biol. Cryst. 2007, 63, 415–419. [Google Scholar] [CrossRef] [PubMed]
- Saraboji, K.; Hakansson, M.; Genheden, S.; Diehl, C.; Qvist, J.; Weininger, U.; Nilsson, U.J.; Leffler, H.; Ryde, U.; Akke, M.; et al. The Carbohydrate-binding site in galectin-3 is preorganized to recognize a sugarlike framework of oxygens: Ultra-high-resolution structures and water dynamics. Biochemistry 2012, 51, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Bum-Erdene, K.; Leffler, H.; Nilsson, U.J.; Blanchard, H. Structural characterisation of human galectin-4 N-terminal carbohydrate recognition domain in complex with glycerol, lactose, 3′-sulfo-lactose, and 2′-fucosyllactose. Sci. Rep. 2016, 6, 20289. [Google Scholar] [CrossRef] [PubMed]
- Jeong, K.H.; Byun, B.J.; Kang, Y.K. Conformational preferences of glycerol in the gas phase and in water. Bull. Korean Chem. Soc. 2012, 33, 917–924. [Google Scholar] [CrossRef]
- Umemoto, K.; Leffler, H.; Venot, A.; Valafar, H.; Prestegard, J.H. Conformational differences in liganded and unliganded states of Galectin-3. Biochemistry 2003, 42, 3688–3695. [Google Scholar] [CrossRef] [PubMed]
- Diehl, C.; Engstrom, O.; Delaine, T.; Hakansson, M.; Genheden, S.; Modig, K.; Leffler, H.; Ryde, U.; Nilsson, U.J.; Akke, M. Protein flexibility and conformational entropy in ligand design targeting the carbohydrate recognition domain of galectin-3. J. Am. Chem. Soc. 2010, 132, 14577–14589. [Google Scholar] [CrossRef] [PubMed]
- Stowell, S.R.; Arthur, C.M.; Slanina, K.A.; Horton, J.R.; Smith, D.F.; Cummings, R.D. Dimeric Galectin-8 induces phosphatidylserine exposure in leukocytes through polylactosamine recognition by the C-terminal domain. J. Biol. Chem. 2008, 283, 20547–20559. [Google Scholar] [CrossRef] [PubMed]
- Seetharaman, J.; Kanigsberg, A.; Slaaby, R.; Leffler, H.; Barondes, S.H.; Rini, J.M. X-ray crystal structure of the human galectin-3 carbohydrate recognition domain at 2.1-A resolution. J. Biol. Chem. 1998, 273, 13047–13052. [Google Scholar] [CrossRef] [PubMed]
- Sorme, P.; Arnoux, P.; Kahl-Knutsson, B.; Leffler, H.; Rini, J.M.; Nilsson, U.J. Structural and thermodynamic studies on cation-Pi interactions in lectin-ligand complexes: High-affinity galectin-3 inhibitors through fine-tuning of an arginine-arene interaction. J. Am. Chem. Soc. 2005, 127, 1737–1743. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, F.M.; Scholz, B.A.; Buzamet, E.; Kopitz, J.; Andre, S.; Menendez, M.; Romero, A.; Solis, D.; Gabius, H.J. Natural single amino acid polymorphism (F19Y) in human galectin-8: Detection of structural alterations and increased growth-regulatory activity on tumor cells. FEBS J. 2014, 281, 1446–1464. [Google Scholar] [CrossRef] [PubMed]
- Pinard, M.A.; Kurian, J.J.; Aggarwal, M.; Agbandje-McKenna, M.; McKenna, R. Cryoannealing-induced space-group transition of crystals of the carbonic anhydrase psCA3. Acta Cryst. Sect. F Struct. Biol. Commun. 2016, 72, 573–577. [Google Scholar] [CrossRef] [PubMed]
- John, S.; Mishra, R. Galectin-9: From cell biology to complex disease dynamics. J. Biosci. 2016, 41, 507–534. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, Y.; Ogawa, T.; Oomizu, S.; Nakakita, S.; Nishi, N.; Kamitori, S.; Hirashima, M.; Nakamura, T. Self-association of the galectin-9 C-terminal domain via the opposite surface of the sugar-binding site. J. Biochem. 2013, 153, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Nishi, N.; Itoh, A.; Shoji, H.; Miyanaka, H.; Nakamura, T. Galectin-8 and galectin-9 are novel substrates for thrombin. Glycobiology 2006, 16, 15C–20C. [Google Scholar] [CrossRef] [PubMed]
- Gallwitz, M.; Enoksson, M.; Thorpe, M.; Hellman, L. The extended cleavage specificity of human thrombin. PLoS ONE 2012, 7, e31756. [Google Scholar] [CrossRef] [PubMed]
- Zick, Y.; Eisenstein, M.; Goren, R.A.; Hadari, Y.R.; Levy, Y.; Ronen, D. Role of galectin-8 as a modulator of cell adhesion and cell growth. Glycoconj. J. 2004, 19, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, S.; Oberg, C.T.; Carlsson, M.C.; Sundin, A.; Nilsson, U.J.; Smith, D.; Cummings, R.D.; Almkvist, J.; Karlsson, A.; Leffler, H. Affinity of galectin-8 and its carbohydrate recognition domains for ligands in solution and at the cell surface. Glycobiology 2007, 17, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Si, Y.; Feng, S.; Gao, J.; Wang, Y.; Zhang, Z.; Meng, Y.; Zhou, Y.; Tai, G.; Su, J. Human galectin-2 interacts with carbohydrates and peptides non-classically: New insight from X-ray crystallography and hemagglutination. Acta Biochim. Biophys. Sin. 2016, 48, 939–947. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Zhang, T.; Wang, P.; Liu, F.; Tai, G.; Zhou, Y. The water network in galectin-3 ligand binding site guides inhibitor design. Acta Biochim. Biophys. Sin. 2015, 47, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Kabsch, W. XDS. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Evans, P.R.; Murshudov, G.N. How good are my data and what is the resolution? Acta Crystallogr. D Biol. Crystallogr. 2013, 69, 1204–1214. [Google Scholar] [CrossRef] [PubMed]
- Potterton, E.; Briggs, P.; Turkenburg, M.; Dodson, E. A graphical user interface to the CCP4 program suite. Acta Crystallogr. D Biol. Crystallogr. 2003, 59, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser crystallographic software. J. Appl. Crystallogr. 2007, 40, 658–674. [Google Scholar] [CrossRef] [PubMed]
- Adams, P.D.; Afonine, P.V.; Bunkoczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.W.; Kapral, G.J.; Grosse-Kunstleve, R.W. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Chen, V.B.; Arendall, W.B., 3rd; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. MolProbity: All-atom structure validation for macromolecular crystallography. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Davis, I.W.; Leaver-Fay, A.; Chen, V.B.; Block, J.N.; Kapral, G.J.; Wang, X.; Murray, L.W.; Arendall, W.B., 3rd; Snoeyink, J.; Richardson, J.S.; et al. MolProbity: All-atom contacts and structure validation for proteins and nucleic acids. Nucleic Acids Res. 2007, 35, 375–383. [Google Scholar] [CrossRef] [PubMed]
- The PyMOL Molecular Graphics System, Version 1.7.2. Schrödinger LLC, 2015. Available online: www.pymol.org (accessed on 25 April 2016).
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Zhi, Y.; Sun, L.; Peng, X.; Zhang, T.; Xue, H.; Tai, G.; Zhou, Y. The inhibitory effects of a rhamnogalacturonan I (RG-I) domain from ginseng pectin on galectin-3 and its structure-activity relationship. J. Biol. Chem. 2013, 288, 33953–33965. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Structure Name | Structure 1 | Structure 2 | Structure 3 | Structure 4 | Structure 5 |
|---|---|---|---|---|---|
| PDB code | 5GZC | 5GZD | 5GZE | 5GZF | 5GZG |
| Resolution (Å) | 20.00–1.08 (1.10–1.08) | 20.00–1.19 (1.21–1.19) | 20.00–1.32 (1.35–1.32) | 20.00–2.00 (2.05–2.00) | 20.00–2.00 (2.03–2.00) |
| Space group | P212121 | C2 | C2 | P43212 | P43212 |
| Unit cell parameters (α, β, γ, a, b, c) (Å) | 90, 90, 90, 40.56, 49.38, 76.68 | 90, 108.93, 90, 60.53, 40.05, 59.18 | 90, 108.02, 90, 60.69, 39.74, 55.80 | 90, 90, 90, 48.87, 48.87, 159.73 | 90, 90, 90, 49.13, 49.13, 159.67 |
| no. of measured reflections | 776,761 (19,377) | 258,417 (10,225) | 187,310 (5330) | 847,50 (6299) | 851,78 (5364) |
| no. of unique reflections | 65,391 (3047) | 40,642 (1864) | 29,391 (1205) | 13,850 (1003) | 14,014 (978) |
| Completeness (%) | 98.0 (93.1) | 99.3 (89.9) | 98.2 (80.2) | 99.7 (98.4) | 99.3 (95.1) |
| multiplicity | 11.9 (6.4) | 6.4 (5.5) | 6.4 (4.4) | 6.1 (6.3) | 6.1 (5.5) |
| Rmerge (%) b | 9.9 (72.7) | 4.6 (31.8) | 7.2 (49.2) | 9.8 (22.5) | 5.6 (70.7) |
| <I/∑(I)> | 10.9 (2.4) | 17.5 (4.2) | 13.4 (2.1) | 13.3 (6.7) | 14.9 (1.9) |
| Refine | |||||
| Resolution limits (Å) | 20.00–1.08 | 20.00–1.19 | 20.00–1.32 | 20.00–2.00 | 20.00–2.00 |
| Rmodel (%) c | 15.67 | 20.55 | 25.22 | 19.80 | 20.27 |
| Rfree (%) d | 16.27 | 21.39 | 28.08 | 21.52 | 25.80 |
| Rmsd bond lengths (Å) | 0.008 | 0.01 | 0.011 | 0.008 | 0.007 |
| Rmsd bond angles (°) | 1.047 | 1.398 | 1.422 | 0.993 | 1.062 |
| Ramachandran plot e residue in favored regions (%) | 99.3% | 98.6% | 95.9% | 97.3% | 96.6% |
| Ramachandran outliers (%) | 1.5 | 0 | 0 | 0.8 | 0.8 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Si, Y.; Wang, Y.; Gao, J.; Song, C.; Feng, S.; Zhou, Y.; Tai, G.; Su, J. Crystallization of Galectin-8 Linker Reveals Intricate Relationship between the N-terminal Tail and the Linker. Int. J. Mol. Sci. 2016, 17, 2088. https://doi.org/10.3390/ijms17122088
Si Y, Wang Y, Gao J, Song C, Feng S, Zhou Y, Tai G, Su J. Crystallization of Galectin-8 Linker Reveals Intricate Relationship between the N-terminal Tail and the Linker. International Journal of Molecular Sciences. 2016; 17(12):2088. https://doi.org/10.3390/ijms17122088
Chicago/Turabian StyleSi, Yunlong, Yue Wang, Jin Gao, Chenyang Song, Shiqiong Feng, Yifa Zhou, Guihua Tai, and Jiyong Su. 2016. "Crystallization of Galectin-8 Linker Reveals Intricate Relationship between the N-terminal Tail and the Linker" International Journal of Molecular Sciences 17, no. 12: 2088. https://doi.org/10.3390/ijms17122088
APA StyleSi, Y., Wang, Y., Gao, J., Song, C., Feng, S., Zhou, Y., Tai, G., & Su, J. (2016). Crystallization of Galectin-8 Linker Reveals Intricate Relationship between the N-terminal Tail and the Linker. International Journal of Molecular Sciences, 17(12), 2088. https://doi.org/10.3390/ijms17122088