
MicroRNA and Heart Failure


 ,
,
Abstract
:1. Introduction
2. MicroRNA Discovery and Biogenesis
3. Landmark MicroRNA Studies in Cardiovascular Field
3.1. Dicer in Cardiac Development
3.2. Specific miRNAs in Cardiac Development
3.2.1. miR-1 and the miR-133a Superfamily
3.2.2. miR-208 and the miR-499 Superfamily
3.2.3. miR-15 Family
3.3. miRNAs in Vascular Integrity
4. Heart Failure
4.1. Etiology of Heart Failure
4.2. Neurohormonal Signaling and Heart Failure
4.3. Diagnosis of Heart Failure
4.4. Treatment of Heart Failure
5.miRNAs in Heart Failure
Putative miRNA Targets and Neurohormone
6. Challenges of MicroRNA Research in Heart Failure
6.1. Consistent miRNA Profiles in Heart Failure Are Yet to Be Identified
6.2. Obstacles to miRNA Therapy in Cardiovascular Diseases
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Eric, J.; Topol, R.M.C. Textbook of Cardiovascular Medicine; Lippincott Williams & Wilkins: New York, NY, USA, 2007. [Google Scholar]
- Vogel, B.; Keller, A.; Frese, K.S.; Leidinger, P.; Sedaghat-Hamedani, F.; Kayvanpour, E.; Kloos, W.; Backe, C.; Thanaraj, A.; Brefort, T.; et al. Multivariate miRNA signatures as biomarkers for non-ischaemic systolic heart failure. Eur. Heart J. 2013, 34, 2812–2822. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Wang, H.; Wu, Q. Atrial natriuretic peptide in cardiovascular biology and disease (NPPA). Gene 2015, 569, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Yandle, T.G.; Richards, A.M. B-type Natriuretic Peptide circulating forms: Analytical and bioactivity issues. Clin. Chim. Acta 2015, 448, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Rademaker, M.T.; Cameron, V.A.; Charles, C.J.; Lainchbury, J.G.; Nicholls, M.G.; Richards, A.M. Adrenomedullin and heart failure. Regul. Pept. 2003, 112, 51–60. [Google Scholar] [CrossRef]
- Vardeny, O.; Miller, R.; Solomon, S.D. Combined neprilysin and renin-angiotensin system inhibition for the treatment of heart failure. JACC Heart Fail. 2014, 2, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, R.L.; Hullinger, T.G.; Semus, H.M.; Dickinson, B.A.; Seto, A.G.; Lynch, J.M.; Stack, C.; Latimer, P.A.; Olson, E.N.; van Rooij, E. Therapeutic inhibition of miR-208a improves cardiac function and survival during heart failure. Circulation 2011, 124, 1537–1547. [Google Scholar] [CrossRef] [PubMed]
- Van Rooij, E.; Olson, E.N. MicroRNA therapeutics for cardiovascular disease: Opportunities and obstacles. Nat. Rev. Drug Discov. 2012, 11, 860–872. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Wightman, B.; Ha, I.; Ruvkun, G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell 1993, 75, 855–862. [Google Scholar] [CrossRef]
- Bentwich, I.; Avniel, A.; Karov, Y.; Aharonov, R.; Gilad, S.; Barad, O.; Barzilai, A.; Einat, P.; Einav, U.; Meiri, E.; et al. Identification of hundreds of conserved and nonconserved human microRNAs. Nat. Genet. 2005, 37, 766–770. [Google Scholar] [CrossRef] [PubMed]
- Kozomara, A.; Griffiths-Jones, S. MiRBase: Annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2014, 42, D68–D73. [Google Scholar] [CrossRef] [PubMed]
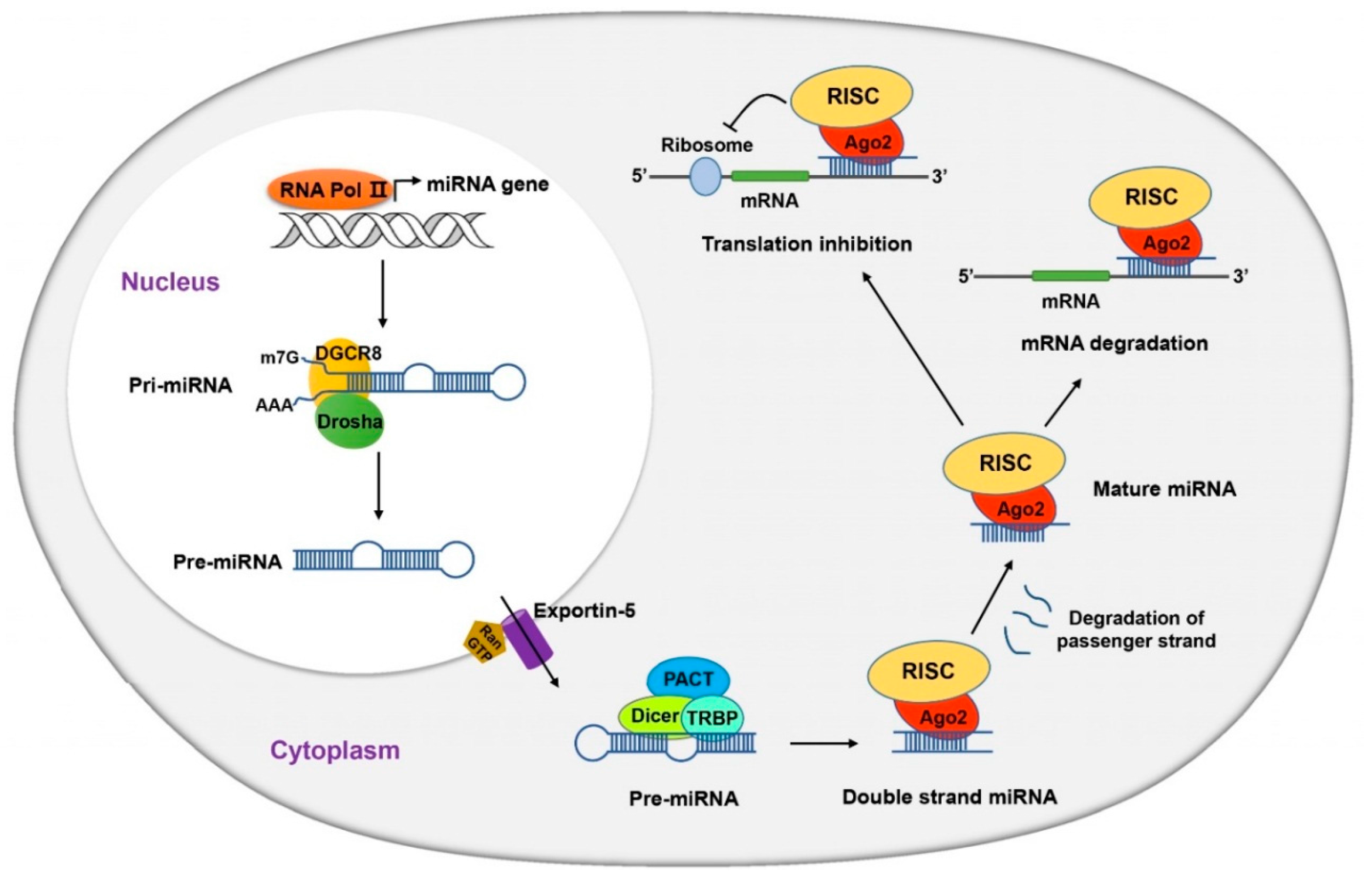
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Therapeutics, M. A Multicenter Phase I Study of MRX34, MicroRNA miR-RX34 Liposomal Injection. Available online: https://www.clinicaltrials.gov/ct2/show/NCT01829971?term=mirna+therapeutics&rank=115 (accessed on 3 December 2015).
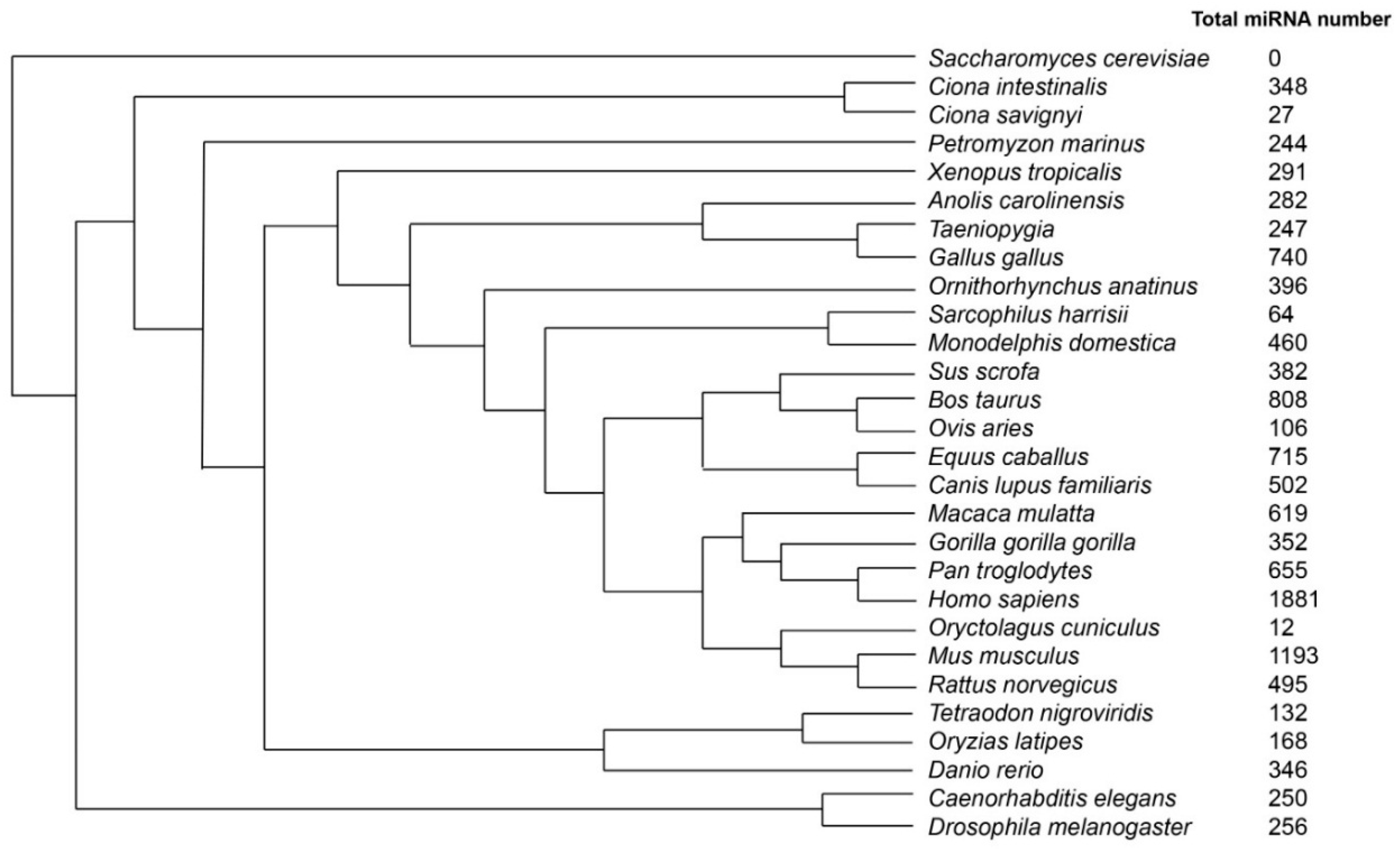
- Species Tree. Available online: http://www.ensembl.org/info/about/speciestree.html (accessed on 29 September 2015).
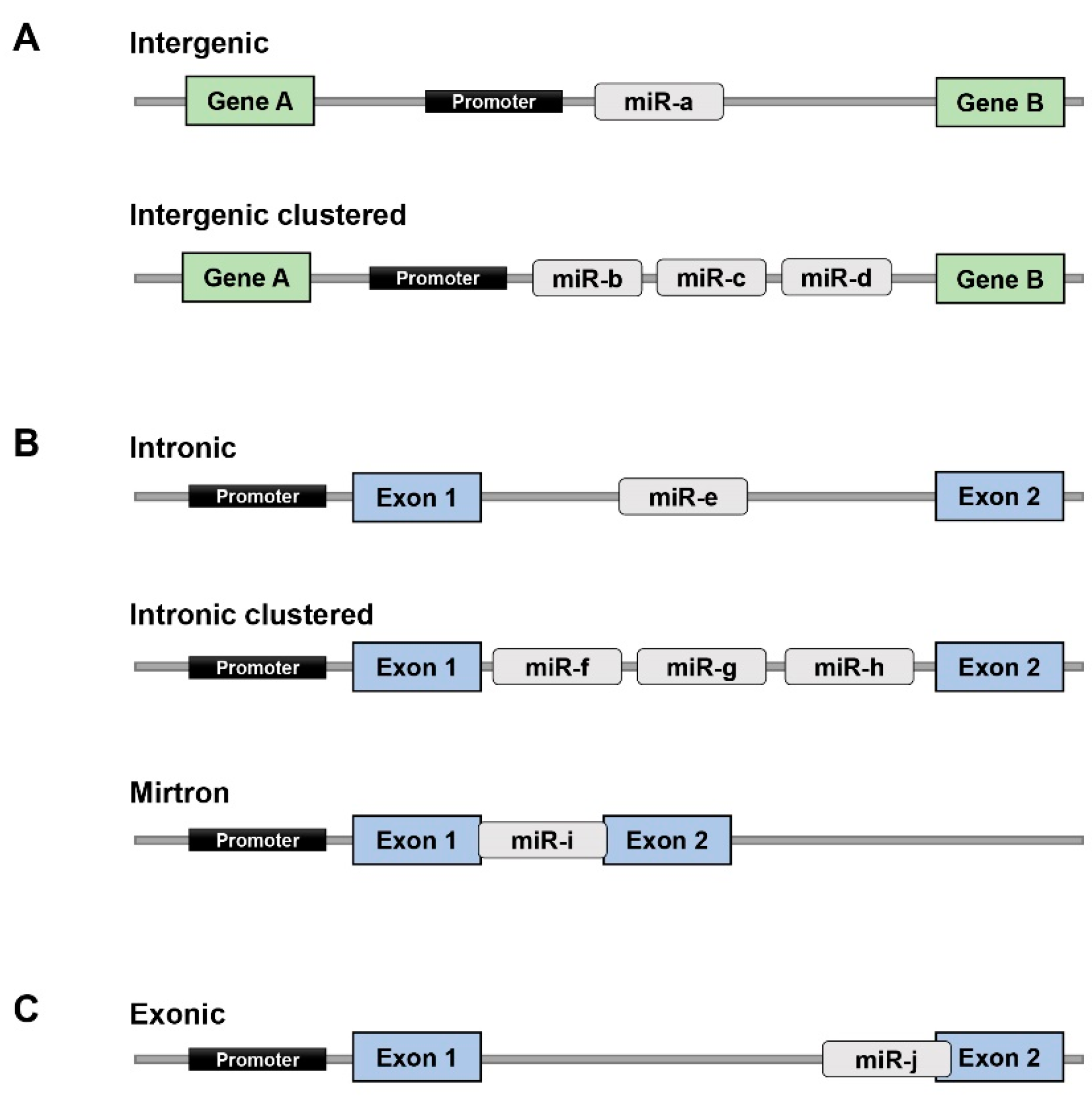
- Olena, A.F.; Patton, J.G. Genomic organization of microRNAs. J. Cell. Physiol. 2010, 222, 540–545. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.; Griffiths-Jones, S.; Ashurst, J.L.; Bradley, A. Identification of mammalian microRNA host genes and transcription units. Genome Res. 2004, 14, 1902–1910. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.H.; Lee, S.; Baek, S.H.; Kim, V.N. MicroRNA genes are transcribed by RNA polymerase II. EMBO J. 2004, 23, 4051–4060. [Google Scholar] [CrossRef] [PubMed]
- Kim, V.N.; Han, J.; Siomi, M.C. Biogenesis of small RNAs in animals. Nat. Rev. Mol. Cell Biol. 2009, 10, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Pillai, R.S. MicroRNA function: Multiple mechanisms for a tiny RNA? RNA 2005, 11, 1753–1761. [Google Scholar] [CrossRef] [PubMed]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Hausser, J.; Zavolan, M. Identification and consequences of miRNA-target interactions—Beyond repression of gene expression. Nat. Rev. Genet. 2014, 15, 599–612. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Hagedorn, C.H.; Cullen, B.R. Human microRNAs are processed from capped, polyadenylated transcripts that can also function as mRNAs. RNA 2004, 10, 1957–1966. [Google Scholar] [CrossRef] [PubMed]
- Leuschner, P.J.; Ameres, S.L.; Kueng, S.; Martinez, J. Cleavage of the siRNA passenger strand during RISC assembly in human cells. EMBO Rep. 2006, 7, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Hata, A. Functions of microRNAs in cardiovascular biology and disease. Annu. Rev. Physiol. 2013, 75, 69–93. [Google Scholar] [CrossRef] [PubMed]
- Novak, J.; Olejnickova, V.; Tkacova, N.; Santulli, G. Mechanistic Role of MicroRNAs in Coupling Lipid Metabolism and Atherosclerosis. Adv. Exp. Med. Biol. 2015, 887, 79–100. [Google Scholar] [PubMed]
- Latronico, M.V.; Condorelli, G. MicroRNAs and cardiac pathology. Nat. Rev. Cardiol. 2009, 6, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Park, C.Y.; Choi, Y.S.; McManus, M.T. Analysis of microRNA knockouts in mice. Hum. Mol. Genet. 2010, 19, R169–R175. [Google Scholar] [CrossRef] [PubMed]
- Santulli, G.; Iaccarino, G.; de Luca, N.; Trimarco, B.; Condorelli, G. Atrial fibrillation and microRNAs. Front. Physiol. 2014, 5, 15. [Google Scholar] [CrossRef] [PubMed]
- Wronska, A.; Kurkowska-Jastrzebska, I.; Santulli, G. Application of microRNAs in diagnosis and treatment of cardiovascular disease. Acta Physiol. (Oxf.) 2015, 213, 60–83. [Google Scholar] [CrossRef] [PubMed]
- Ono, K. MicroRNAs and Cardiovascular Remodeling. Adv. Exp. Med. Biol. 2015, 888, 197–213. [Google Scholar] [PubMed]
- Gnecchi, M.; Pisano, F.; Bariani, R. MicroRNA and Cardiac Regeneration. Adv. Exp. Med. Biol. 2015, 887, 119–141. [Google Scholar] [PubMed]
- Bernstein, E.; Kim, S.Y.; Carmell, M.A.; Murchison, E.P.; Alcorn, H.; Li, M.Z.; Mills, A.A.; Elledge, S.J.; Anderson, K.V.; Hannon, G.J. Dicer is essential for mouse development. Nat. Genet. 2003, 35, 215–217. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Carmell, M.A.; Rivas, F.V.; Marsden, C.G.; Thomson, J.M.; Song, J.J.; Hammond, S.M.; Joshua-Tor, L.; Hannon, G.J. Argonaute2 is the catalytic engine of mammalian RNAi. Science 2004, 305, 1437–1441. [Google Scholar] [CrossRef] [PubMed]
- Kanellopoulou, C.; Muljo, S.A.; Kung, A.L.; Ganesan, S.; Drapkin, R.; Jenuwein, T.; Livingston, D.M.; Rajewsky, K. Dicer-deficient mouse embryonic stem cells are defective in differentiation and centromeric silencing. Genes Dev. 2005, 19, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Saxena, A.; Tabin, C.J. miRNA-processing enzyme Dicer is necessary for cardiac outflow tract alignment and chamber septation. Proc. Natl. Acad. Sci. USA 2010, 107, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.F.; Murchison, E.P.; Tang, R.; Callis, T.E.; Tatsuguchi, M.; Deng, Z.; Rojas, M.; Hammond, S.M.; Schneider, M.D.; Selzman, C.H.; et al. Targeted deletion of Dicer in the heart leads to dilated cardiomyopathy and heart failure. Proc. Natl. Acad. Sci. USA 2008, 105, 2111–2116. [Google Scholar] [CrossRef] [PubMed]
- Da Costa Martins, P.A.; Bourajjaj, M.; Gladka, M.; Kortland, M.; van Oort, R.J.; Pinto, Y.M.; Molkentin, J.D.; de Windt, L.J. Conditional dicer gene deletion in the postnatal myocardium provokes spontaneous cardiac remodeling. Circulation 2008, 118, 1567–1576. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.F.; Mandel, E.M.; Thomson, J.M.; Wu, Q.; Callis, T.E.; Hammond, S.M.; Conlon, F.L.; Wang, D.Z. The role of microRNA-1 and microRNA-133 in skeletal muscle proliferation and differentiation. Nat. Genet. 2006, 38, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Wystub, K.; Besser, J.; Bachmann, A.; Boettger, T.; Braun, T. miR-1/133a clusters cooperatively specify the cardiomyogenic lineage by adjustment of myocardin levels during embryonic heart development. PLoS Genet. 2013, 9, e1003793. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Samal, E.; Srivastava, D. Serum response factor regulates a muscle-specific microRNA that targets Hand2 during cardiogenesis. Nature 2005, 436, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Ransom, J.F.; Li, A.; Vedantham, V.; von Drehle, M.; Muth, A.N.; Tsuchihashi, T.; McManus, M.T.; Schwartz, R.J.; Srivastava, D. Dysregulation of cardiogenesis, cardiac conduction, and cell cycle in mice lacking miRNA-1–2. Cell 2007, 129, 303–317. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, S.; He, A.; Kong, S.W.; Lu, J.; Bejar, R.; Bodyak, N.; Lee, K.H.; Ma, Q.; Kang, P.M.; Golub, T.R.; et al. MicroRNA-1 negatively regulates expression of the hypertrophy-associated calmodulin and Mef2a genes. Mol. Cell. Biol. 2009, 29, 2193–2204. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Bezprozvannaya, S.; Williams, A.H.; Qi, X.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. MicroRNA-133a regulates cardiomyocyte proliferation and suppresses smooth muscle gene expression in the heart. Genes Dev. 2008, 22, 3242–3254. [Google Scholar] [CrossRef] [PubMed]
- Biyashev, D.; Veliceasa, D.; Topczewski, J.; Topczewska, J.M.; Mizgirev, I.; Vinokour, E.; Reddi, A.L.; Licht, J.D.; Revskoy, S.Y.; Volpert, O.V. miR-27b controls venous specification and tip cell fate. Blood 2012, 119, 2679–2687. [Google Scholar] [CrossRef] [PubMed]
- Chinchilla, A.; Lozano, E.; Daimi, H.; Esteban, F.J.; Crist, C.; Aranega, A.E.; Franco, D. MicroRNA profiling during mouse ventricular maturation: A role for miR-27 modulating Mef2c expression. Cardiovasc. Res. 2011, 89, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.R.; Wu, M.; Yu, H.; Long, S.; Stevens, A.; Engers, D.W.; Sackin, H.; Daniels, J.S.; Dawson, E.S.; Hopkins, C.R.; et al. Selective inhibition of the Kir2 family of inward rectifier potassium channels by a small molecule probe: The discovery, SAR, and pharmacological characterization of ML133. ACS Chem. Biol. 2011, 6, 845–856. [Google Scholar] [CrossRef] [PubMed]
- Callis, T.E.; Pandya, K.; Seok, H.Y.; Tang, R.H.; Tatsuguchi, M.; Huang, Z.P.; Chen, J.F.; Deng, Z.; Gunn, B.; Shumate, J.; et al. MicroRNA-208a is a regulator of cardiac hypertrophy and conduction in mice. J. Clin. Investig. 2009, 119, 2772–2786. [Google Scholar] [CrossRef] [PubMed]
- Wilson, K.D.; Hu, S.; Venkatasubrahmanyam, S.; Fu, J.D.; Sun, N.; Abilez, O.J.; Baugh, J.J.; Jia, F.; Ghosh, Z.; Li, R.A.; et al. Dynamic microRNA expression programs during cardiac differentiation of human embryonic stem cells: Role for miR-499. Circ. Cardiovasc. Genet. 2010, 3, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Sluijter, J.P.; van Mil, A.; van Vliet, P.; Metz, C.H.; Liu, J.; Doevendans, P.A.; Goumans, M.J. MicroRNA-1 and -499 regulate differentiation and proliferation in human-derived cardiomyocyte progenitor cells. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Porrello, E.R.; Johnson, B.A.; Aurora, A.B.; Simpson, E.; Nam, Y.J.; Matkovich, S.J.; Dorn, G.W., 2nd; van Rooij, E.; Olson, E.N. miR-15 family regulates postnatal mitotic arrest of cardiomyocytes. Circ. Res. 2011, 109, 670–679. [Google Scholar] [CrossRef] [PubMed]
- Porrello, E.R.; Mahmoud, A.I.; Simpson, E.; Johnson, B.A.; Grinsfelder, D.; Canseco, D.; Mammen, P.P.; Rothermel, B.A.; Olson, E.N.; Sadek, H.A. Regulation of neonatal and adult mammalian heart regeneration by the miR-15 family. Proc. Natl. Acad. Sci. USA 2013, 110, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Kuhnert, F.; Mancuso, M.R.; Hampton, J.; Stankunas, K.; Asano, T.; Chen, C.Z.; Kuo, C.J. Attribution of vascular phenotypes of the murine Egfl7 locus to the microRNA miR-126. Development 2008, 135, 3989–3993. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Aurora, A.B.; Johnson, B.A.; Qi, X.; McAnally, J.; Hill, J.A.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. The endothelial-specific microRNA miR-126 governs vascular integrity and angiogenesis. Dev. Cell 2008, 15, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Fish, J.E.; Santoro, M.M.; Morton, S.U.; Yu, S.; Yeh, R.F.; Wythe, J.D.; Ivey, K.N.; Bruneau, B.G.; Stainier, D.Y.; Srivastava, D. miR-126 regulates angiogenic signaling and vascular integrity. Dev. Cell 2008, 15, 272–284. [Google Scholar] [CrossRef] [PubMed]
- Harris, T.A.; Yamakuchi, M.; Ferlito, M.; Mendell, J.T.; Lowenstein, C.J. MicroRNA-126 regulates endothelial expression of vascular cell adhesion molecule 1. Proc. Natl. Acad. Sci. USA 2008, 105, 1516–1521. [Google Scholar] [CrossRef] [PubMed]
- Doebele, C.; Bonauer, A.; Fischer, A.; Scholz, A.; Reiss, Y.; Urbich, C.; Hofmann, W.K.; Zeiher, A.M.; Dimmeler, S. Members of the microRNA-17-92 cluster exhibit a cell-intrinsic antiangiogenic function in endothelial cells. Blood 2010, 115, 4944–4950. [Google Scholar] [CrossRef] [PubMed]
- Mogilyansky, E.; Rigoutsos, I. The miR-17/92 cluster: A comprehensive update on its genomics, genetics, functions and increasingly important and numerous roles in health and disease. Cell Death Differ. 2013, 20, 1603–1614. [Google Scholar] [CrossRef] [PubMed]
- Bonauer, A.; Carmona, G.; Iwasaki, M.; Mione, M.; Koyanagi, M.; Fischer, A.; Burchfield, J.; Fox, H.; Doebele, C.; Ohtani, K.; et al. MicroRNA-92a controls angiogenesis and functional recovery of ischemic tissues in mice. Science 2009, 324, 1710–1713. [Google Scholar] [CrossRef] [PubMed]
- Santulli, G. MicroRNAs and Endothelial (Dys) Function. J. Cell. Physiol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Yancy, C.W.; Jessup, M.; Bozkurt, B.; Butler, J.; Casey, D.E., Jr.; Drazner, M.H.; Fonarow, G.C.; Geraci, S.A.; Horwich, T.; Januzzi, J.L.; et al. 2013 ACCF/AHA guideline for the management of heart failure: Executive summary: A report of the American College of Cardiology Foundation/American Heart Association Task Force on practice guidelines. Circulation 2013, 128, 1810–1852. [Google Scholar] [CrossRef] [PubMed]
- Alla, F.; Zannad, F.; Filippatos, G. Epidemiology of acute heart failure syndromes. Heart Fail. Rev. 2007, 12, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Stewart, S.; MacIntyre, K.; Hole, D.J.; Capewell, S.; McMurray, J.J. More “malignant” than cancer? Five-year survival following a first admission for heart failure. Eur. J. Heart Fail. 2001, 3, 315–322. [Google Scholar] [CrossRef]
- Heidenreich, P.A.; Albert, N.M.; Allen, L.A.; Bluemke, D.A.; Butler, J.; Fonarow, G.C.; Ikonomidis, J.S.; Khavjou, O.; Konstam, M.A.; Maddox, T.M.; et al. Forecasting the impact of heart failure in the United States: A policy statement from the American Heart Association. Circ. Heart Fail. 2013, 6, 606–619. [Google Scholar] [CrossRef] [PubMed]
- Goyal, A.; Norton, C.R.; Thomas, T.N.; Davis, R.L.; Butler, J.; Ashok, V.; Zhao, L.; Vaccarino, V.; Wilson, P.W. Predictors of incident heart failure in a large insured population: A one million person-year follow-up study. Circ. Heart Fail. 2010, 3, 698–705. [Google Scholar] [CrossRef] [PubMed]
- Lam, C.S.; Lyass, A.; Kraigher-Krainer, E.; Massaro, J.M.; Lee, D.S.; Ho, J.E.; Levy, D.; Redfield, M.M.; Pieske, B.M.; Benjamin, E.J.; et al. Cardiac dysfunction and noncardiac dysfunction as precursors of heart failure with reduced and preserved ejection fraction in the community. Circulation 2011, 124, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Roger, V.L. Epidemiology of heart failure. Circ. Res. 2013, 113, 646–659. [Google Scholar] [CrossRef] [PubMed]
- Owan, T.E.; Hodge, D.O.; Herges, R.M.; Jacobsen, S.J.; Roger, V.L.; Redfield, M.M. Trends in prevalence and outcome of heart failure with preserved ejection fraction. N. Engl. J. Med. 2006, 355, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, R.S.; Tu, J.V.; Lee, D.S.; Austin, P.C.; Fang, J.; Haouzi, A.; Gong, Y.; Liu, P.P. Outcome of heart failure with preserved ejection fraction in a population-based study. N. Engl. J. Med. 2006, 355, 260–269. [Google Scholar] [CrossRef] [PubMed]
- Hogg, K.; Swedberg, K.; McMurray, J. Heart failure with preserved left ventricular systolic function; epidemiology, clinical characteristics, and prognosis. J. Am. Coll. Cardiol. 2004, 43, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Schrier, R.W.; Abraham, W.T. Hormones and hemodynamics in heart failure. N. Engl. J. Med. 1999, 341, 577–585. [Google Scholar] [PubMed]
- Zois, N.E.; Bartels, E.D.; Hunter, I.; Kousholt, B.S.; Olsen, L.H.; Goetze, J.P. Natriuretic peptides in cardiometabolic regulation and disease. Nat. Rev. Cardiol. 2014, 11, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Samson, W.K. Adrenomedullin and the control of fluid and electrolyte homeostasis. Annu. Rev. Physiol. 1999, 61, 363–389. [Google Scholar] [CrossRef] [PubMed]
- Emeto, T.I.; Moxon, J.V.; Rush, C.; Woodward, L.; Golledge, J. Relevance of urocortins to cardiovascular disease. J. Mol. Cell. Cardiol. 2011, 51, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Fluck, D.C. Catecholamines. Br. Heart J. 1972, 34, 869–873. [Google Scholar] [CrossRef] [PubMed]
- Lefkowitz, R.J.; Rockman, H.A.; Koch, W.J. Catecholamines, cardiac beta-adrenergic receptors, and heart failure. Circulation 2000, 101, 1634–1637. [Google Scholar] [CrossRef] [PubMed]
- Spinarova, L.; Vitovec, J. Neurohumoral changes in chronic heart failure. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czech Repub. 2007, 151, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Attina, T.; Camidge, R.; Newby, D.E.; Webb, D.J. Endothelin antagonism in pulmonary hypertension, heart failure, and beyond. Heart 2005, 91, 825–831. [Google Scholar] [CrossRef] [PubMed]
- Potter, L.R.; Abbey-Hosch, S.; Dickey, D.M. Natriuretic peptides, their receptors, and cyclic guanosine monophosphate-dependent signaling functions. Endocr. Rev. 2006, 27, 47–72. [Google Scholar] [CrossRef] [PubMed]
- Rademaker, M.T.; Richards, A.M. Cardiac natriuretic peptides for cardiac health. Clin. Sci. (Lond.) 2005, 108, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Braunwald, E. Biomarkers in heart failure. N. Engl. J. Med. 2008, 358, 2148–2159. [Google Scholar] [CrossRef] [PubMed]
- Rademaker, M.T.; Charles, C.J.; Richards, A.M. Urocortin 1 administration from onset of rapid left ventricular pacing represses progression to overt heart failure. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H1536–H1544. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.E.; Pemberton, C.J.; Yandle, T.G.; Fisher, S.F.; Lainchbury, J.G.; Frampton, C.M.; Rademaker, M.T.; Richards, A.M. Urocortin 2 infusion in healthy humans: Hemodynamic, neurohormonal, and renal responses. J. Am. Coll. Cardiol. 2007, 49, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Nishikimi, T.; Miyata, A.; Horio, T.; Yoshihara, F.; Nagaya, N.; Takishita, S.; Yutani, C.; Matsuo, H.; Matsuoka, H.; Kangawa, K. Urocortin, a member of the corticotropin-releasing factor family, in normal and diseased heart. Am. J. Physiol. Heart Circ. Physiol. 2000, 279, H3031–H3039. [Google Scholar] [PubMed]
- McMurray, J.J.; Adamopoulos, S.; Anker, S.D.; Auricchio, A.; Bohm, M.; Dickstein, K.; Falk, V.; Filippatos, G.; Fonseca, C.; Gomez-Sanchez, M.A.; et al. ESC guidelines for the diagnosis and treatment of acute and chronic heart failure 2012: The Task Force for the Diagnosis and Treatment of Acute and Chronic Heart Failure 2012 of the European Society of Cardiology. Developed in collaboration with the Heart Failure Association (HFA) of the ESC. Eur. J. Heart Fail. 2012, 14, 803–869. [Google Scholar] [PubMed]
- Klip, I.T.; Voors, A.A.; Anker, S.D.; Hillege, H.L.; Struck, J.; Squire, I.; van Veldhuisen, D.J.; Dickstein, K.; investigators, O. Prognostic value of mid-regional pro-adrenomedullin in patients with heart failure after an acute myocardial infarction. Heart 2011, 97, 892–898. [Google Scholar] [CrossRef] [PubMed]
- Van Veldhuisen, D.J.; Linssen, G.C.; Jaarsma, T.; van Gilst, W.H.; Hoes, A.W.; Tijssen, J.G.; Paulus, W.J.; Voors, A.A.; Hillege, H.L. B-type natriuretic peptide and prognosis in heart failure patients with preserved and reduced ejection fraction. J. Am. Coll. Cardiol. 2013, 61, 1498–1506. [Google Scholar] [CrossRef] [PubMed]
- Lipshultz, S.E.; Rifai, N.; Sallan, S.E.; Lipsitz, S.R.; Dalton, V.; Sacks, D.B.; Ottlinger, M.E. Predictive value of cardiac troponin T in pediatric patients at risk for myocardial injury. Circulation 1997, 96, 2641–2648. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.E., 3rd; Bodor, G.S.; Davila-Roman, V.G.; Delmez, J.A.; Apple, F.S.; Ladenson, J.H.; Jaffe, A.S. Cardiac troponin I. A marker with high specificity for cardiac injury. Circulation 1993, 88, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Hayashida, K.; Kume, N.; Murase, T.; Minami, M.; Nakagawa, D.; Inada, T.; Tanaka, M.; Ueda, A.; Kominami, G.; Kambara, H.; et al. Serum soluble lectin-like oxidized low-density lipoprotein receptor-1 levels are elevated in acute coronary syndrome: A novel marker for early diagnosis. Circulation 2005, 112, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Semigran, M.; Shin, J.T. Heart Failure, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2013; p. 1. [Google Scholar]
- Gaggin, H.K.; Januzzi, J.L., Jr. Biomarkers and diagnostics in heart failure. Biochim. Biophys. Acta 2013, 1832, 2442–2450. [Google Scholar] [CrossRef] [PubMed]
- Troughton, R.W.; Frampton, C.M.; Brunner-La Rocca, H.P.; Pfisterer, M.; Eurlings, L.W.; Erntell, H.; Persson, H.; O’Connor, C.M.; Moertl, D.; Karlstrom, P.; et al. Effect of B-type natriuretic peptide-guided treatment of chronic heart failure on total mortality and hospitalization: An individual patient meta-analysis. Eur. Heart J. 2014, 35, 1559–1567. [Google Scholar] [CrossRef] [PubMed]
- Richards, A.M. The natriuretic peptides in heart failure. Basic Res. Cardiol. 2004, 99, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Januzzi, J.L.; van Kimmenade, R.; Lainchbury, J.; Bayes-Genis, A.; Ordonez-Llanos, J.; Santalo-Bel, M.; Pinto, Y.M.; Richards, M. NT-proBNP testing for diagnosis and short-term prognosis in acute destabilized heart failure: An international pooled analysis of 1256 patients: The International Collaborative of NT-proBNP Study. Eur. Heart J. 2006, 27, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Fuat, A.; Murphy, J.J.; Hungin, A.P.; Curry, J.; Mehrzad, A.A.; Hetherington, A.; Johnston, J.I.; Smellie, W.S.; Duffy, V.; Cawley, P. The diagnostic accuracy and utility of a B-type natriuretic peptide test in a community population of patients with suspected heart failure. Br. J. Gen. Pract. 2006, 56, 327–333. [Google Scholar] [PubMed]
- Christenson, R.H.; Azzazy, H.M.; Duh, S.H.; Maynard, S.; Seliger, S.L.; Defilippi, C.R. Impact of increased body mass index on accuracy of B-type natriuretic peptide (BNP) and N-terminal proBNP for diagnosis of decompensated heart failure and prediction of all-cause mortality. Clin. Chem. 2010, 56, 633–641. [Google Scholar] [CrossRef] [PubMed]
- Richards, M.; Di Somma, S.; Mueller, C.; Nowak, R.; Peacock, W.F.; Ponikowski, P.; Mockel, M.; Hogan, C.; Wu, A.H.; Clopton, P.; et al. Atrial fibrillation impairs the diagnostic performance of cardiac natriuretic peptides in dyspneic patients: Results from the BACH Study (Biomarkers in ACute Heart Failure). JACC Heart Fail. 2013, 1, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Rahimtoola, S.H. Digitalis therapy for patients in clinical heart failure. Circulation 2004, 109, 2942–2946. [Google Scholar] [CrossRef] [PubMed]
- Ambrosy, A.P.; Butler, J.; Ahmed, A.; Vaduganathan, M.; van Veldhuisen, D.J.; Colucci, W.S.; Gheorghiade, M. The use of digoxin in patients with worsening chronic heart failure: Reconsidering an old drug to reduce hospital admissions. J. Am. Coll. Cardiol. 2014, 63, 1823–1832. [Google Scholar] [CrossRef] [PubMed]
- Yusuf, S.; Pfeffer, M.A.; Swedberg, K.; Granger, C.B.; Held, P.; McMurray, J.J.; Michelson, E.L.; Olofsson, B.; Ostergren, J.; Investigators, C. Committees, Effects of candesartan in patients with chronic heart failure and preserved left-ventricular ejection fraction: The CHARM-Preserved Trial. Lancet 2003, 362, 777–781. [Google Scholar] [CrossRef]
- Massie, B.M.; Carson, P.E.; McMurray, J.J.; Komajda, M.; McKelvie, R.; Zile, M.R.; Anderson, S.; Donovan, M.; Iverson, E.; Staiger, C.; et al. Irbesartan in patients with heart failure and preserved ejection fraction. N. Engl. J. Med. 2008, 359, 2456–2467. [Google Scholar] [CrossRef] [PubMed]
- Capuano, A.; Scavone, C.; Vitale, C.; Sportiello, L.; Rossi, F.; Rosano, G.M.; Coats, A.J. Mineralocorticoid receptor antagonists in heart failure with preserved ejection fraction (HFpEF). Int. J. Cardiol. 2015, 200, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Pitt, B.; Pfeffer, M.A.; Assmann, S.F.; Boineau, R.; Anand, I.S.; Claggett, B.; Clausell, N.; Desai, A.S.; Diaz, R.; Fleg, J.L.; et al. Spironolactone for heart failure with preserved ejection fraction. N. Engl. J. Med. 2014, 370, 1383–1392. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, M.A.; Claggett, B.; Assmann, S.F.; Boineau, R.; Anand, I.S.; Clausell, N.; Desai, A.S.; Diaz, R.; Fleg, J.L.; Gordeev, I.; et al. Regional variation in patients and outcomes in the Treatment of Preserved Cardiac Function Heart Failure With an Aldosterone Antagonist (TOPCAT) trial. Circulation 2015, 131, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, S.L.; Kober, L.; Jhund, P.S.; Solomon, S.D.; Kjekshus, J.; McKelvie, R.S.; Zile, M.R.; Granger, C.B.; Wikstrand, J.; Komajda, M.; et al. International geographic variation in event rates in trials of heart failure with preserved and reduced ejection fraction. Circulation 2015, 131, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Rossignol, P.; Zannad, F. Regional differences in heart failure with preserved ejection fraction trials: When nephrology meets cardiology but east does not meet west. Circulation 2015, 131, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Richards, A.M. Therapeutic potential of infused cardiac natriuretic peptides in myocardial infarction. Heart 2009, 95, 1299–1300. [Google Scholar] [CrossRef] [PubMed]
- Nishikimi, T.; Maeda, N.; Matsuoka, H. The role of natriuretic peptides in cardioprotection. Cardiovasc. Res. 2006, 69, 318–328. [Google Scholar] [CrossRef] [PubMed]
- Filippatos, G.; Farmakis, D.; Parissis, J.; Lekakis, J. Drug therapy for patients with systolic heart failure after the PARADIGM-HF trial: In need of a new paradigm of LCZ696 implementation in clinical practice. BMC Med. 2015, 13, 35. [Google Scholar] [CrossRef] [PubMed]
- Bayes-Genis, A.; Barallat, J.; Galan, A.; de Antonio, M.; Domingo, M.; Zamora, E.; Urrutia, A.; Lupon, J. Soluble neprilysin is predictive of cardiovascular death and heart failure hospitalization in heart failure patients. J. Am. Coll. Cardiol. 2015, 65, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Minguet, J.; Sutton, G.; Ferrero, C.; Gomez, T.; Bramlage, P. LCZ696: A new paradigm for the treatment of heart failure? Expert Opin. Pharmacother. 2015, 16, 435–446. [Google Scholar] [PubMed]
- Von Lueder, T.G.; Krum, H. New medical therapies for heart failure. Nat. Rev. Cardiol. 2015, 12, 730–740. [Google Scholar] [CrossRef] [PubMed]
- Aronson, D.; Krum, H. Novel therapies in acute and chronic heart failure. Pharmacol. Ther. 2012, 135, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Mitrovic, V.; Seferovic, P.M.; Simeunovic, D.; Ristic, A.D.; Miric, M.; Moiseyev, V.S.; Kobalava, Z.; Nitsche, K.; Forssmann, W.G.; Luss, H.; et al. Haemodynamic and clinical effects of ularitide in decompensated heart failure. Eur. Heart J. 2006, 27, 2823–2832. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.Y.; Chen, H.H.; Lisy, O.; Swan, S.; Cannon, C.; Lieu, H.D.; Burnett, J.C., Jr. Pharmacodynamics of a novel designer natriuretic peptide, CD-NP, in a first-in-human clinical trial in healthy subjects. J. Clin. Pharmacol. 2009, 49, 668–673. [Google Scholar] [CrossRef] [PubMed]
- Lapp, H.; Mitrovic, V.; Franz, N.; Heuer, H.; Buerke, M.; Wolfertz, J.; Mueck, W.; Unger, S.; Wensing, G.; Frey, R. Cinaciguat (BAY 58-2667) improves cardiopulmonary hemodynamics in patients with acute decompensated heart failure. Circulation 2009, 119, 2781–2788. [Google Scholar] [CrossRef] [PubMed]
- Ponikowski, P.; Mitrovic, V.; Ruda, M.; Fernandez, A.; Voors, A.A.; Vishnevsky, A.; Cotter, G.; Milo, O.; Laessing, U.; Zhang, Y.; et al. A randomized, double-blind, placebo-controlled, multicentre study to assess haemodynamic effects of serelaxin in patients with acute heart failure. Eur. Heart J. 2014, 35, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.Y.; Frampton, C.M.; Crozier, I.G.; Troughton, R.W.; Richards, A.M. Urocortin-2 infusion in acute decompensated heart failure: Findings from the UNICORN study (urocortin-2 in the treatment of acute heart failure as an adjunct over conventional therapy). JACC Heart Fail. 2013, 1, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Rademaker, M.T.; Charles, C.J.; Ellmers, L.J.; Lewis, L.K.; Nicholls, M.G.; Richards, A.M. Prolonged urocortin 2 administration in experimental heart failure: Sustained hemodynamic, endocrine, and renal effects. Hypertension 2011, 57, 1136–1144. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.E.; Pemberton, C.J.; Yandle, T.G.; Fisher, S.F.; Lainchbury, J.G.; Frampton, C.M.; Rademaker, M.T.; Richards, M. Urocortin 2 infusion in human heart failure. Eur. Heart J. 2007, 28, 2589–2597. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Gu, X.; Li, Z.; Li, X.; Li, H.; Chang, J.; Chen, P.; Jin, J.; Xi, B.; Chen, D.; et al. Neuregulin-1/erbB-activation improves cardiac function and survival in models of ischemic, dilated, and viral cardiomyopathy. J. Am. Coll. Cardiol. 2006, 48, 1438–1447. [Google Scholar] [CrossRef] [PubMed]
- Tarigopula, M.; Davis, R.T., 3rd; Mungai, P.T.; Ryba, D.M.; Wieczorek, D.F.; Cowan, C.L.; Violin, J.D.; Wolska, B.M.; Solaro, R.J. Cardiac myosin light chain phosphorylation and inotropic effects of a biased ligand, TRV120023, in a dilated cardiomyopathy model. Cardiovasc. Res. 2015, 107, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Boerrigter, G.; Soergel, D.G.; Violin, J.D.; Lark, M.W.; Burnett, J.C., Jr. TRV120027, a novel β-arrestin biased ligand at the angiotensin II type I receptor, unloads the heart and maintains renal function when added to furosemide in experimental heart failure. Circ. Heart Fail. 2012, 5, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Sabbah, H.N.; Gupta, R.C.; Kohli, S.; Wang, M.; Rastogi, S.; Zhang, K.; Zimmermann, K.; Diedrichs, N.; Albrecht-Kupper, B.E. Chronic therapy with a partial adenosine A1-receptor agonist improves left ventricular function and remodeling in dogs with advanced heart failure. Circ. Heart Fail. 2013, 6, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Dawson, K.; Wakili, R.; Ordog, B.; Clauss, S.; Chen, Y.; Iwasaki, Y.; Voigt, N.; Qi, X.Y.; Sinner, M.F.; Dobrev, D.; et al. MicroRNA29: A mechanistic contributor and potential biomarker in atrial fibrillation. Circulation 2013, 127, 1466–1475, 1475e1–28. [Google Scholar] [CrossRef] [PubMed]
- Eitel, I.; Adams, V.; Dieterich, P.; Fuernau, G.; de Waha, S.; Desch, S.; Schuler, G.; Thiele, H. Relation of circulating microRNA-133a concentrations with myocardial damage and clinical prognosis in ST-elevation myocardial infarction. Am. Heart J. 2012, 164, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Watson, C.J.; Gupta, S.K.; O’Connell, E.; Thum, S.; Glezeva, N.; Fendrich, J.; Gallagher, J.; Ledwidge, M.; Grote-Levi, L.; McDonald, K.; et al. MicroRNA signatures differentiate preserved from reduced ejection fraction heart failure. Eur. J. Heart Fail. 2015, 17, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Wong, L.L.; Armugam, A.; Sepramaniam, S.; Karolina, D.S.; Lim, K.Y.; Lim, J.Y.; Chong, J.P.; Ng, J.Y.; Chen, Y.T.; Chan, M.M.; et al. Circulating microRNAs in heart failure with reduced and preserved left ventricular ejection fraction. Eur. J. Heart Fail. 2015, 17, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Ellis, K.L.; Cameron, V.A.; Troughton, R.W.; Frampton, C.M.; Ellmers, L.J.; Richards, A.M. Circulating microRNAs as candidate markers to distinguish heart failure in breathless patients. Eur. J. Heart Fail. 2013, 15, 1138–1147. [Google Scholar] [CrossRef] [PubMed]
- Tijsen, A.J.; Creemers, E.E.; Moerland, P.D.; de Windt, L.J.; van der Wal, A.C.; Kok, W.E.; Pinto, Y.M. MiR423-5p as a circulating biomarker for heart failure. Circ. Res. 2010, 106, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
- Endo, K.; Naito, Y.; Ji, X.; Nakanishi, M.; Noguchi, T.; Goto, Y.; Nonogi, H.; Ma, X.; Weng, H.; Hirokawa, G.; et al. MicroRNA 210 as a biomarker for congestive heart failure. Biol. Pharm. Bull. 2013, 36, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Niu, H.; Ban, T.; Xu, L.; Li, Y.; Wang, N.; Sun, L.; Ai, J.; Yang, B. Elevated plasma microRNA-1 predicts heart failure after acute myocardial infarction. Int. J. Cardiol. 2013, 166, 259–260. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, Y.; Nakanishi, M.; Nonogi, H.; Goto, Y.; Iwai, N. Assessment of plasma miRNAs in congestive heart failure. Circ. J. 2011, 75, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Corsten, M.F.; Dennert, R.; Jochems, S.; Kuznetsova, T.; Devaux, Y.; Hofstra, L.; Wagner, D.R.; Staessen, J.A.; Heymans, S.; Schroen, B. Circulating microRNA-208b and microRNA-499 reflect myocardial damage in cardiovascular disease. Circ. Cardiovasc. Genet. 2010, 3, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, S.; Sakata, Y.; Suna, S.; Nakatani, D.; Usami, M.; Hara, M.; Kitamura, T.; Hamasaki, T.; Nanto, S.; Kawahara, Y.; et al. Circulating p53-responsive microRNAs are predictive indicators of heart failure after acute myocardial infarction. Circ. Res. 2013, 113, 322–326. [Google Scholar] [CrossRef] [PubMed]
- Goren, Y.; Kushnir, M.; Zafrir, B.; Tabak, S.; Lewis, B.S.; Amir, O. Serum levels of microRNAs in patients with heart failure. Eur. J. Heart Fail. 2012, 14, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Jing, Z.C.; Ellinor, P.T.; Liang, D.; Zhang, H.; Liu, Y.; Chen, X.; Pan, L.; Lyon, R.; Liu, Y.; et al. MicroRNA-134 as a potential plasma biomarker for the diagnosis of acute pulmonary embolism. J. Transl. Med. 2011, 9, 159. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.S.; Chen, Y.; Jiang, H.; Lu, J.P.; Zhang, G.; Geng, J.; Zhang, Q.; Shen, J.H.; Zhou, X.; Zhu, W.; et al. Serum miR-210 and miR-30a expressions tend to revert to fetal levels in Chinese adult patients with chronic heart failure. Cardiovasc. Pathol. Off. J. Soc. Cardiovasc. Pathol. 2013, 22, 444–450. [Google Scholar] [CrossRef] [PubMed]
- Goren, Y.; Meiri, E.; Hogan, C.; Mitchell, H.; Lebanony, D.; Salman, N.; Schliamser, J.E.; Amir, O. Relation of reduced expression of miR-150 in platelets to atrial fibrillation in patients with chronic systolic heart failure. Am. J. Cardiol. 2014, 113, 976–981. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.J.; Han, M.; Yang, F.Y.; Wei, G.C.; Liang, Z.G.; Yao, H.; Ji, C.W.; Xie, R.S.; Gong, C.L.; Tian, Y. Biological significance of miR-126 expression in atrial fibrillation and heart failure. Braz. J. Med. Biol. Res. 2015, 48, 983–989. [Google Scholar] [CrossRef] [PubMed]
- Lai, K.B.; Sanderson, J.E.; Izzat, M.B.; Yu, C.M. Micro-RNA and mRNA myocardial tissue expression in biopsy specimen from patients with heart failure. Int. J. Cardiol. 2015, 199, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Qiang, L.; Hong, L.; Ningfu, W.; Huaihong, C.; Jing, W. Expression of miR-126 and miR-508-5p in endothelial progenitor cells is associated with the prognosis of chronic heart failure patients. Int. J. Cardiol. 2013, 168, 2082–2088. [Google Scholar] [CrossRef] [PubMed]
- Gidlof, O.; Smith, J.G.; Miyazu, K.; Gilje, P.; Spencer, A.; Blomquist, S.; Erlinge, D. Circulating cardio-enriched microRNAs are associated with long-term prognosis following myocardial infarction. BMC Cardiovasc. Disord. 2013, 13, 12. [Google Scholar] [CrossRef] [PubMed]
- Nair, N.; Kumar, S.; Gongora, E.; Gupta, S. Circulating miRNA as novel markers for diastolic dysfunction. Mol. Cell. Biochem. 2013, 376, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Voellenkle, C.; van Rooij, J.; Cappuzzello, C.; Greco, S.; Arcelli, D.; Di Vito, L.; Melillo, G.; Rigolini, R.; Costa, E.; Crea, F.; et al. MicroRNA signatures in peripheral blood mononuclear cells of chronic heart failure patients. Physiol. Genom. 2010, 42, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Naga Prasad, S.V.; Duan, Z.H.; Gupta, M.K.; Surampudi, V.S.; Volinia, S.; Calin, G.A.; Liu, C.G.; Kotwal, A.; Moravec, C.S.; Starling, R.C.; et al. Unique microRNA profile in end-stage heart failure indicates alterations in specific cardiovascular signaling networks. J. Biol. Chem. 2009, 284, 27487–27499. [Google Scholar] [CrossRef] [PubMed]
- Sucharov, C.; Bristow, M.R.; Port, J.D. miRNA expression in the failing human heart: Functional correlates. J. Mol. Cell. Cardiol. 2008, 45, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Duan, Q.; Chen, C.; Yang, L.; Li, N.; Gong, W.; Li, S.; Wang, D.W. MicroRNA regulation of unfolded protein response transcription factor XBP1 in the progression of cardiac hypertrophy and heart failure in vivo. J. Transl. Med. 2015, 13, 363. [Google Scholar] [CrossRef] [PubMed]
- Da Costa Martins, P.A.; De Windt, L.J. MicroRNAs in control of cardiac hypertrophy. Cardiovasc. Res. 2012, 93, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Sun, X.; Ren, J.; Li, X.; Gao, X.; Lu, C.; Zhang, Y.; Sun, H.; Wang, Y.; Wang, H.; et al. miR-1 exacerbates cardiac ischemia-reperfusion injury in mouse models. PLoS ONE 2012, 7, e50515. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Zhong, Y.; Cheng, C.; Liu, B.; Wang, L.; Li, A.; Xiong, L.; Liu, S. miR-30-regulated autophagy mediates angiotensin II-induced myocardial hypertrophy. PLoS ONE 2013, 8, e53950. [Google Scholar] [CrossRef] [PubMed]
- Thum, T.; Gross, C.; Fiedler, J.; Fischer, T.; Kissler, S.; Bussen, M.; Galuppo, P.; Just, S.; Rottbauer, W.; Frantz, S.; et al. MicroRNA-21 contributes to myocardial disease by stimulating MAP kinase signalling in fibroblasts. Nature 2008, 456, 980–984. [Google Scholar] [CrossRef] [PubMed]
- Sayed, D.; He, M.; Hong, C.; Gao, S.; Rane, S.; Yang, Z.; Abdellatif, M. MicroRNA-21 is a downstream effector of AKT that mediates its antiapoptotic effects via suppression of Fas ligand. J. Biol. Chem. 2010, 285, 20281–20290. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Yang, Y.; Wang, Y.; Li, J.; Schiller, P.W.; Peng, T. MicroRNA-195 promotes palmitate-induced apoptosis in cardiomyocytes by down-regulating Sirt1. Cardiovasc. Res. 2011, 92, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jia, Z.; Zhang, C.; Sun, M.; Wang, W.; Chen, P.; Ma, K.; Zhang, Y.; Li, X.; Zhou, C. miR-499 protects cardiomyocytes from H2O2-induced apoptosis via its effects on Pdcd4 and Pacs2. RNA Biol. 2014, 11, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zhou, M.; Li, C.; Zhou, J.; Li, H.; Zhu, D.; Wang, Z.; Chen, A.; Zhao, Q. MicroRNA-92a inhibition attenuates hypoxia/reoxygenation-induced myocardiocyte apoptosis by targeting Smad7. PLoS ONE 2014, 9, e100298. [Google Scholar] [CrossRef] [PubMed]
- Wong, L.L.; Wee, A.S.; Lim, J.Y.; Ng, J.Y.; Chong, J.P.; Liew, O.W.; Lilyanna, S.; Martinez, E.C.; Ackers-Johnson, M.A.; Vardy, L.A.; et al. Natriuretic peptide receptor 3 (NPR3) is regulated by microRNA-100. J. Mol. Cell. Cardiol. 2015, 82, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Yang, P.; Xiong, Q.; Song, X.; Yang, X.; Liu, L.; Yuan, W.; Rui, Y.C. MicroRNA-125a/b-5p inhibits endothelin-1 expression in vascular endothelial cells. J. Hypertens. 2010, 28, 1646–1654. [Google Scholar] [CrossRef] [PubMed]
- Eskildsen, T.V.; Schneider, M.; Sandberg, M.B.; Skov, V.; Bronnum, H.; Thomassen, M.; Kruse, T.A.; Andersen, D.C.; Sheikh, S.P. The microRNA-132/212 family fine-tunes multiple targets in Angiotensin II signalling in cardiac fibroblasts. J. Renin Angiotensin Aldosterone Syst. 2015, 16, 1288–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotlo, K.U.; Hesabi, B.; Danziger, R.S. Implication of microRNAs in atrial natriuretic peptide and nitric oxide signaling in vascular smooth muscle cells. Am. J. Physiol. Cell Physiol. 2011, 301, C929–C937. [Google Scholar] [CrossRef] [PubMed]
- Rane, S.; He, M.; Sayed, D.; Yan, L.; Vatner, D.; Abdellatif, M. An antagonism between the AKT and β-adrenergic signaling pathways mediated through their reciprocal effects on miR-199a-5p. Cell Signal. 2010, 22, 1054–1062. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.M.; Lee, E.J.; Buckenberger, J.A.; Schmittgen, T.D.; Elton, T.S. MicroRNA-155 regulates human angiotensin II type 1 receptor expression in fibroblasts. J. Biol. Chem. 2006, 281, 18277–18284. [Google Scholar] [CrossRef] [PubMed]
- Arora, P.; Wu, C.; Khan, A.M.; Bloch, D.B.; Davis-Dusenbery, B.N.; Ghorbani, A.; Spagnolli, E.; Martinez, A.; Ryan, A.; Tainsh, L.T.; et al. Atrial natriuretic peptide is negatively regulated by microRNA-425. J. Clin. Investig. 2013, 123, 3378–3382. [Google Scholar] [CrossRef] [PubMed]
- Maharjan, S.; Mopidevi, B.; Kaw, M.K.; Puri, N.; Kumar, A. Human aldosterone synthase gene polymorphism promotes miRNA binding and regulates gene expression. Physiol. Genom. 2014, 46, 860–865. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. Elife 2015, 4, e05005. [Google Scholar] [CrossRef] [PubMed]
- TargetScan Human Prediction of MicroRNA Targets. Available online: http://www.targetscan.org/ (acceseed on 3 December 2015).
- Wong, N.; Wang, X. MiRDB: An online resource for microRNA target prediction and functional annotations. Nucleic Acids Res. 2015, 43, D146–D152. [Google Scholar] [CrossRef] [PubMed]
- MiRDB. Available online: http://mirdb.org/miRDB/ (accessed on 3 December 2015).
- Betel, D.; Koppal, A.; Agius, P.; Sander, C.; Leslie, C. Comprehensive modeling of microRNA targets predicts functional non-conserved and non-canonical sites. Genome Biol. 2010, 11, R90. [Google Scholar] [CrossRef] [PubMed]
- MicroRNA.org-Targets and Expression. Available online: http://www.microrna.org/microrna/home.do (accessed on 3 December 2015).
- Brennecke, J.; Stark, A.; Russell, R.B.; Cohen, S.M. Principles of microRNA-target recognition. PLoS Biol. 2005, 3, e85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, D.; Liu, H.; Huang, Y. Survey of Computational Algorithms for MicroRNA Target Prediction. Curr. Genom. 2009, 10, 478–492. [Google Scholar] [CrossRef] [PubMed]
- Janssen, H.L.; Reesink, H.W.; Lawitz, E.J.; Zeuzem, S.; Rodriguez-Torres, M.; Patel, K.; van der Meer, A.J.; Patick, A.K.; Chen, A.; Zhou, Y.; et al. Treatment of HCV infection by targeting microRNA. N. Engl. J. Med. 2013, 368, 1685–1694. [Google Scholar] [CrossRef] [PubMed]
- Bader, A.G. miR-34—A microRNA replacement therapy is headed to the clinic. Front. Genet. 2012, 3, 120. [Google Scholar] [CrossRef] [PubMed]
- Santaris Pharma A/S. Miravirsen Study in Null Responder to Pegylated Interferon Alpha Plus Ribavirin Subjects With Chronic Hepatitis C. Available online: https://clinicaltrials.gov/show/NCT01727934 (accessed on 3 December 2015).
- Cloonan, N.; Wani, S.; Xu, Q.; Gu, J.; Lea, K.; Heater, S.; Barbacioru, C.; Steptoe, A.L.; Martin, H.C.; Nourbakhsh, E.; et al. MicroRNAs and their isomiRs function cooperatively to target common biological pathways. Genome Biol. 2011, 12, R126. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, Z.; Gemeinhart, R.A. Progress in microRNA delivery. J. Control. Release 2013, 172, 962–974. [Google Scholar] [CrossRef] [PubMed]
- Ko, Y.T.; Hartner, W.C.; Kale, A.; Torchilin, V.P. Gene delivery into ischemic myocardium by double-targeted lipoplexes with anti-myosin antibody and TAT peptide. Gene Ther. 2009, 16, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Kwekkeboom, R.F.; Lei, Z.; Doevendans, P.A.; Musters, R.J.; Sluijter, J.P. Targeted delivery of miRNA therapeutics for cardiovascular diseases: Opportunities and challenges. Clin. Sci. (Lond.) 2014, 127, 351–365. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Established Risk Factors | Increased HF Risk | Life Style Factors |
|---|---|---|
| Coronary artery disease | Higher serum creatinine | Obesity |
| Hypertension | Lower FEV1:FVC ratios | Smoking |
| Diabetes mellitus | Lower hemogloblin concentrations | Lack of exercise |
| Atrial fibrillation | Excessive alcohol consumption | |
| Valvular heart disease | ||
| Dilated cardiomyopathy * |
| Neurohormone | Receptor | Cardiovascular Functions | Pathological Effects |
|---|---|---|---|
| Catecholamines epinephrine and norepinephrine [75,76] | α-AR β-AR | Activation of α-AR produces vasoconstriction effect. Activation of β-AR procures myocardial contraction (both inotropic and chronotropic), and vasodilation effects. | Arrhythmias, cardiomyopathy, and sudden death |
| Renin | Renin cleaves angiotensinogen into angiotensin I. | Same as Angiotensin II | |
| Angiotensin II [71,77] | AGTR1, AGTR2 | Angiotensin I then converted by angiotensin-converting enzyme to angiotensin II. Activation of AGTRs produces vasoconstriction effect, it also stimulates the SNS and increases the secretion of aldosterone and subsequently leads to production of arginine vasopressin. | Hypertrophy of the myocardium and cardiac remodeling |
| Aldosterone [77] | NR3C2 | Activation of NR3C2 leads to sodium retention, potassium excretion and increase blood pressure. | Cardiac fibrosis and remodeling |
| Arginine vasopressin [71,77] | V1R V2R | Activation of V1R leads to vasoconstriction effect. Stimulation of V2R leads to retention of water, and antidiuretic effect. | Hyponatremia and antidiuresis |
| Endothelin [71,77,78] | EDNRA EDNRB | Activation of EDNRA causes vasoconstriction, while activation of EDNRB leads to vasodilation. | Hypertrophy. Systemic and renal vasoconstriction |
| ANP [79,80] | NPR1 NPR3 | Activation of NPR1 leads to vasodilation, diuresis, natriuresis. It also suppresses RAAS, SNS, and have an anti-hypertrophic effect. | Hypotension |
| BNP [79,80] | NPR1 NPR3 | Activation of NPR1 leads to vasodilation, diuresis, natriuresis. It also suppresses RAAS, SNS, and have an anti-hypertrophic effect. | Hypotension |
| CNP [79] | NPR2 NPR3 | Activation of NPR1 leads to vasodilation, diuresis, natriuresis and have an anti-proliferative effect. | |
| ADM [5,81] | GPR-182 | Vasodilatation with inotropism and natriuresis | |
| Urocortins [82,83,84] | CRHRs | Positive inotropic and chronotropic effects, arterial and venous dilatation |
| HF Signaling Cascades | Biomarkers for HF |
|---|---|
| Neurohormonal activation | Norepinephrine, Renin activity, Angiotensin, Aldosterone, Arginine-Vasopressin |
| Myocardial overload | BNP, NT-proBNP, MR-proANP, MR-proADM |
| Cardiac injury | Troponin T, Troponin I, LOX-1, GDF-15 |
| Cardiac remodeling | IL-6, TNFα, CRP, MMP, Galectin-3, Soluble ST2 |
| Study Cohort | NT-proBNP, LVEF and Other Criteria | Platform | miRNA Identified | Diagnostic Potential | Reference | |
|---|---|---|---|---|---|---|
| Discovery | Validation | |||||
| Whole blood and plasma, no-HF (n = 28), HFREF (n = 39) and HFPEF (n = 19) | Plasma from no-HF (n = 30), HFREF (n = 30) and HFPEF (n = 30) | NT-proBNP: 3086 ± 421 pg/mL; HFPEF: LVEF ≥ 50%, HFREF: LVEF ≤ 40% | miRNA microarray and RT-PCR | miR-1233, -183-3p, -190a, -193b-3p, -193b-5p, -211-5p, -494, and -671-5p | HF | Wong et al. [129] |
| miR-125a-5p, -183-3p, -193b-3p, -211-5p, -494, -638, and -671-5p | HFREF | |||||
| miR-1233, -183-3p, -190a, -193b-3p, -193b-5p, and -545-5p | HFPEF | |||||
| miR-125a-5p, -190a, -550a-5p, and -638 | HFREF vs. HFPEF | |||||
| Whole blood, from control (n = 39) and HFREF (n = 53) | Serum from controls (n = 8) and HFREF (n = 14) | NT-proBNP: 2399 ± 3395 ng/L, HFREF: LVEF < 50% | miRNA microarray and RT-PCR | miR-200b-5p, miR-622, miR-1228-5p | HFREF | Vogel et al. [2] |
| Study Cohort | NT-proBNP, LVEF and Other Criteria | Platform | miRNA Identified | Diagnostic Potential | Reference | |
|---|---|---|---|---|---|---|
| Discovery | Validation | |||||
| Serum from control (n = 32), AF (n = 35), HF (n = 32), HF-AF (n = 36) | NYHA class III, IV, Log(NT-proBNP): 4.07 ± 0.51, LVEF: 48.32% ± 6.00% | RT-PCR | miR-126 | Severity of AF and HF | Wei et al. [141] | |
| Serum pooled from n = 15 per group in no-HF, HFREF and HFPEF | Serum from n = 75 per group in no-HF, HFREF and HFPEF | HFPEF: NYHA class IV, LVEF ≥ 50%, BNP: 215 (126–353) pg/mL, HFREF: LVEF < 50%, BNP: 139 (71–254) pg/mL | qPCR array, RT-PCR | miR-30c, miR-146a, miR-221, miR-328, miR-375 | HF and HFREF vs. HFPEF | Watson et al. [128] |
| Platelets and serum from control (n = 35), HF (n = 26) and AF-HF (n = 15) | HF: LVEF < 40%, NYHA class I-IV, BNP: 147 (47–416) pg/mL | RT-PCR | miR-150 | AF-HF | Goren et al. [140] | |
| Serum from control (n = 7) and HF (n = 7) | Serum from control (n = 65) and HF (n = 21) | Patients with acute stage of AMI onset (Killip class > II) developed HF | qPCR array, RT-PCR | miR-192, miR-194, miR-34a | HF after AMI | Matsumoto et al. [136] |
| Serum from 2 pooled samples of control (n = 6) and HF (n = 6) | Serum from control (n = 30) and HF (n = 30) | Chronic stable class C HFREF with LVEF < 40%, BNP: 180 (98–276) pg/mL | qPCR array, RT-PCR | miR-423-5p, -320a, -22, -92b, -17, -532-3p, -92a, -30a, -21, -101 | HFREF | Goren et al. [137] |
| Serum from control (n = 18), HF (n = 22) | NYHA class III, IV, pro-BNP ≥ 1000 ng/L | RT-PCR | miR-210, miR-30a | HF | Zhao et al. [139] | |
| Study Cohort | NT-proBNP, LVEF and Other Criteria | Platform | miRNA Identified | Diagnostic Potential | Reference | |
|---|---|---|---|---|---|---|
| Discovery | Validation | |||||
| Plasma from AMI patients (n = 49) with various EF | AMI patient: cardiac troponin, creatine kinase-MB, Q-waves and ST-segment elevation | RT-PCR | miR-1 | HF after AMI | Zhang et al. [133] | |
| Plasma from control (n = 20) and HF (n = 33) | Framingham criteria, NT-pro-BNP > 200 pmol/L | RT-PCR | miR-499, miR-122 | Acute HF | Corsten et al. [135] | |
| Plasma from control (n = 12), HF (n = 12) | Plasma from control (n = 39), HF (n = 30) | Framingham criteria, NT-proBNP > 1000 ng/L | miRNA microarray RT-PCR | miR-423-5p, -18b-3p, -129-5p, -1254, -675, -622 | Acute HF | Tijsen et al. [131] |
| Plasma from ACS (n = 424) | Coronary artery bypass grafting patients and ACS patient with STEMI and NSTEMI | RT-PCR | miR-1, miR-208, miR-499-5p | HF after MI | Gidlöf et al. [144] | |
| Plasma from control (n = 14) and HF (n = 32) | Plasma from HF (n = 44) and control (n = 15) | Discovery cohort: HFREF: 27.3 ± 9.0, HFPEF: 57.8 ± 7.0, NT-proBNP:460.8 (141.3–2511.9) pmol/L, Validation cohort: HFREF:27.0 ± 7.7, HFPEF:62.0 ± 6.4, NT-proBNP: 493.28 (25.7–3801.9) pmol/L | qPCR array RT-PCR | miR-185, miR-103, miR-142-3p, miR-30b, miR-342-3p, miR-150 | Acute HF | Ellis et al. [130] |
| Plasma from HF (n = 8) and control (n = 3) | Control (n = 17), HF (NYHA II) (n = 17), NYHA III (n = 6) and NYHA IV (n = 10) | NYHA class II–IV | miRNA microarray RT-PCR | miR-126 | HF | Fukushima et al. [134] |
| Study Cohort | NT-proBNP, LVEF and Other Criteria | Platform | miRNA Identified | Diagnostic Potential | Reference | |
|---|---|---|---|---|---|---|
| Discovery | Validation | |||||
| Myocardial biopsy from control (n = 17) and HF (n = 17) | LVEF mean: 30%, HF due to myocarditis or DCM | RT-PCR | miR-1, -21, -23, -29, -130, -195, -199 | HF | Lai et al. [142] | |
| LV Tissue from non-failing (n = 10) and DCM (n = 30) | LV Tissue from non-failing (n = 10) and DCM (n = 20) | DCM with EF 15% ± 1% | miRNA microarray RT-PCR | miR-1, -29b, -7, -378, -214, -342, -145, -125b, -181b | HF | Naga Prasad et al. [147] |
| LV Tissue Non-failing (n = 6), IDCM (n = 5), Ischemic DCM (n = 5) | IDC and ISC patients | miRNA microarray RT-PCR | miR-100, miR-195, miR-92, miR-133b | HF | Sucharov et al. [148] | |
| Study Cohort | NT-proBNP, LVEF and Other Criteria | Platform | miRNA Identified | Diagnostic Potential | Reference | |
|---|---|---|---|---|---|---|
| Discovery | Validation | |||||
| Mononuclear from control (n = 6), NYHA II (n = 8) and NYHA III, IV (n = 5) | NYHA class II–IV | RT-PCR | miR-210 | HF | Endo et al. [132] | |
| Buffy coat HFPEF (n = 8), DCM (n = 10), DCM-CHF (n = 13), Control (n = 8) | HFPEF with mean LVEF 61.13 and mean BNP 353.99 pg/mL, DCM-HF with mean LVEF 19.23 and BNP 2247 pg/mL | miRNA microarray RT-PCR | miR-454, miR-500, miR-1246, | HFPEF | Nair et al. [145] | |
| miR-142-3p, miR-124-5p | DCM-HF | |||||
| PBMC from control (n = 9) and HF (n = 15) | PBMC from control (n = 19) and HF (n = 34) | NYHA class III/IV with mean LVEF ≤ 36% | RT-PCR | miR-139, miR-142-5p, miR-107 | Chronic HF | Voellenkle et al. [146] |
| EPC from control (n = 10), ICM-HF (n = 10) and NICM-HF (n = 10) | EPC from control (n = 30), ICM-HF (n = 55) and NICM-HF (n = 51) | NYHA class III, IV | qPCR array RT-PCR | miR-126, miR-508-5p | HF | Qiang et al. [143] |
| miRNA | Targetscan | miRDB | miRanda | ||
|---|---|---|---|---|---|
| Conserved | Poorly Conserved | Gene (Target Score *) | Good mirSVR Score and Conserved | Non-Good mirSVR Score and Conserved | |
| miR-1 | – | AGTR1 | – | AGTR1, EDNRB | AGT, ACE, EDN1, EDNRA |
| miR-100 | – | – | – | – | NPR3 |
| miR-101-3p | – | ACE | – | AGTR2, CALCRL, EDN1, EDNRB, NR3C2 | AGT, CALCRL, EDN1 |
| miR-103a-3p | CRHR2 | AGT, AGTR1, NPPA | – | REN | AGT, CRHR1, UCN2, NR3C2, NPR2, NPPA, EDNRA, EDN1, ATP6AP2, ACE, AGTR2, AGTR1 |
| miR-107 | CRHR1 | AGT, AGTR1, NPPA | – | REN | AGT, AGTR1, AGTR2,ACE, ATP6AP2, EDN1, EDNRA, NPPA, NPR2, NR3C2, UCN2, CRHR1 |
| miR-122 | – | – | – | ATP6AP2, EDN1, NPR3, CRHR1 | ACE, ATP6AP2, EDNRA, CRHR1, CRHR2, NR3C2, CYP11B2 |
| miR-1228-5p | – | – | – | – | – |
| miR-1233-3p | CRHR2 | – | – | – | – |
| miR-124-5p | – | – | – | AGTR1, EDNRB, NR3C2 | ACE, EDN1, NPR1, CYP11B2, NR3C2, CRHR1 |
| miR-1246 | – | – | – | – | – |
| miR-1254 | – | ACE, NPR1, CYP11B2 | NPR3(63) | – | – |
| miR-125a-5p | CRHR2 | ACE, CYP11B2 | – | NPR3, CYP11B2 | – |
| miR-125b-5p | CRHR1 | ACE, CYP11B2 | – | NPR3, CYP11B2 | AGTR2, ACE, EDN1, EDNRA |
| miR-126-3p | – | – | – | – | – |
| miR-129-5p | – | AGT, NPR1, NPR2 | NR3C2(84), AGTR1(76) | EDN1, EDNRA, EDNRB, NPR3, NR3C2 | ACE, CALCRL, ATP6AP2, EDN1, EDNRA, EDNRB, NPR2, NPR3, AGT |
| miR-130a-3p | – | NPR1 | EDN1(69) | ATP6AP2, EDN1, NR3C2 | AGT, ACE, EDN1, EDNRA, NR3C2, CRHR1 |
| miR-133b | – | – | ATP6AP2(54) | ATP6AP2, CRHR1 | |
| miR-139-5p | – | NPPA | – | CALCRL, EDNRB, NPPA, NPR3, NR3C2 | ACE, CALCRL, EDNRA, EDNRB, NR3C2 |
| miR-142-3p | – | - | – | CALCRL, NR3C2 | ACE, CALCRL, EDNRA, EDNRB, NR3C2 |
| miR-142-5p | AGTR2 | AGT, ACE | – | – | – |
| miR-145-5p | – | AGT | – | AGTR2, CALCRL | AGTR2, ACE, ATP6AP2, EDN1, EDNRB, CRHR1 |
| miR-146a-5p | – | CRHR2, NPR1, CRHR2 | – | CALCRL, EDNRB, NPR2, NPR3 | – |
| miR-150-5p | CRHR2 | GRP182, NPR1 | – | ATP6AP2, EDNRB, NPR3 | – |
| miR-17-5p | – | AGTR2, NPR1 | – | AGTR2, ACE, NPR3 | AGTR2, ACE, CALCRL, EDN1, EDNRA, EDNRB, NPR3, NR3C2 |
| miR-181b-5p | – | AGT, AGTR1 | ADM(74), CALCRL(56) | AGTR1, ADM, CALCRL, NPR3, ATP6AP2, EDNRB, NR3C2 | AGT, ACE, EDRNA |
| miR-183-3p | – | AGTR1 | – | – | – |
| miR-185-5p | CRHR2 | ACE, NPR1, CYP11B2 | – | CYP11B2 | – |
| miR-18b-3p | – | CYP11B2 | – | – | – |
| miR-190a | – | – | – | – | – |
| miR-192 | – | – | – | NPR3 | AGTR2, ACE, CALCRL, EDN1, UCN2, CRHR1 |
| miR193b-3p | – | AGT, CYP11B2, CRHR2 | – | EDN1 | – |
| miR-193b-5p | – | NPR1 | – | – | – |
| miR-194-5p | NPPA | – | EDN1(70) | EDN1, NPPA, NPR3 | – |
| miR-195-5p | – | AGT, CYP11B2, CRHR2 | – | AGTR2, NPR2, NPR3 | – |
| miR-199a-5p | – | ACE | – | AGTR2, DNRA, EDNRB, UCN2 | AGTR1, ACE, ATP6AP2, EDN1, EDNRA, CYP11B2, CRHR1, CRHR2 |
| miR-200b-5p | – | AGTR1 | – | – | – |
| miR-208a | AGTR2 | – | – | ATP6AB2, EDNRB | AGTR1, CALCRL, UCN2 |
| miR-21-5p | – | – | NPPB(69) | EDNRB | EDNRA, NPPA, NPPB |
| miR-210-5p | – | NPR1 | – | CRHR2 | NR3C2 |
| miR-211-5p | – | – | NR3C2(86) | CALCRL, ATP6AP2, EDNRA, NPR3, NR3C2, CRHR2 | ATP6AP2, EDN1, EDNRA, CRHR1, CRHR2 |
| miR-214-3p | – | ACE, REN | – | AGTR1, CALCRL, REN, EDN1, EDNRB, CRHR1 | EDN1, EDNRA, NPPA, NPR2, UCN2 |
| miR-22-3p | – | AGT | – | ACE, EDNRA, NPR3, CRHR1 | ACE, NPPA, NPR2, CYP11B2, NR3C2, CRHR1, CRHR2 |
| miR-221-3p | – | ACE | – | NPR3, NR3C2 | ACE, CALCRL, EDNRA, EDNRB, NPR2, NR3C2, CRHR1 |
| miR-23a-3p | – | NPR1 | NPR3(60) | AGTR2, CALCRL, EDNRB | ACE, ADM, CALCRL, EDN1, EDNRA, NR3C2 |
| miR-29a-3p | – | – | – | EDNRB, NPPA, NPR3 | AGTR1, ACE, EDNRB, CYP11B2, UCN2 |
| miR-29b-3p | – | – | – | EDNRB, NPPA, NPR3 | AGTR1, ACE, EDNRB, CYP11B2, UCN2 |
| miR-30a-5p | – | – | EDNRA(54) | EDN1, EDNRA, EDNRB, NPR3 | AGTR1, AGTR2, EDNRA, EDNRB, NR3C2 |
| miR-30b-5p | – | – | EDNRA(54) | AGTR1, EDN1, EDNRA, EDNRB, NPR3 | AGTR2, EDNRA, EDNRB, NR3C2 |
| miR-30c-5p | – | – | EDNRA(54) | AGTR1, EDN1, EDNRA, EDNRB, NPR3 | AGTR2, EDNRA, EDNRB, NR3C2 |
| miR-320a | – | NPPB | – | EDNRA, NPPB, NPR3, NR3C2, EDNRA, NPR3, NR3C2 | – |
| miR-328-3p | – | AGT, CYP11B2 | – | UCN2 | CRHR2 |
| miR-342-3p | – | – | – | UCN2, CRHR2 | – |
| miR-34a-5p | CRHR1 | NPR1 | UCN2(95), CRHR1(54) | AGTR1, EDNRB, NR3C2, UCN2, CRHR1 | AGT, ACE, CALCRL, EDN1, EDNRA, EDNRB |
| miR-375 | – | AGT, GTR1 | – | ATP6AP2 | AGT |
| miR-378a-5p | – | AGTR2, NPR1, CYP11B2 | – | EDN1, CYP11B2, CRHR1 | – |
| miR-423-5p | – | AGT, REN, CRHR2 | CRHR2(56) | – | – |
| miR-454 | – | NPR1 | – | ATP6AP2, EDN1, NPR3, NR3C2 | – |
| miR-494 | – | – | – | AGTR1, END1, EDNRA, EDNRB, NPR3 | – |
| miR-499-5p | – | – | – | CALCRL, ATP6AP2 | – |
| miR-500a-5p | – | AGTR2 | CALCRL(53) | – | – |
| miR-508-5p | – | ACE | – | – | NPR1 |
| miR-532-3p | – | GPR182, CRHR2 | NPR3(64) | – | – |
| miR-545-5p | – | AGTR1, NPPA | – | – | – |
| miR-550a-5p | – | GPR182, NPR1 | – | NPR1 | – |
| miR-622 | – | AGT, NPPA, NPR1 | – | – | – |
| miR-638 | – | CYP11B2 | – | – | – |
| miR-671-5p | – | ACE, CYP11B2, CRHR2 | DN1(82) | – | – |
| miR-675 | – | – | – | – | – |
| miR-7-5p | – | AGT, AGTR1 | – | AGTR1, EDN1, NPR3 | ACE, CALCRL, ATP6AP2, EDN1, EDNRA, CRHR1, CRHR2 |
| miR-92a-3p | – | NPR1 | – | AGTR2, ADM, EDNRB, NR3C2 | AGTR1, CALCRL, EDNRA, EDNRB, NPR2, NR3C2 |
| miR-92b-3p | – | NPR1 | – | AGTR2, ADM, EDNRB, NR3C2 | EDNRA |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wong, L.L.; Wang, J.; Liew, O.W.; Richards, A.M.; Chen, Y.-T. MicroRNA and Heart Failure. Int. J. Mol. Sci. 2016, 17, 502. https://doi.org/10.3390/ijms17040502
Wong LL, Wang J, Liew OW, Richards AM, Chen Y-T. MicroRNA and Heart Failure. International Journal of Molecular Sciences. 2016; 17(4):502. https://doi.org/10.3390/ijms17040502
Chicago/Turabian StyleWong, Lee Lee, Juan Wang, Oi Wah Liew, Arthur Mark Richards, and Yei-Tsung Chen. 2016. "MicroRNA and Heart Failure" International Journal of Molecular Sciences 17, no. 4: 502. https://doi.org/10.3390/ijms17040502
APA StyleWong, L. L., Wang, J., Liew, O. W., Richards, A. M., & Chen, Y. -T. (2016). MicroRNA and Heart Failure. International Journal of Molecular Sciences, 17(4), 502. https://doi.org/10.3390/ijms17040502