
Testosterone Deficiency Induces Changes of the Transcriptomes of Visceral Adipose Tissue in Miniature Pigs Fed a High-Fat and High-Cholesterol Diet


Abstract
:1. Introduction
2. Results
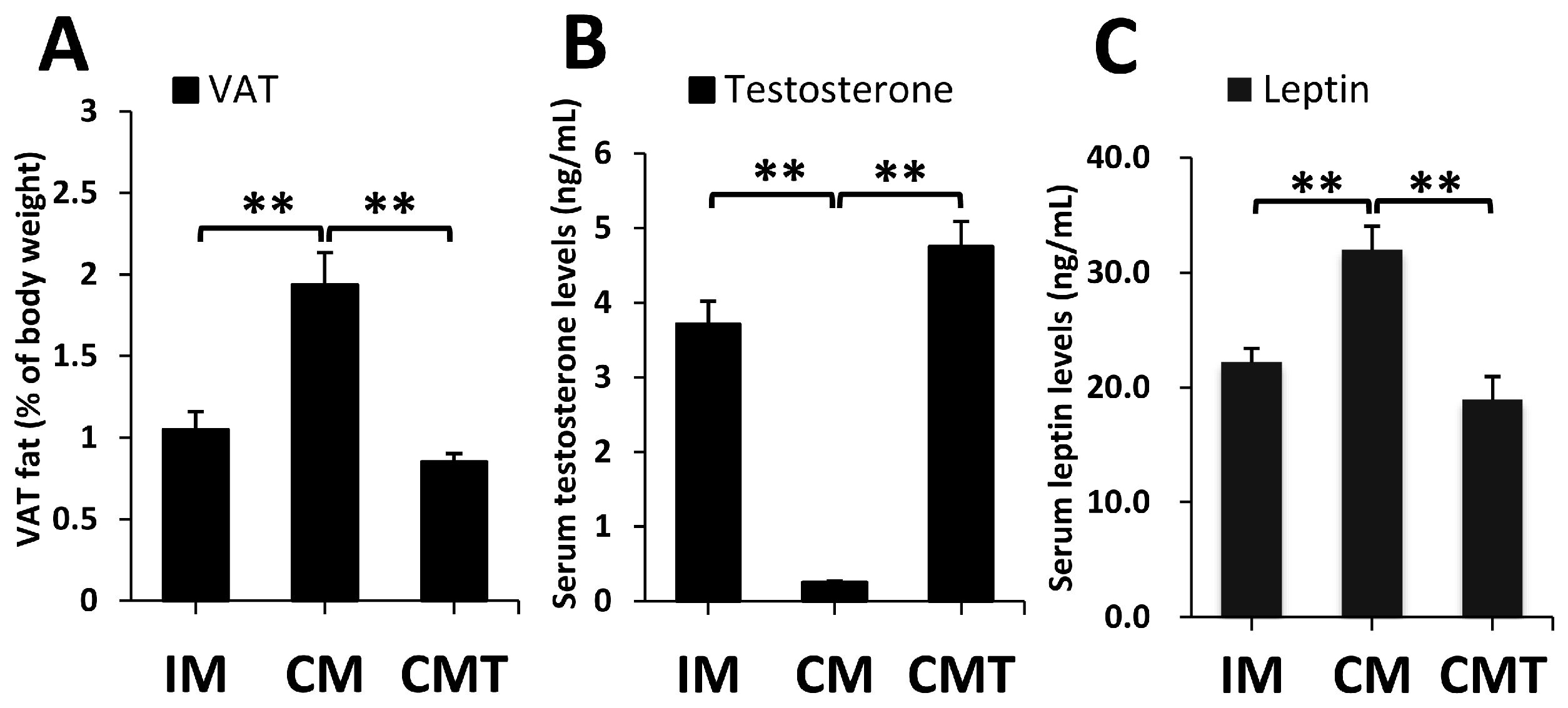
2.1. Effects of Castration and Testosterone Treatment on VAT Fat and Serum Leptin Levels
2.2. Characteristics of RNA-Seq
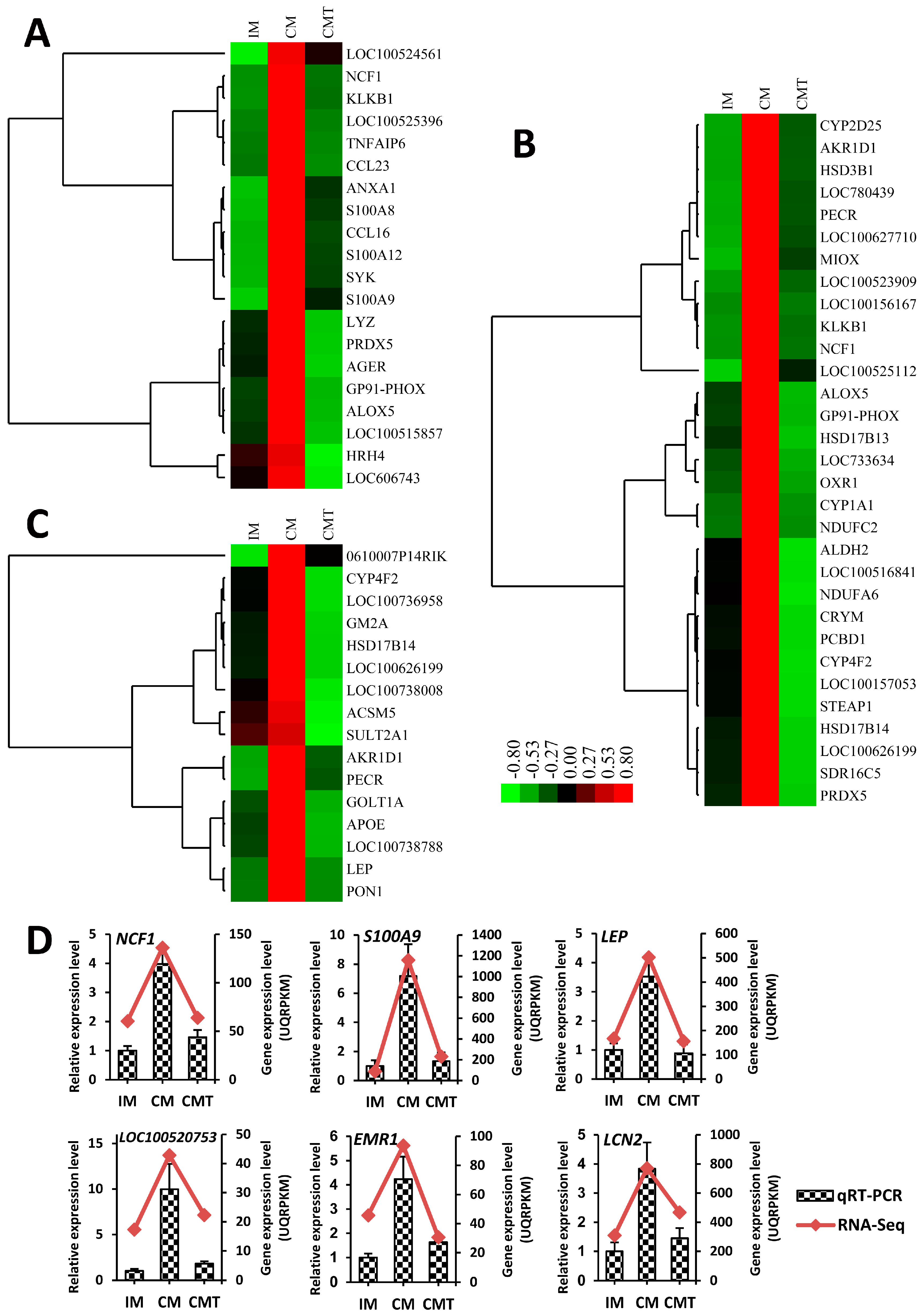
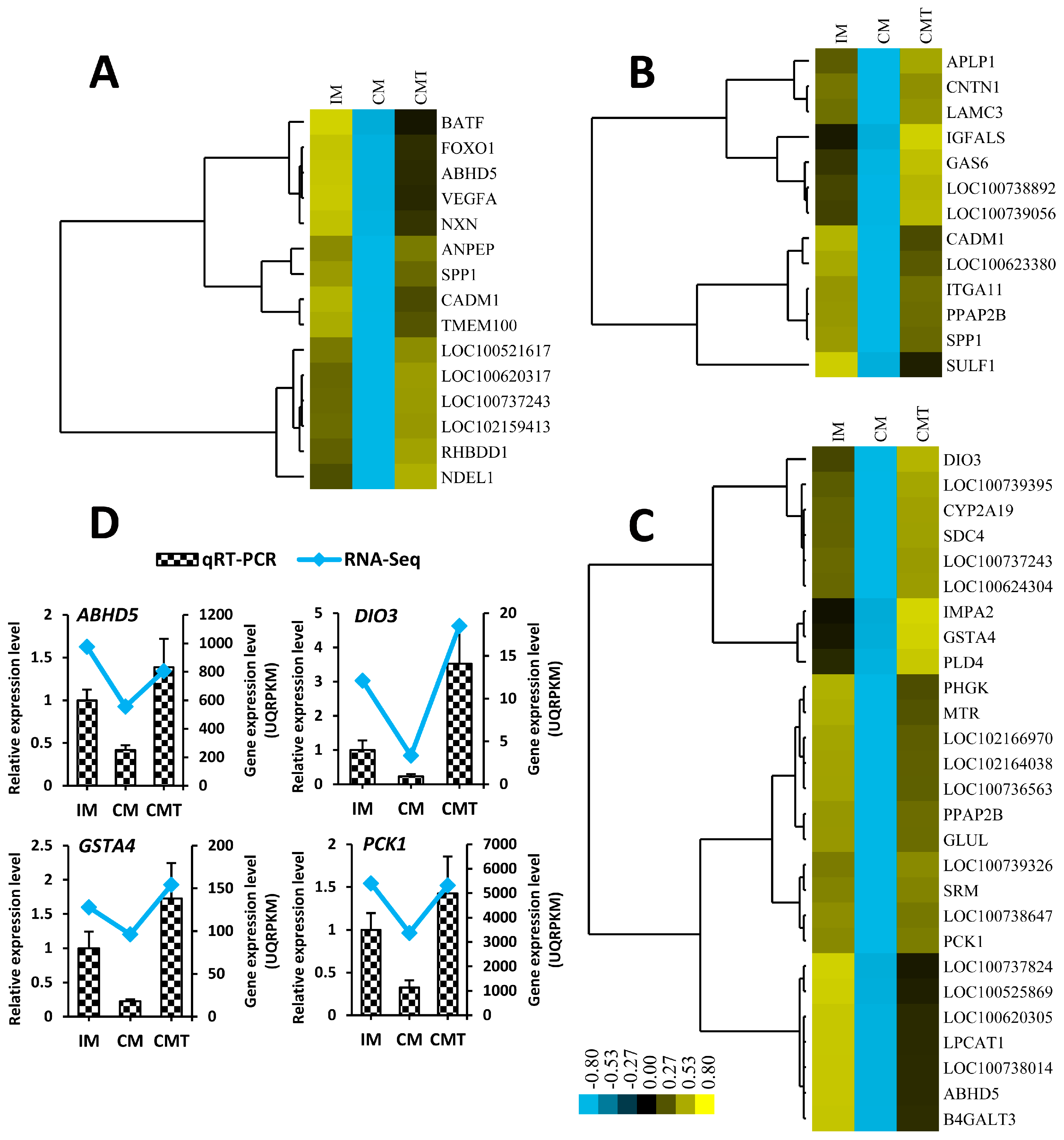
2.3. Gene Expression Analysis
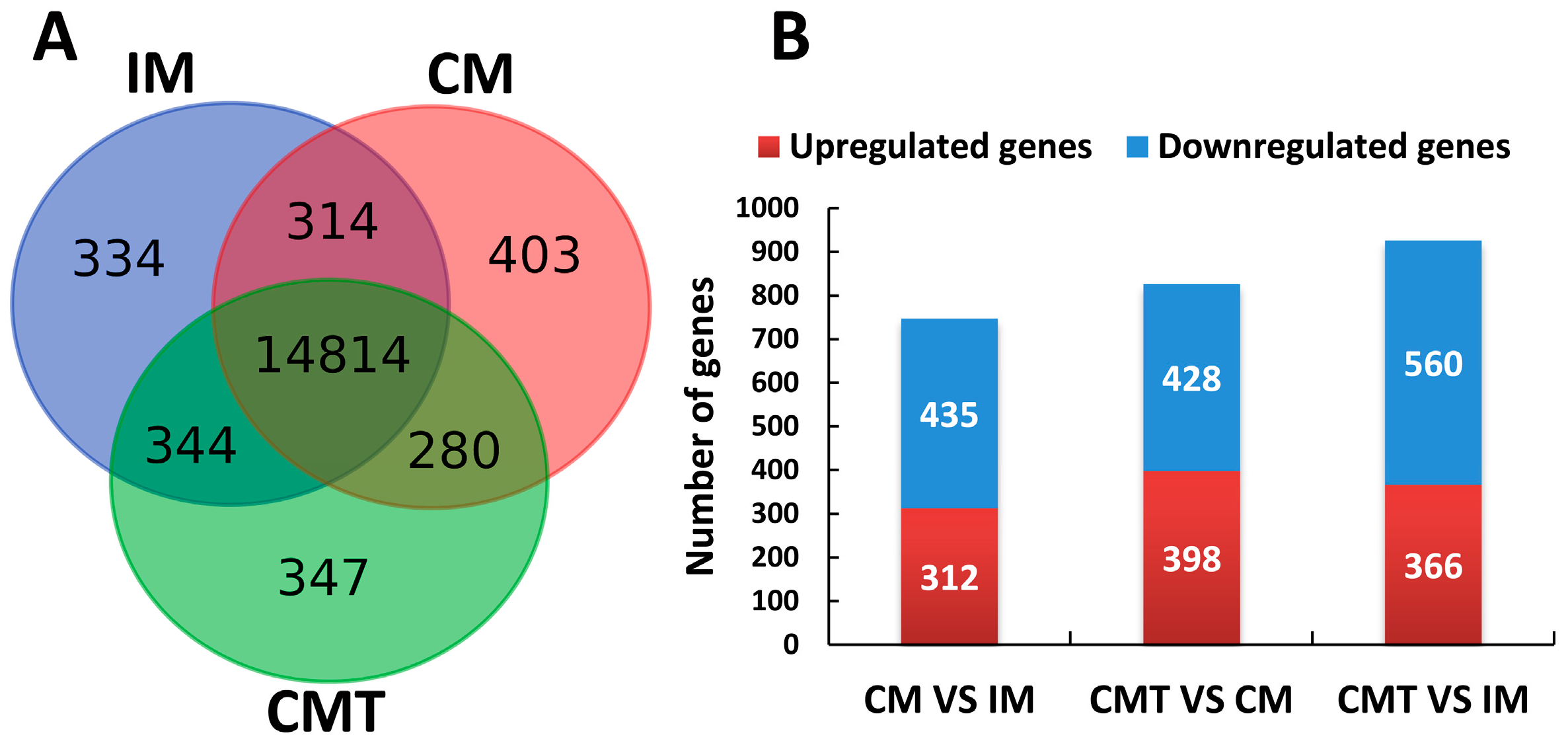
2.4. Identification of Differentially Expressed Genes (DEGs)
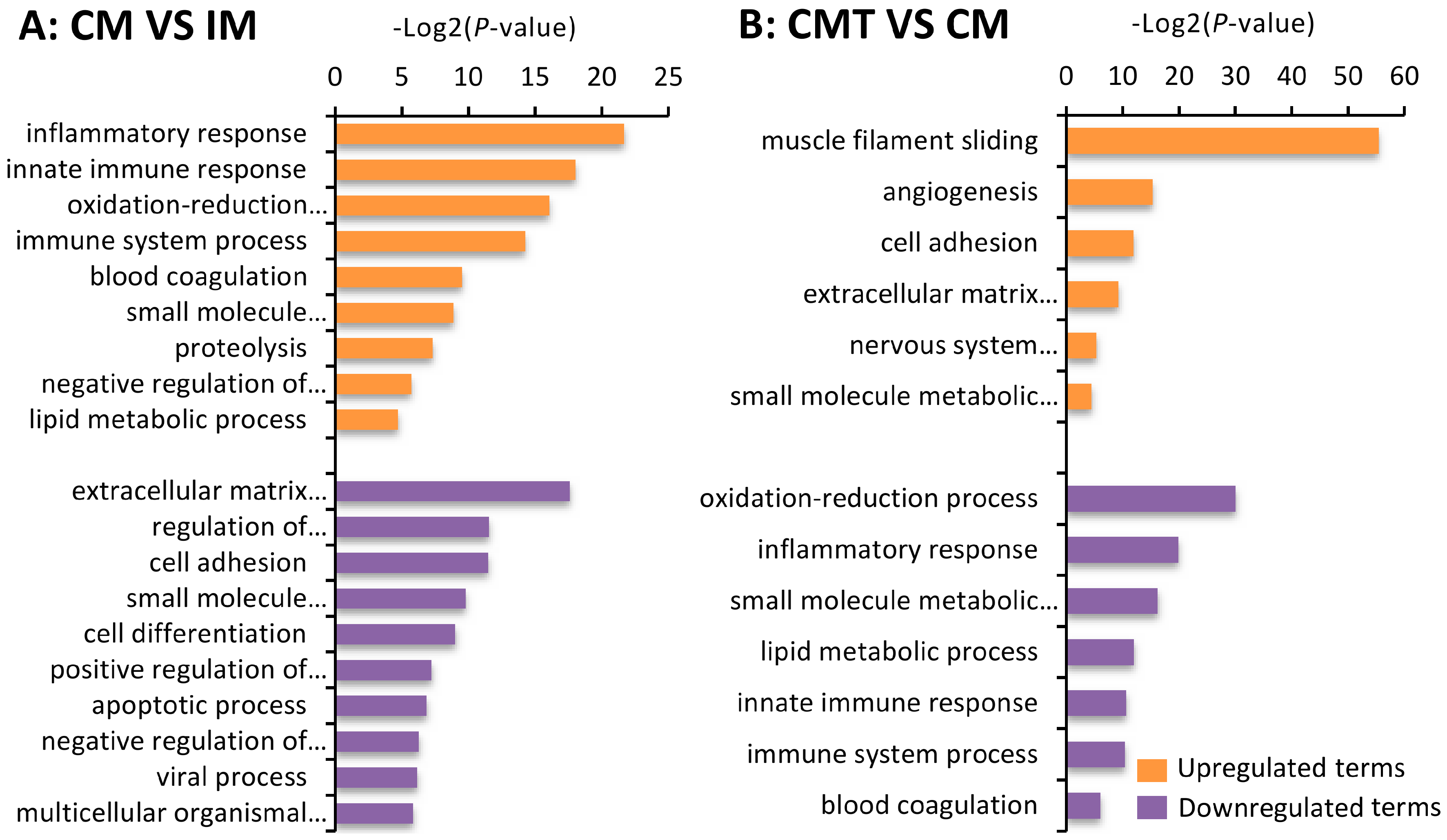
2.5. Gene Ontology (GO) Enrichment Analysis of DEGs between Different Groups
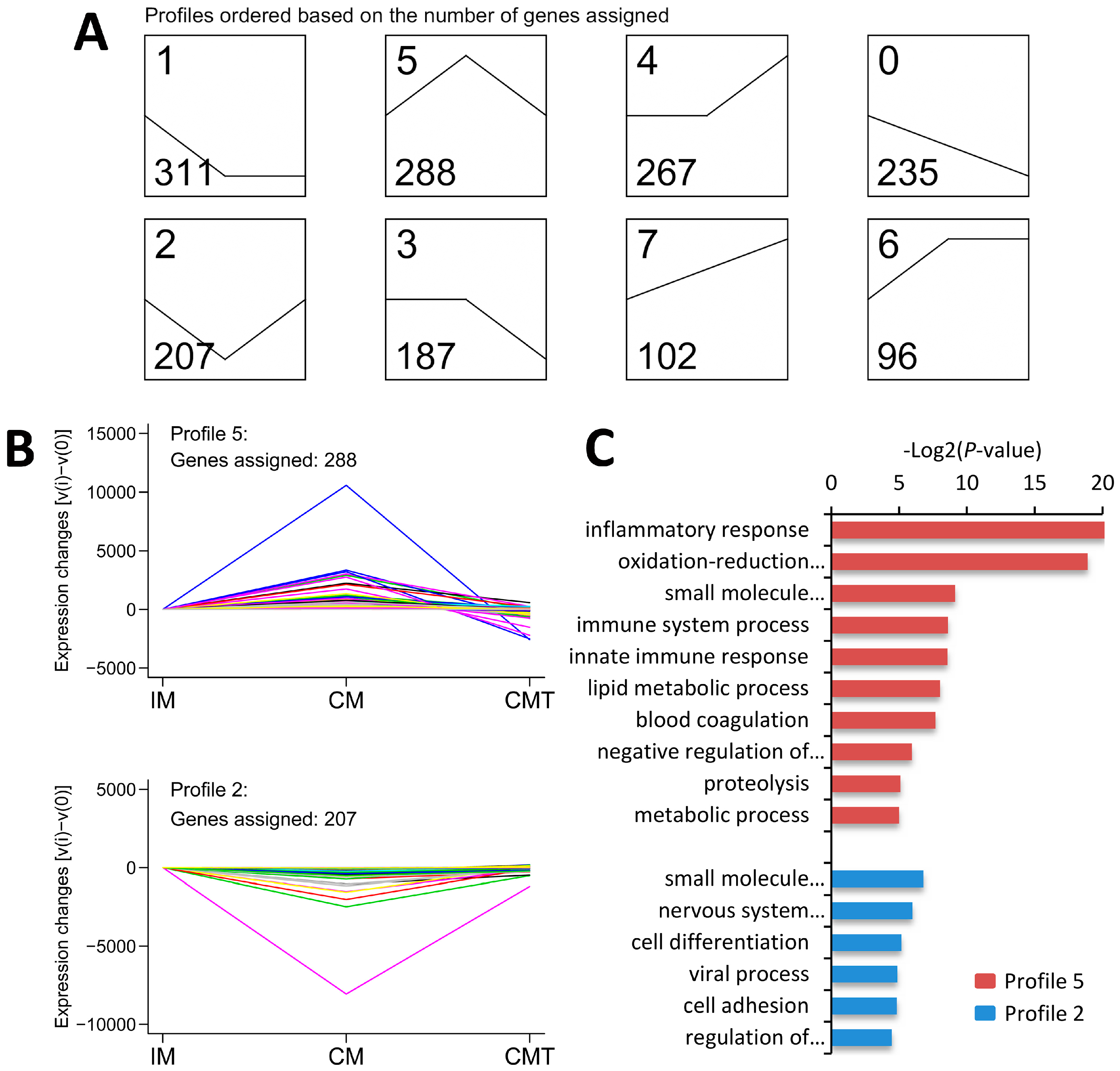
2.6. Series-Cluster Analysis of DEGs
2.7. GO Enrichment Analysis of DEGs Involved in Profile 5 and 2
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. RNA Extraction, cDNA Library Construction and Ion Proton Sequencing
4.3. Sequence Mapping and DEGs Filtering
4.4. Cluster Analysis
4.5. Gene Ontology (GO) Analysis
4.6. Quantitative Real-Time RT-PCR
4.7. Statistical Analysis
4.8. Data Accessibility
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zheng, H.; Zhang, C.; Yang, W.; Wang, Y.; Lin, Y.; Yang, P.; Yu, Q.; Fan, J.; Liu, E. Fat and cholesterol diet induced lipid metabolic disorders and insulin resistance in rabbit. Exp. Clin. Endocrinol. Diabetes 2009, 117, 400–405. [Google Scholar] [CrossRef] [PubMed]
- Muir, R.; Ballan, J.; Clifford, B.; McMullen, S.; Khan, R.; Shmygol, A.; Quenby, S.; Elmes, M. Modelling maternal obesity: The effects of a chronic high-fat, high-cholesterol diet on uterine expression of contractile-associated proteins and ex vivo contractile activity during labour in the rat. Clin. Sci. 2016, 130, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, D.; Aldred, H.; Clark, S.; Channer, K.S.; Jones, T.H. Clinical and biochemical assessment of hypogonadism in men with type 2 diabetes: Correlations with bioavailable testosterone and visceral adiposity. Diabetes Care 2007, 30, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Tsai, E.C.; Boyko, E.J.; Leonetti, D.L.; Fujimoto, W.Y. Low serum testosterone level as a predictor of increased visceral fat in Japanese–American men. Int. J. Obes. Relat. Metab. Disord. 2000, 24, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Saad, F. Androgen therapy in men with testosterone deficiency: Can testosterone reduce the risk of cardiovascular disease? Diabetes Metab. Res. Rev. 2012, 28 (Suppl. S2), 52–59. [Google Scholar] [CrossRef] [PubMed]
- Allan, C.A.; McLachlan, R.I. Androgens and obesity. Curr. Opin. Endocrinol. Diabetes Obes. 2010, 17, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Yassin, A.; Doros, G. Testosterone therapy in hypogonadal men results in sustained and clinically meaningful weight loss. Clin. Obes. 2013, 3, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, D.; Goodwin, E.; Channer, K.S.; Jones, T.H. Testosterone replacement therapy improves insulin resistance, glycaemic control, visceral adiposity and hypercholesterolaemia in hypogonadal men with type 2 diabetes. Eur. J. Endocrinol. 2006, 154, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Kalinchenko, S.Y.; Tishova, Y.A.; Mskhalaya, G.J.; Gooren, L.J.; Giltay, E.J.; Saad, F. Effects of testosterone supplementation on markers of the metabolic syndrome and inflammation in hypogonadal men with the metabolic syndrome: The double-blinded placebo-controlled moscow study. Clin. Endocrinol. 2010, 73, 602–612. [Google Scholar] [CrossRef] [PubMed]
- Traish, A.M.; Zitzmann, M. The complex and multifactorial relationship between testosterone deficiency (TD), obesity and vascular disease. Rev. Endocr. Metab. Disord. 2015, 16, 249–268. [Google Scholar] [CrossRef] [PubMed]
- Maneschi, E.; Morelli, A.; Filippi, S.; Cellai, I.; Comeglio, P.; Mazzanti, B.; Mello, T.; Calcagno, A.; Sarchielli, E.; Vignozzi, L.; et al. Testosterone treatment improves metabolic syndrome-induced adipose tissue derangements. J. Endocrinol. 2012, 215, 347–362. [Google Scholar] [CrossRef] [PubMed]
- Abdelhamed, A.; Hisasue, S.; Shirai, M.; Matsushita, K.; Wakumoto, Y.; Tsujimura, A.; Tsukamoto, T.; Horie, S. Testosterone replacement alters the cell size in visceral fat but not in subcutaneous fat in hypogonadal aged male rats as a late-onset hypogonadism animal model. Res. Rep. Urol. 2015, 7, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Saad, F.; Yassin, A.; Doros, G.; Haider, A. Effects of long-term treatment with testosterone on weight and waist size in 411 hypogonadal men with obesity classes I–III: Observational data from two registry studies. Int. J. Obes. 2016, 40, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Christoffersen, B.O.; Grand, N.; Golozoubova, V.; Svendsen, O.; Raun, K. Gender-associated differences in metabolic syndrome-related parameters in Göttingen minipigs. Comp. Med. 2007, 57, 493–504. [Google Scholar] [PubMed]
- Christoffersen, B.; Golozoubova, V.; Pacini, G.; Svendsen, O.; Raun, K. The young Göttingen minipig as a model of childhood and adolescent obesity: Influence of diet and gender. Obesity 2013, 21, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Koopmans, S.J.; Schuurman, T. Considerations on pig models for appetite, metabolic syndrome and obese type 2 diabetes: From food intake to metabolic disease. Eur. J. Pharmacol. 2015, 759, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Zhang, L.; Chen, M.; Jiang, X.; Xu, N. Castration-induced changes in microRNA expression profiles in subcutaneous adipose tissue of male pigs. J. Appl. Genet. 2014, 55, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Jiang, X.; Pan, Y.; Chen, L.; Zhang, L.; Zhu, K.; Cai, Y.; Ling, Y.; Chen, F.; Xu, X.; et al. Transcriptomic analysis of hepatic responses to testosterone deficiency in miniature pigs fed a high-cholesterol diet. BMC Genom. 2015, 16, 59. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Xi, H.; Pan, Y.; Jiang, X.; Chen, L.; Cai, Y.; Zhu, K.; Chen, C.; Xu, X.; Chen, M. Effect of testosterone deficiency on cholesterol metabolism in pigs fed a high-fat and high-cholesterol diet. Lipids Health Dis. 2015, 14, 18. [Google Scholar] [CrossRef] [PubMed]
- Harada, N.; Hanaoka, R.; Horiuchi, H.; Kitakaze, T.; Mitani, T.; Inui, H.; Yamaji, R. Castration influences intestinal microflora and induces abdominal obesity in high-fat diet-fed mice. Sci. Rep. 2016, 6, 23001. [Google Scholar] [CrossRef] [PubMed]
- Rubinow, K.B.; Wang, S.; den Hartigh, L.J.; Subramanian, S.; Morton, G.J.; Buaas, F.W.; Lamont, D.; Gray, N.; Braun, R.E.; Page, S.T. Hematopoietic androgen receptor deficiency promotes visceral fat deposition in male mice without impairing glucose homeostasis. Andrology 2015, 3, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Donner, D.G.; Elliott, G.E.; Beck, B.R.; Bulmer, A.C.; Du Toit, E.F. Impact of diet-induced obesity and testosterone deficiency on the cardiovascular system: A novel rodent model representative of males with testosterone-deficient metabolic syndrome (TDMetS). PLoS ONE 2015, 10, e0138019. [Google Scholar] [CrossRef] [PubMed]
- Kiess, W.; Reich, A.; Meyer, K.; Glasow, A.; Deutscher, J.; Klammt, J.; Yang, Y.; Muller, G.; Kratzsch, J. A role for leptin in sexual maturation and puberty? Horm. Res. 1999, 51 (Suppl. S3), 55–63. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.W.; Zhao, X.F.; Yao, Y.C.; Jiang, X.L.; Zhang, L.F.; Xu, N.Y.; Wu, C.X. Castration modulates leptin and its receptor in the hypothalamus from male pigs. J. Anim. Vet. Adv. 2010, 9, 1696–1701. [Google Scholar]
- Wabitsch, M.; Blum, W.F.; Muche, R.; Braun, M.; Hube, F.; Rascher, W.; Heinze, E.; Teller, W.; Hauner, H. Contribution of androgens to the gender difference in leptin production in obese children and adolescents. J. Clin. Investig. 1997, 100, 808–813. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, D.; Clarke, S.; Stanworth, R.; Channer, K.S.; Jones, T.H. The effect of testosterone replacement therapy on adipocytokines and C-reactive protein in hypogonadal men with type 2 diabetes. Eur. J. Endocrinol. 2007, 156, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, S.M.; Perez-Tilve, D.; Greer, T.M.; Coburn, B.A.; Grant, E.; Basford, J.E.; Tschop, M.H.; Hui, D.Y. Defective lipid delivery modulates glucose tolerance and metabolic response to diet in apolipoprotein E-deficient mice. Diabetes 2008, 57, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Perrard, X.D.; Perrard, J.L.; Mukherjee, A.; Rosales, C.; Chen, Y.; Smith, C.W.; Pownall, H.J.; Ballantyne, C.M.; Wu, H. ApoE and the role of very low density lipoproteins in adipose tissue inflammation. Atherosclerosis 2012, 223, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.F.; Jiang, X.L.; Hua, X.C.; Lu, Y.P.; Xu, N.Y. Molecular cloning, expression and polymorphism of the porcine apolipoprotein A5 gene in a Jinhua × Pietrain F2 reference population. Animal 2010, 4, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Timpson, N.J.; Walter, K.; Min, J.L.; Tachmazidou, I.; Malerba, G.; Shin, S.Y.; Chen, L.; Futema, M.; Southam, L.; Iotchkova, V.; et al. A rare variant in APOC3 is associated with plasma triglyceride and VLDL levels in Europeans. Nat. Commun. 2014, 5, 4871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drosatos, K.; Sanoudou, D.; Kypreos, K.E.; Kardassis, D.; Zannis, V.I. A dominant negative form of the transcription factor c-jun affects genes that have opposing effects on lipid homeostasis in mice. J. Biol. Chem. 2007, 282, 19556–19564. [Google Scholar] [CrossRef] [PubMed]
- Dallinga-Thie, G.M.; van Tol, A.; Hattori, H.; van Vark-van der Zee, L.C.; Jansen, H.; Sijbrands, E.J. Plasma apolipoprotein A5 and triglycerides in type 2 diabetes. Diabetologia 2006, 49, 1505–1511. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.J.; Bao, S.; Bolin, E.R.; Burris, D.L.; Xu, X.; Sun, Q.; Killilea, D.W.; Shen, Q.; Ziouzenkova, O.; Belury, M.A.; et al. Zinc deficiency augments leptin production and exacerbates macrophage infiltration into adipose tissue in mice fed a high-fat diet. J. Nutr. 2013, 143, 1036–1045. [Google Scholar] [CrossRef] [PubMed]
- Michaud, A.; Drolet, R.; Noel, S.; Paris, G.; Tchernof, A. Visceral fat accumulation is an indicator of adipose tissue macrophage infiltration in women. Metabolism 2012, 61, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W., Jr. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Yu, S.; Lambert, J.D. Dietary cocoa ameliorates obesity-related inflammation in high fat-fed mice. Eur. J. Nutr. 2014, 53, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wu, Y.; Zhang, Y.; Leroith, D.; Bernlohr, D.A.; Chen, X. The role of lipocalin 2 in the regulation of inflammation in adipocytes and macrophages. Mol. Endocrinol. 2008, 22, 1416–1426. [Google Scholar] [CrossRef] [PubMed]
- Catalan, V.; Gomez-Ambrosi, J.; Rodriguez, A.; Ramirez, B.; Silva, C.; Rotellar, F.; Gil, M.J.; Cienfuegos, J.A.; Salvador, J.; Fruhbeck, G. Increased adipose tissue expression of lipocalin-2 in obesity is related to inflammation and matrix metalloproteinase-2 and metalloproteinase-9 activities in humans. J. Mol. Med. 2009, 87, 803–813. [Google Scholar] [CrossRef] [PubMed]
- Sekimoto, R.; Kishida, K.; Nakatsuji, H.; Nakagawa, T.; Funahashi, T.; Shimomura, I. High circulating levels of S100A8/A9 complex (calprotectin) in male Japanese with abdominal adiposity and dysregulated expression of S100A8 and S100A9 in adipose tissues of obese mice. Biochem. Biophys. Res. Commun. 2012, 419, 782–789. [Google Scholar] [CrossRef] [PubMed]
- Yamaoka, M.; Maeda, N.; Nakamura, S.; Mori, T.; Inoue, K.; Matsuda, K.; Sekimoto, R.; Kashine, S.; Nakagawa, Y.; Tsushima, Y.; et al. Gene expression levels of S100 protein family in blood cells are associated with insulin resistance and inflammation (peripheral blood S100 mRNAs and metabolic syndrome). Biochem. Biophys. Res. Commun. 2013, 433, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Sekimoto, R.; Fukuda, S.; Maeda, N.; Tsushima, Y.; Matsuda, K.; Mori, T.; Nakatsuji, H.; Nishizawa, H.; Kishida, K.; Kikuta, J.; et al. Visualized macrophage dynamics and significance of S100A8 in obese fat. Proc. Natl. Acad. Sci. USA 2015, 112, E2058–E2066. [Google Scholar] [CrossRef] [PubMed]
- Gletsu-Miller, N.; Hansen, J.M.; Jones, D.P.; Go, Y.M.; Torres, W.E.; Ziegler, T.R.; Lin, E. Loss of total and visceral adipose tissue mass predicts decreases in oxidative stress after weight-loss surgery. Obesity 2009, 17, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Wainright, K.S.; Fleming, N.J.; Rowles, J.L.; Welly, R.J.; Zidon, T.M.; Park, Y.M.; Gaines, T.L.; Scroggins, R.J.; Anderson-Baucum, E.K.; Hasty, A.H.; et al. Retention of sedentary obese visceral white adipose tissue phenotype with intermittent physical activity despite reduced adiposity. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 309, R594–R602. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, K.; Puri, N.; Kim, D.H.; Hinds, T.D.; Stechschulte, L.A.; Favero, G.; Rodella, L.; Shapiro, J.I.; Jude, D.; Abraham, N.G. PPARδ binding to heme oxygenase 1 promoter prevents angiotensin II-induced adipocyte dysfunction in Goldblatt hypertensive rats. Int. J. Obes. 2014, 38, 456–465. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Kim, A.Y.; Choi, J.W.; Kim, M.; Yasue, S.; Son, H.J.; Masuzaki, H.; Park, K.S.; Kim, J.B. Dysregulation of adipose glutathione peroxidase 3 in obesity contributes to local and systemic oxidative stress. Mol. Endocrinol. 2008, 22, 2176–2189. [Google Scholar] [CrossRef] [PubMed]
- DeFuria, J.; Bennett, G.; Strissel, K.J.; Perfield, J.W., 2nd; Milbury, P.E.; Greenberg, A.S.; Obin, M.S. Dietary blueberry attenuates whole-body insulin resistance in high fat-fed mice by reducing adipocyte death and its inflammatory sequelae. J. Nutr. 2009, 139, 1510–1516. [Google Scholar] [CrossRef] [PubMed]
- Curtis, J.M.; Grimsrud, P.A.; Wright, W.S.; Xu, X.; Foncea, R.E.; Graham, D.W.; Brestoff, J.R.; Wiczer, B.M.; Ilkayeva, O.; Cianflone, K.; et al. Downregulation of adipose glutathione S-transferase A4 leads to increased protein carbonylation, oxidative stress, and mitochondrial dysfunction. Diabetes 2010, 59, 1132–1142. [Google Scholar] [CrossRef] [PubMed]
- Lass, A.; Zimmermann, R.; Haemmerle, G.; Riederer, M.; Schoiswohl, G.; Schweiger, M.; Kienesberger, P.; Strauss, J.G.; Gorkiewicz, G.; Zechner, R. Adipose triglyceride lipase-mediated lipolysis of cellular fat stores is activated by CGI-58 and defective in Chanarin-Dorfman Syndrome. Cell Metab. 2006, 3, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Omatsu, N.; Morimoto, E.; Nakashima, H.; Ueno, K.; Tanaka, T.; Satouchi, K.; Hirose, F.; Osumi, T. CGI-58 facilitates lipolysis on lipid droplets but is not involved in the vesiculation of lipid droplets caused by hormonal stimulation. J. Lipid Res. 2007, 48, 1078–1089. [Google Scholar] [CrossRef] [PubMed]
- Miao, H.; Ou, J.; Ma, Y.; Guo, F.; Yang, Z.; Wiggins, M.; Liu, C.; Song, W.; Han, X.; Wang, M.; et al. Macrophage CGI-58 deficiency activates ROS-inflammasome pathway to promote insulin resistance in mice. Cell Rep. 2014, 7, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.M.; Betters, J.L.; Lord, C.; Ma, Y.; Han, X.; Yang, K.; Alger, H.M.; Melchior, J.; Sawyer, J.; Shah, R.; et al. CGI-58 knockdown in mice causes hepatic steatosis but prevents diet-induced obesity and glucose intolerance. J. Lipid Res. 2010, 51, 3306–3315. [Google Scholar] [CrossRef] [PubMed]
- Erikson, D.W.; Burghardt, R.C.; Bayless, K.J.; Johnson, G.A. Secreted phosphoprotein 1 (SPP1, osteopontin) binds to integrin alphavbeta6 on porcine trophectoderm cells and integrin alphavbeta3 on uterine luminal epithelial cells, and promotes trophectoderm cell adhesion and migration. Biol. Reprod. 2009, 81, 814–825. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.J.; Shen, X.D.; Wen, J.B.; Kong, Y.; Chu, J.J.; Yan, G.Q.; Li, T.; Liu, D.; Wu, M.Q.; Zeng, G.H.; et al. Osteopontin-induced brown adipogenesis from white preadipocytes through a PI3K-AKT dependent signaling. Biochem. Biophys. Res. Commun. 2015, 459, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Luedtke, C.C.; McKee, M.D.; Cyr, D.G.; Gregory, M.; Kaartinen, M.T.; Mui, J.; Hermo, L. Osteopontin expression and regulation in the testis, efferent ducts, and epididymis of rats during postnatal development through to adulthood. Biol. Reprod. 2002, 66, 1437–1448. [Google Scholar] [CrossRef] [PubMed]
- Bertin, F.R.; Lemarié, C.A.; Robins, R.S.; Blostein, M.D. Growth arrest-specific 6 regulates thrombin-induced expression of vascular cell adhesion molecule-1 through forkhead box O1 in endothelial cells. J. Thromb. Haemost. 2015, 13, 2260–2272. [Google Scholar] [CrossRef] [PubMed]
- Ammoun, S.; Provenzano, L.; Zhou, L.; Barczyk, M.; Evans, K.; Hilton, D.A.; Hafizi, S.; Hanemann, C.O. Axl/Gas6/NFκB signalling in schwannoma pathological proliferation, adhesion and survival. Oncogene 2014, 33, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.H.; Chung, R.H.; Lee, W.J.; Lin, M.W.; Chuang, L.M.; Quertermous, T.; Assimes, T.; Hung, Y.J.; Yu, Y.W. Effect of common genetic variants of growth arrest-specific 6 gene on insulin resistance, obesity and type 2 diabetes in an Asian population. PLoS ONE 2015, 10, e0135681. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Li, Y.; Dai, W. Serum sex hormone and growth arrest-specific protein 6 levels in male patients with coronary heart disease. Asian J. Androl. 2016, 18, 644–649. [Google Scholar] [PubMed]
- Son, B.K.; Akishita, M.; Iijima, K.; Ogawa, S.; Maemura, K.; Yu, J.; Takeyama, K.; Kato, S.; Eto, M.; Ouchi, Y. Androgen receptor-dependent transactivation of growth arrest-specific gene 6 mediates inhibitory effects of testosterone on vascular calcification. J. Biol. Chem. 2010, 285, 7537–7544. [Google Scholar] [CrossRef] [PubMed]
- Qiao, M.; Wu, H.Y.; Guo, L.; Mei, S.Q.; Zhang, P.P.; Li, F.E.; Zheng, R.; Deng, C.Y. Imprinting analysis of porcine DIO3 gene in two fetal stages and association analysis with carcass and meat quality traits. Mol. Biol. Rep. 2012, 39, 2329–2335. [Google Scholar] [CrossRef] [PubMed]
- Beale, E.G.; Hammer, R.E.; Antoine, B.; Forest, C. Disregulated glyceroneogenesis: PCK1 as a candidate diabetes and obesity gene. Trends Endocrinol. Metab. 2004, 15, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Niang, F.; Benelli, C.; Ribière, C.; Collinet, M.; Mehebik-Mojaat, N.; Penot, G.; Forest, C.; Jaubert, A.M. Leptin induces nitric oxide-mediated inhibition of lipolysis and glyceroneogenesis in rat white adipose tissue. J. Nutr. 2011, 141, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Singh, D.; Zeng, Z.; Coleman, S.J.; Huang, Y.; Savich, G.L.; He, X.; Mieczkowski, P.; Grimm, S.A.; Perou, C.M.; et al. Mapsplice: Accurate mapping of RNA-seq reads for splice junction discovery. Nucleic Acids Res. 2010, 38, e178. [Google Scholar] [CrossRef] [PubMed]
- Miecznikowski, J.C.; Liu, S.; Ren, X. Statistical modeling for differential transcriptome analysis using RNA-seq technology. J. Solid Tumors 2012, 2, 33–44. [Google Scholar] [CrossRef]
- Dillies, M.A.; Rau, A.; Aubert, J.; Hennequet-Antier, C.; Jeanmougin, M.; Servant, N.; Keime, C.; Marot, G.; Castel, D.; Estelle, J.; et al. A comprehensive evaluation of normalization methods for illumina high-throughput RNA sequencing data analysis. Brief. Bioinform. 2013, 14, 671–683. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. Degseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Ernst, J.; Bar-Joseph, Z. Stem: A tool for the analysis of short time series gene expression data. BMC Bioinform. 2006, 7, 191. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆Ct method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Terms | IM | CM | CMT |
|---|---|---|---|
| All reads | 26,818,106 | 24,658,796 | 23,870,297 |
| Un mapped reads | 4,906,529 | 4,295,752 | 4,089,813 |
| Mapped reads | 21,911,577 | 20,363,044 | 19,780,484 |
| Mapping rate | 0.817 | 0.826 | 0.829 |
| Unique mapping | 20,705,544 | 19,206,414 | 18,686,768 |
| Unique mapping rate | 0.772 | 0.779 | 0.783 |
| Repeat mapping | 1,206,744 | 1,157,303 | 1,094,352 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Cai, Y.; Wei, S.; Ling, Y.; Zhu, L.; Li, D.; Cai, Z. Testosterone Deficiency Induces Changes of the Transcriptomes of Visceral Adipose Tissue in Miniature Pigs Fed a High-Fat and High-Cholesterol Diet. Int. J. Mol. Sci. 2016, 17, 2125. https://doi.org/10.3390/ijms17122125
Zhang L, Cai Y, Wei S, Ling Y, Zhu L, Li D, Cai Z. Testosterone Deficiency Induces Changes of the Transcriptomes of Visceral Adipose Tissue in Miniature Pigs Fed a High-Fat and High-Cholesterol Diet. International Journal of Molecular Sciences. 2016; 17(12):2125. https://doi.org/10.3390/ijms17122125
Chicago/Turabian StyleZhang, Lifan, Yueqin Cai, Shengjuan Wei, Yun Ling, Liang Zhu, Dongfeng Li, and Zhaowei Cai. 2016. "Testosterone Deficiency Induces Changes of the Transcriptomes of Visceral Adipose Tissue in Miniature Pigs Fed a High-Fat and High-Cholesterol Diet" International Journal of Molecular Sciences 17, no. 12: 2125. https://doi.org/10.3390/ijms17122125
APA StyleZhang, L., Cai, Y., Wei, S., Ling, Y., Zhu, L., Li, D., & Cai, Z. (2016). Testosterone Deficiency Induces Changes of the Transcriptomes of Visceral Adipose Tissue in Miniature Pigs Fed a High-Fat and High-Cholesterol Diet. International Journal of Molecular Sciences, 17(12), 2125. https://doi.org/10.3390/ijms17122125