Therapeutic Effect of Losartan, an Angiotensin II Type 1 Receptor Antagonist, on CCl4-Induced Skeletal Muscle Injury


 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
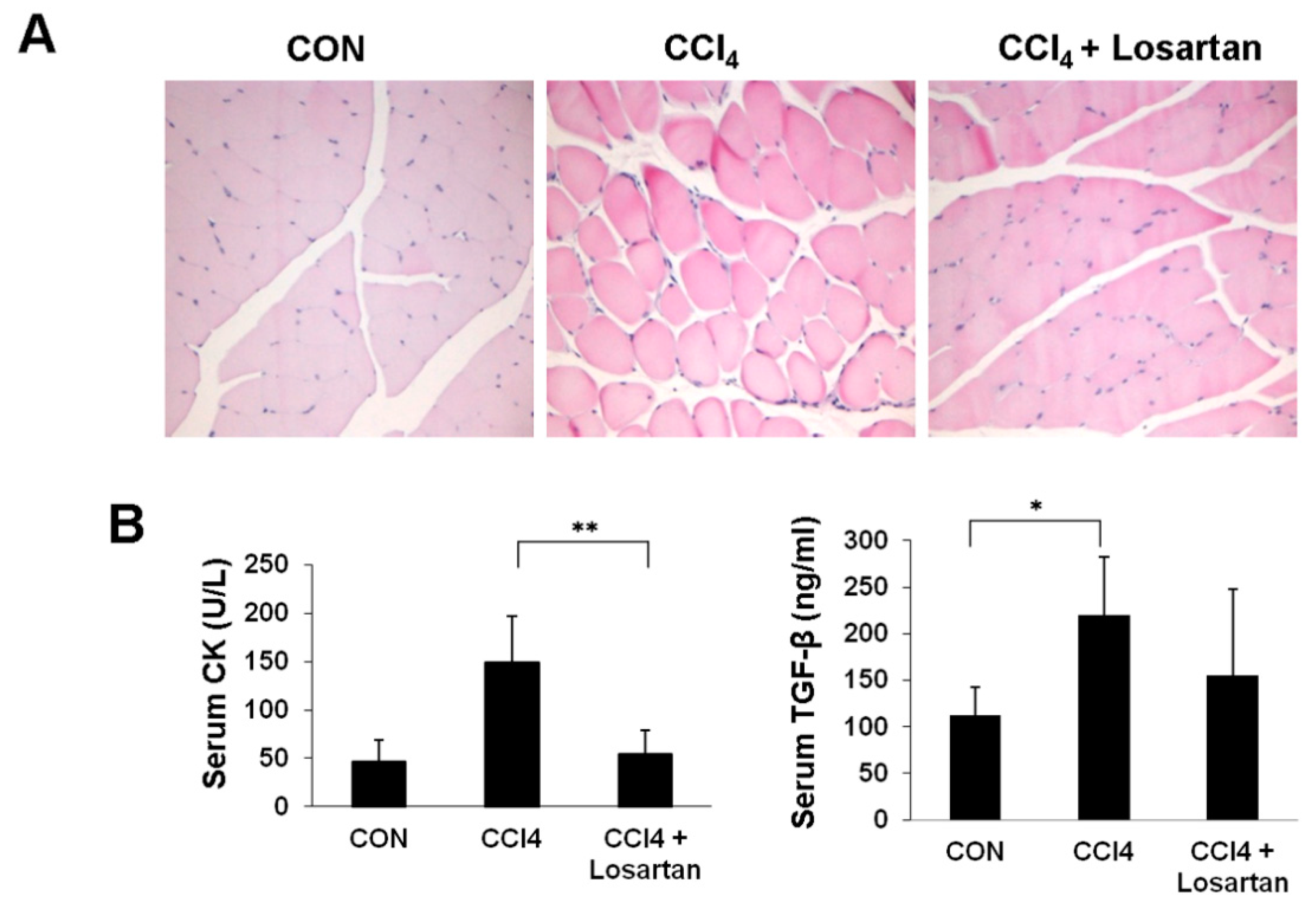
2.1. Chronic CCl4 Injection Induces Muscle Damage
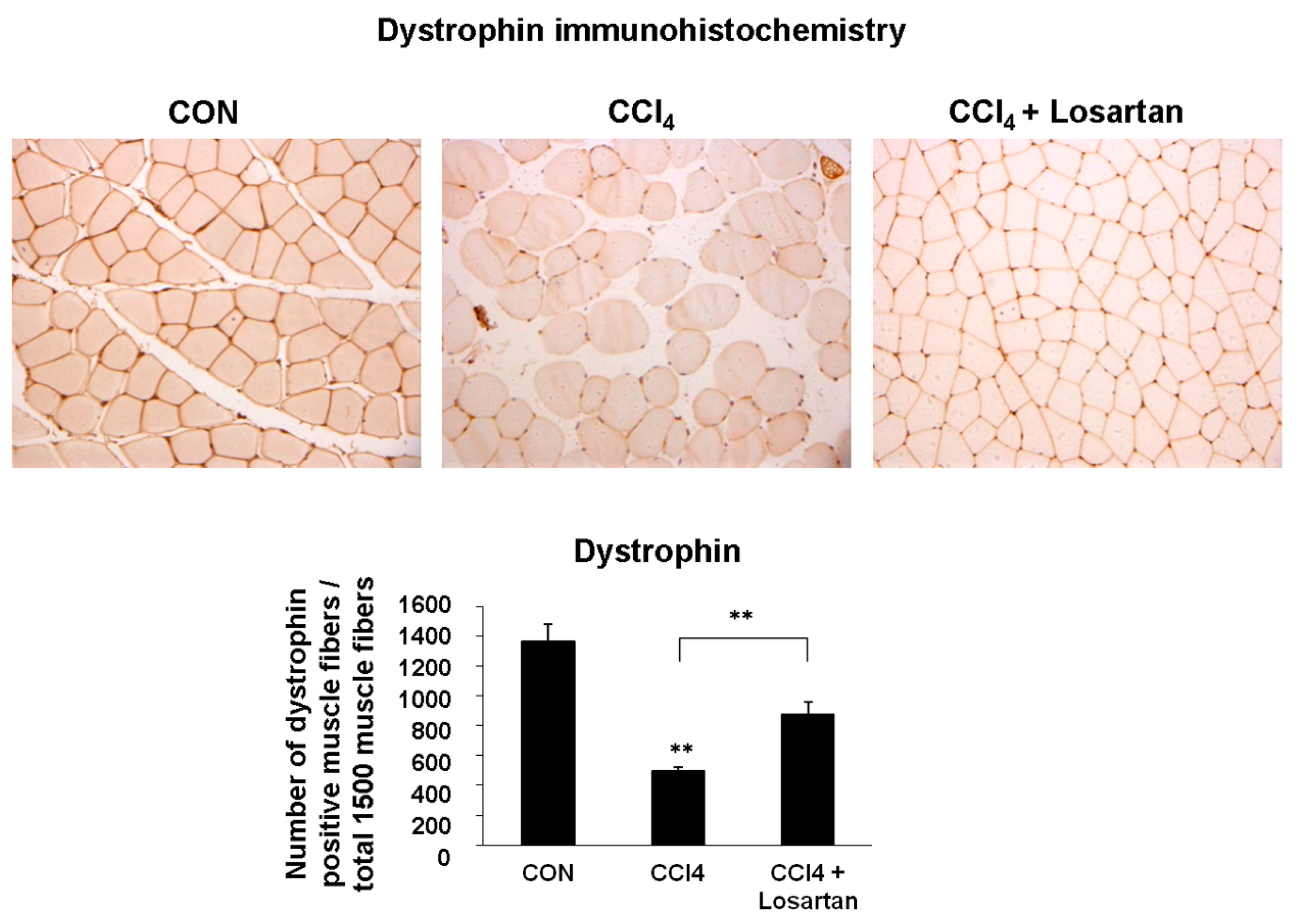
2.2. Expression of Dystrophin in Skeletal Muscle


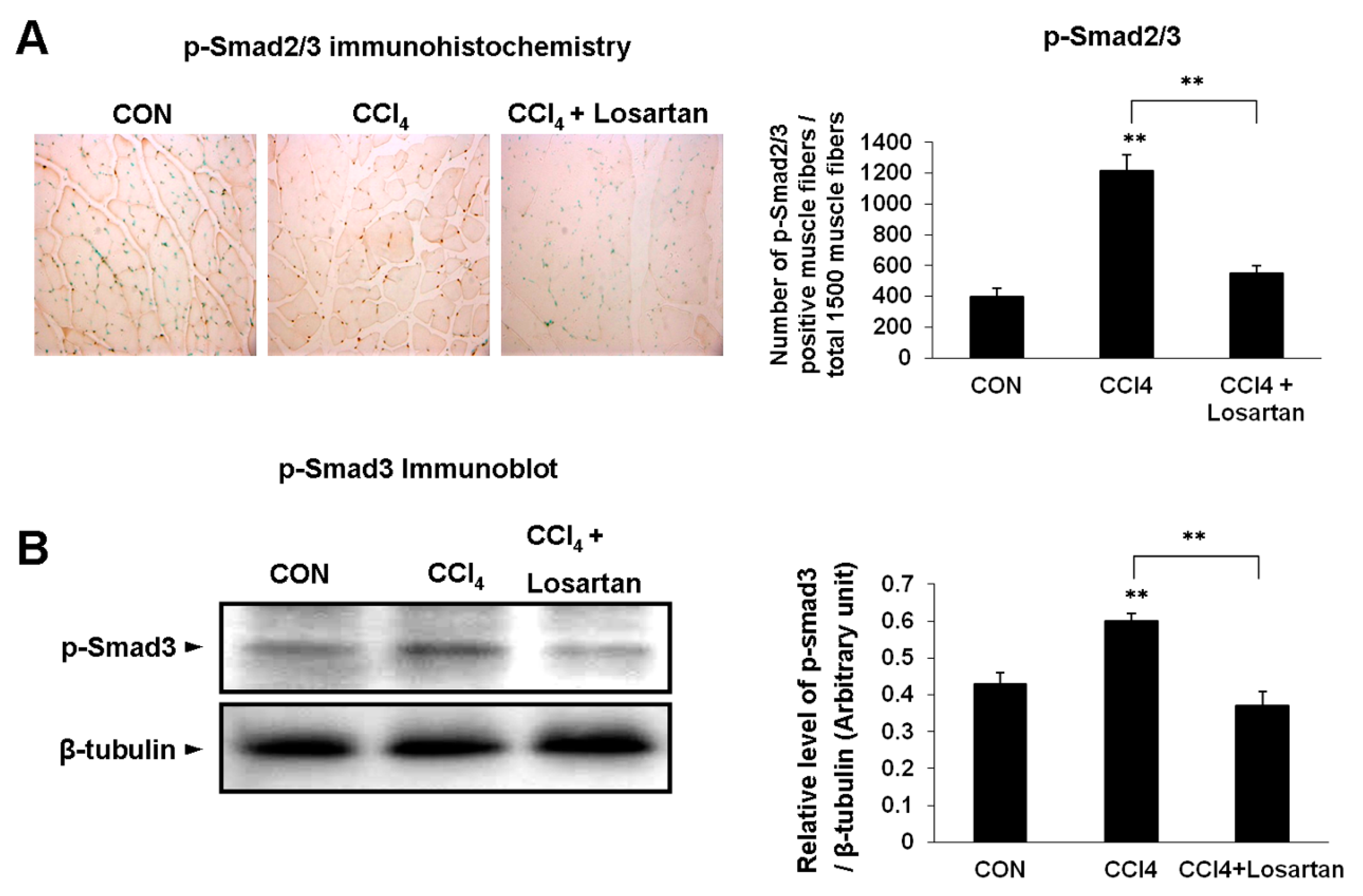
2.3. Expression of p(phospho)-Smad2/3 and p-Smad3 in Skeletal Muscle

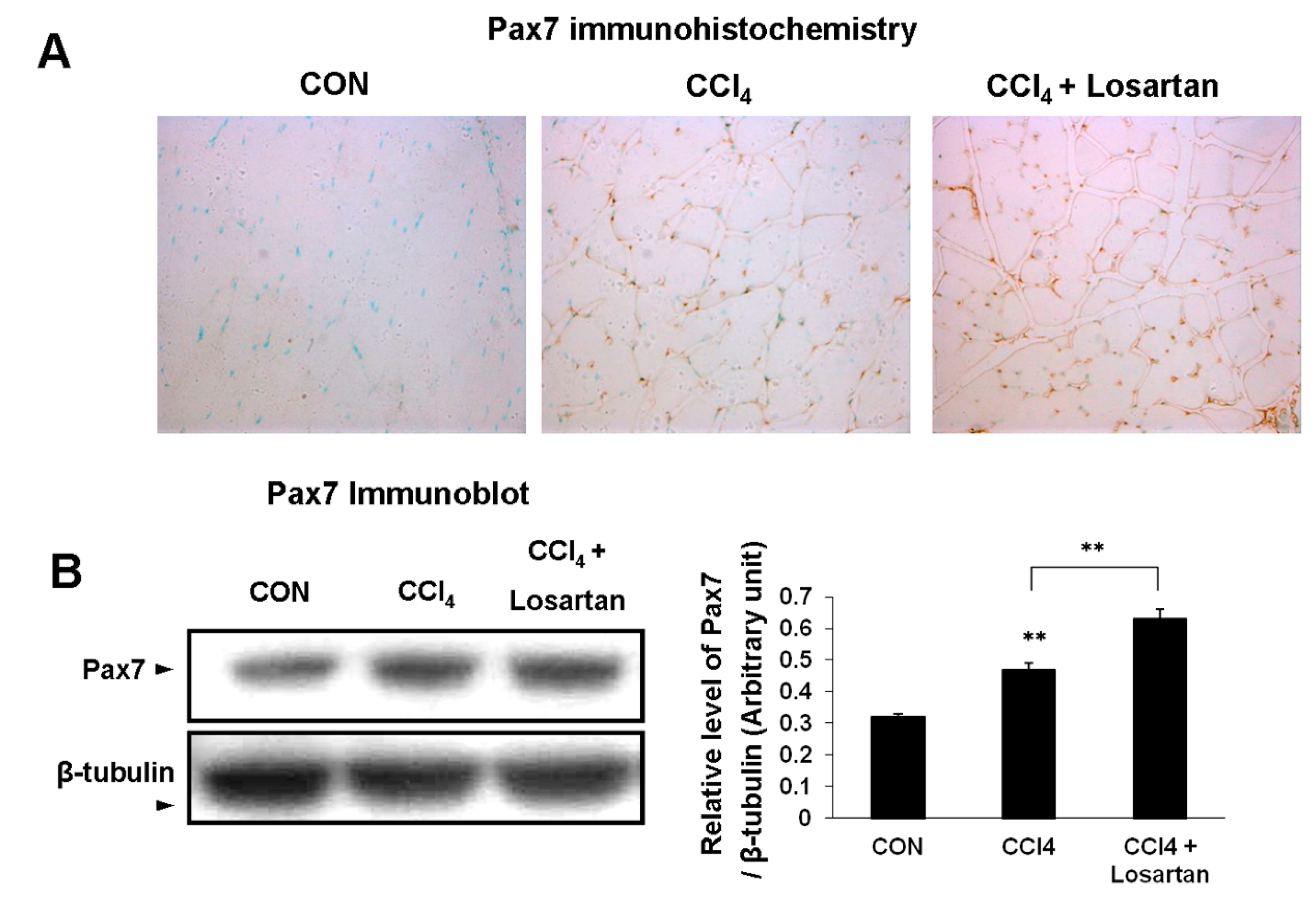
2.4. Expression of Pax7 in Skeletal Muscle
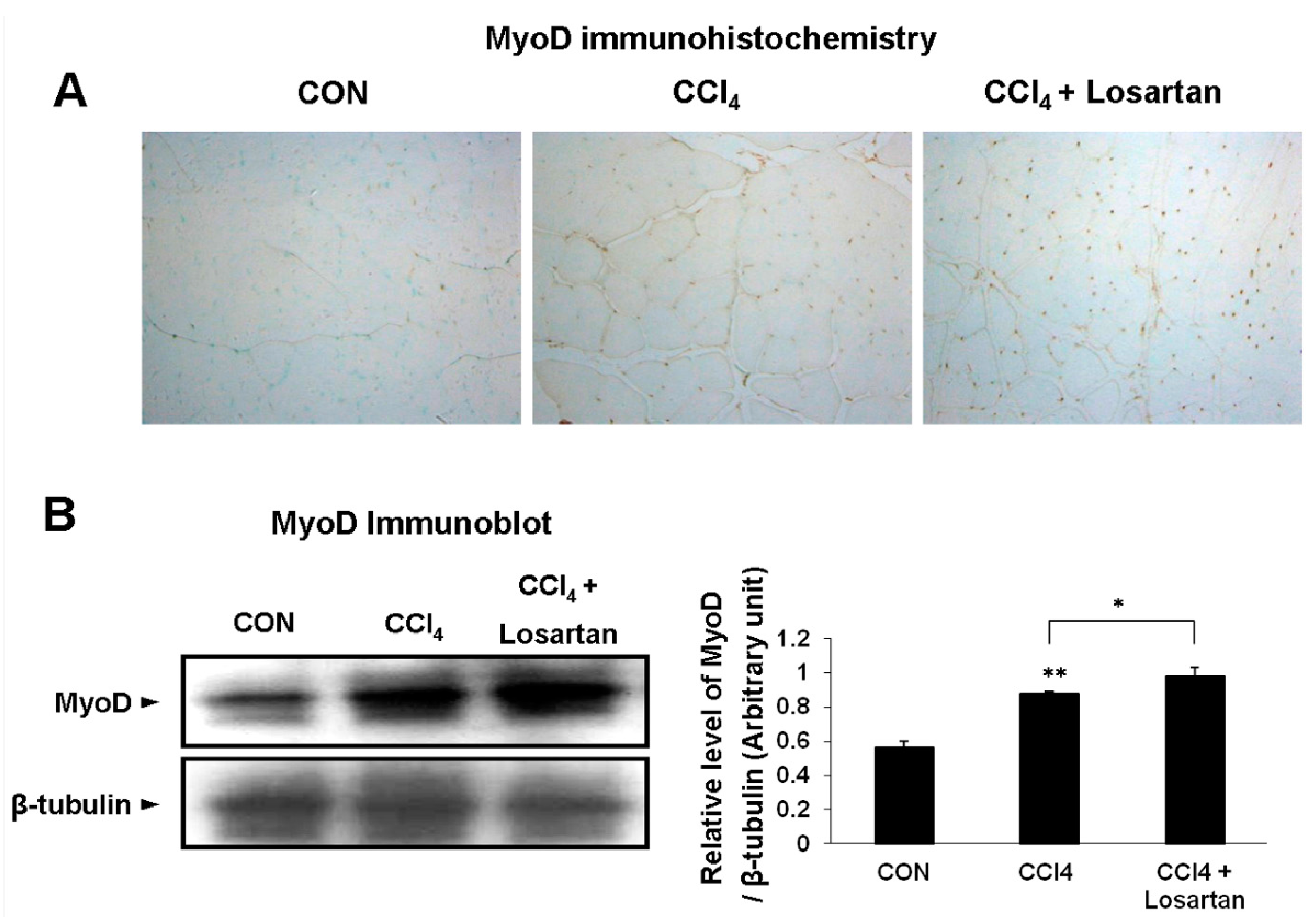
2.5. Expression of MyoD in Skeletal Muscle


2.6. Expression of Myogenin in Skeletal Muscle

3. Discussion
4. Experimental Section
4.1. Animals and Experimental Design
4.2. Serum Biochemistry
4.3. Histopathology and Immunohistochemistry
4.4. Immunoblot Analysis
4.5. Statistical Analysis
5. Conclsions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Weber, L.W.; Boll, M.; Stampfl, A. Hepatotoxicity and mechanism of action of haloalkanes: Carbon tetrachloride as a toxicological model. Crit. Rev. Toxicol. 2003, 33, 105–136. [Google Scholar] [CrossRef] [PubMed]
- Raucy, J.L.; Kraner, J.C.; Lasker, J.M. Bioactivation of halogenated hydrocarbons by cytochrome P4502E1. Crit. Rev. Toxicol. 1993, 23, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Zangar, R.C.; Benson, J.M.; Burnett, V.L.; Springer, D.L. Cytochrome P450 2E1 is the primary enzyme responsible for low-dose carbon tetrachloride metabolism in human liver microsomes. Chem. Biol. Interact. 2000, 125, 233–243. [Google Scholar] [CrossRef]
- Donthamsetty, S.; Bhave, V.S.; Mitra, M.S.; Latendresse, J.R.; Mehendale, H.M. Nonalcoholic fatty liver sensitizes rats to carbon tetrachloride hepatotoxicity. Hepatology 2007, 45, 391–403. [Google Scholar] [CrossRef] [PubMed]
- Park, J.K.; Jeong, D.H.; Park, H.Y.; Son, K.H.; Shin, D.H.; Do, S.H.; Lee, H.R. Hepatoprotective effect of arazyme on CCl 4-induced acute hepatic injury in SMP30 knock-out mice. Toxicology 2008, 246, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Heldin, C.H.; Miyazono, K.; ten Dijke, P. TGF-β signalling from cell membrane to nucleus through SMAD proteins. Nature 1997, 390, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Cohn, R.D.; van Erp, C.; Habashi, J.P.; Soleimani, A.A.; Klein, E.C.; Lisi, M.T.; Gamradt, M.; Rhys, C.M.; Holm, T.M.; Loeys, B.L.; et al. Angiotensin II type 1 receptor blockade attenuates TGF-β-induced failure of muscle regeneration in multiple myopathic states. Nat. Med. 2007, 13, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Olson, E.N.; Sternberg, E.; Hu, J.S.; Spizz, G.; Wilcox, C. Regulation of myogenic differentiation by type β transforming growth factor. J. Cell Biol. 1986, 103, 1799–1805. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Foster, W.; Deasy, B.M.; Chan, Y.; Prisk, V.; Tang, Y.; Cummins, J.; Huard, J. Transforming growth factor-β1 induces the differentiation of myogenic cells into fibrotic cells in injured skeletal muscle: A key event in muscle fibrogenesis. Am. J. Pathol. 2004, 164, 1007–1019. [Google Scholar] [CrossRef]
- Bernasconi, P.; Torchiana, E.; Confalonieri, P.; Brugnoni, R.; Barresi, R.; Mora, M.; Cornelio, F.; Morandi, L.; Mantegazza, R. Expression of transforming growth factor-β 1 in dystrophic patient muscles correlates with fibrosis. Pathogenetic role of a fibrogenic cytokine. J. Clin. Investig. 1995, 96, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Dina, R.; Jafari, M. Angiotensin II-receptor antagonists: An overview. Am. J. Health Syst. Pharm. 2000, 57, 1231–1241. [Google Scholar] [PubMed]
- Onder, G.; Vedova, C.D.; Pahor, M. Effects of ACE inhibitors on skeletal muscle. Curr. Pharm. Des. 2006, 12, 2057–2064. [Google Scholar] [CrossRef] [PubMed]
- Bedair, H.S.; Karthikeyan, T.; Quintero, A.; Li, Y.; Huard, J. Angiotensin II receptor blockade administered after injury improves muscle regeneration and decreases fibrosis in normal skeletal muscle. Am. J. Sports Med. 2008, 36, 1548–1554. [Google Scholar] [CrossRef] [PubMed]
- Collins, C.A.; Zammit, P.S.; Ruiz, A.P.; Morgan, J.E.; Partridge, T.A. A population of myogenic stem cells that survives skeletal muscle aging. Stem Cells 2007, 25, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Olguin, H.C.; Olwin, B.B. Pax-7 up-regulation inhibits myogenesis and cell cycle progression in satellite cells: A potential mechanism for self-renewal. Dev. Biol. 2004, 275, 375–388. [Google Scholar] [CrossRef] [PubMed]
- Dedkov, E.I.; Kostrominova, T.Y.; Borisov, A.B.; Carlson, B.M. MyoD and myogenin protein expression in skeletal muscles of senile rats. Cell Tissue Res. 2003, 311, 401–416. [Google Scholar] [PubMed]
- Le Grand, F.; Rudnicki, M.A. Skeletal muscle satellite cells and adult myogenesis. Curr. Opin. Cell Biol. 2007, 19, 628–633. [Google Scholar] [CrossRef] [PubMed]
- Lamandé, S.R.; North, K.N. Activating internal ribosome entry to treat Duchenne muscular dystrophy. Nat. Med. 2014, 20, 987–988. [Google Scholar] [CrossRef] [PubMed]
- Blake, D.J.; Weir, A.; Newey, S.E.; Davies, K.E. Function and genetics of dystrophin and dystrophin-related proteins in muscle. Physiol. Rev. 2002, 82, 291–329. [Google Scholar] [CrossRef] [PubMed]
- Mitra, A.; Satelli, A.; Yan, J.; Xueqing, X.; Gagea, M.; Hunter, C.A.; Mishra, L.; Li, S. IL-30 (IL27p28) attenuates liver fibrosis through inducing NKG2D-rae1 interaction between NKT and activated hepatic stellate cells in mice. Hepatology 2014, 60, 2027–2039. [Google Scholar] [CrossRef] [PubMed]
- Pan, R.L.; Xiang, L.X.; Wang, P.; Liu, X.Y.; Nie, L.; Huang, W.; Shao, J.Z. Low-molecular-weight fibroblast growth factor 2 attenuates hepatic fibrosis by epigenetic down-regulation of δ-like1. Hepatology 2015, 61, 1708–1720. [Google Scholar] [CrossRef] [PubMed]
- Park, J.K.; Ki, M.R.; Lee, H.R.; Hong, I.H.; Ji, A.R.; Ishigami, A.; Park, S.I.; Kim, J.M.; Chung, H.Y.; Yoo, S.E.; et al. Vitamin C deficiency attenuates liver fibrosis by way of up-regulated peroxisome proliferator-activated receptor-γ expression in senescence marker protein 30 knockout mice. Hepatology 2010, 51, 1766–1777. [Google Scholar] [CrossRef] [PubMed]
- Tahashi, Y.; Matsuzaki, K.; Date, M.; Yoshida, K.; Furukawa, F.; Sugano, Y.; Matsushita, M.; Himeno, Y.; Inagaki, Y.; Inoue, K. Differential regulation of TGF-β signal in hepatic stellate cells between acute and chronic rat liver injury. Hepatology 2002, 35, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Acharjee, S.; Chung, T.K.; Gopinadhan, S.; Shankar, S.R.; Wang, Y.; Li, L.; Vercherat, C.; Gulbagci, N.T.; Rossner, M.; Taneja, R. Sharp-1 regulates TGF-β signaling and skeletal muscle regeneration. J. Cell Sci. 2014, 127, 599–608. [Google Scholar]
- Burks, T.N.; Andres-Mateos, E.; Marx, R.; Mejias, R.; van Erp, C.; Simmers, J.L.; Walston, J.D.; Ward, C.W.; Cohn, R.D. Losartan restores skeletal muscle remodeling and protects against disuse atrophy in sarcopenia. Sci. Transl. Med. 2011, 3, 82ra37. [Google Scholar] [CrossRef] [PubMed]
- Park, J.K.; Ki, M.R.; Lee, E.M.; Kim, A.Y.; You, S.Y.; Han, S.Y.; Lee, E.J.; Hong, I.H.; Kwon, S.H.; Kim, S.J.; et al. Losartan improves adipose tissue-derived stem cell niche by inhibiting transforming growth factor-β and fibrosis in skeletal muscle injury. Cell Transplant. 2012, 21, 2407–2424. [Google Scholar] [CrossRef] [PubMed]
- Arthur, P.G.; Grounds, M.D.; Shavlakadze, T. Oxidative stress as a therapeutic target during muscle wasting: Considering the complex interactions. Curr. Opin. Clin. Nutr. Metab. Care 2008, 11, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Moylan, J.S.; Reid, M.B. Oxidative stress, chronic disease, and muscle wasting. Muscle Nerve 2007, 35, 411–429. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Kavazis, A.N.; McClung, J.M. Oxidative stress and disuse muscle atrophy. J. Appl. Physiol. 2007, 102, 2389–2397. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.A.; Stauber, F.; Waters, C.; Always, S.E.; Stauber, W.T. Transforming growth factor-β following skeletal muscle strain injury in rats. J. Appl. Physiol. 2007, 102, 755–761. [Google Scholar] [CrossRef] [PubMed]
- Bedair, H.; Liu, T.T.; Kaar, J.L.; Badlani, S.; Russell, A.J.; Li, Y.; Huard, J. Matrix metalloproteinase-1 therapy improves muscle healing. J. Appl. Physiol. 2007, 102, 2338–2345. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.S.; Li, Y.; Foster, W.; Horaguchi, T.; Somogyi, G.; Fu, F.H.; Huard, J. Antifibrotic effects of suramin in injured skeletal muscle after laceration. J. Appl. Physiol. 2003, 95, 771–780. [Google Scholar] [CrossRef] [PubMed]
- Foster, W.; Li, Y.; Usas, A.; Somogyi, G.; Huard, J. γ interferon as an antifibrosis agent in skeletal muscle. J. Orthop. Res. 2003, 21, 798–804. [Google Scholar] [CrossRef]
- Fukushima, K.; Badlani, N.; Usas, A.; Riano, F.; Fu, F.; Huard, J. The use of an antifibrosis agent to improve muscle recovery after laceration. Am. J. Sports Med. 2001, 29, 394–402. [Google Scholar] [PubMed]
- Lim, D.S.; Lutucuta, S.; Bachireddy, P.; Youker, K.; Evans, A.; Entman, M.; Roberts, R.; Marian, A.J. Angiotensin II blockade reverses myocardial fibrosis in a transgenic mouse model of human hypertrophic cardiomyopathy. Circulation 2001, 103, 789–791. [Google Scholar] [CrossRef] [PubMed]
- Kunert-Keil, C.; Gredes, T.; Lucke, S.; Morgenstern, S.; Mielczarek, A.; Sporniak-Tutak, K.; Gedrange, T.; Spassov, A. Caveolin-1, caveolin-3 and VEGF expression in the masticatory muscles of mdx mice. Folia Histochem. Cytobiol. 2011, 49, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Gopinath, S.D.; Rando, T.A. Stem cell review series: Aging of the skeletal muscle stem cell niche. Aging Cell 2008, 7, 590–598. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Bi, P.; Liu, W.; Asakura, A.; Keller, C.; Kuang, S. Constitutive notch activation upregulates pax7 and promotes the self-renewal of skeletal muscle satellite cells. Mol. Cell. Biol. 2012, 32, 2300–2311. [Google Scholar] [CrossRef] [PubMed]
- Brzoska, E.; Ciemerych, M.A.; Przewozniak, M.; Zimowska, M. Regulation of muscle stem cells activation: The role of growth factors and extracellular matrix. Vitam. Horm. 2011, 87, 239–276. [Google Scholar] [PubMed]
- Carosio, S.; Berardinelli, M.G.; Aucello, M.; Musarò, A. Impact of ageing on muscle cell regeneration. Ageing Res. Rev. 2011, 10, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Carlson, M.E.; Conboy, M.J.; Hsu, M.; Barchas, L.; Jeong, J.; Agrawal, A.; Mikels, A.J.; Agrawal, S.; Schaffer, D.V.; Conboy, I.M. Relative roles of TGF-β1 and Wnt in the systemic regulation and aging of satellite cell responses. Aging Cell 2009, 8, 676–689. [Google Scholar] [CrossRef] [PubMed]
- Fakhfakh, R.; Lamarre, Y.; Skuk, D.; Tremblay, J.P. Losartan enhances the success of myoblast transplantation. Cell Transplant. 2012, 21, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Gressner, A.M.; Weiskirchen, R.; Breitkopf, K.; Dooley, S. Roles of TGF-β in hepatic fibrosis. Front. Biosci. 2002, 7, d793–807. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.; Gonzalo, T.; Kok, R.J.; Sancho-Bru, P.; van Beuge, M.; Swart, J.; Prakash, J.; Temming, K.; Fondevila, C.; Beljaars, L.; et al. Reduction of advanced liver fibrosis by short-term targeted delivery of an angiotensin receptor blocker to hepatic stellate cells in rats. Hepatology 2010, 51, 942–952. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, O.-K.; Park, J.-K.; Lee, E.-J.; Lee, E.-M.; Kim, A.-Y.; Jeong, K.-S. Therapeutic Effect of Losartan, an Angiotensin II Type 1 Receptor Antagonist, on CCl4-Induced Skeletal Muscle Injury. Int. J. Mol. Sci. 2016, 17, 227. https://doi.org/10.3390/ijms17020227
Hwang O-K, Park J-K, Lee E-J, Lee E-M, Kim A-Y, Jeong K-S. Therapeutic Effect of Losartan, an Angiotensin II Type 1 Receptor Antagonist, on CCl4-Induced Skeletal Muscle Injury. International Journal of Molecular Sciences. 2016; 17(2):227. https://doi.org/10.3390/ijms17020227
Chicago/Turabian StyleHwang, Ok-Kyung, Jin-Kyu Park, Eun-Joo Lee, Eun-Mi Lee, Ah-Young Kim, and Kyu-Shik Jeong. 2016. "Therapeutic Effect of Losartan, an Angiotensin II Type 1 Receptor Antagonist, on CCl4-Induced Skeletal Muscle Injury" International Journal of Molecular Sciences 17, no. 2: 227. https://doi.org/10.3390/ijms17020227
APA StyleHwang, O. -K., Park, J. -K., Lee, E. -J., Lee, E. -M., Kim, A. -Y., & Jeong, K. -S. (2016). Therapeutic Effect of Losartan, an Angiotensin II Type 1 Receptor Antagonist, on CCl4-Induced Skeletal Muscle Injury. International Journal of Molecular Sciences, 17(2), 227. https://doi.org/10.3390/ijms17020227