
High Morphologic Plasticity of Microglia/Macrophages Following Experimental Intracerebral Hemorrhage in Rats

Abstract
:1. Introduction
2. Results
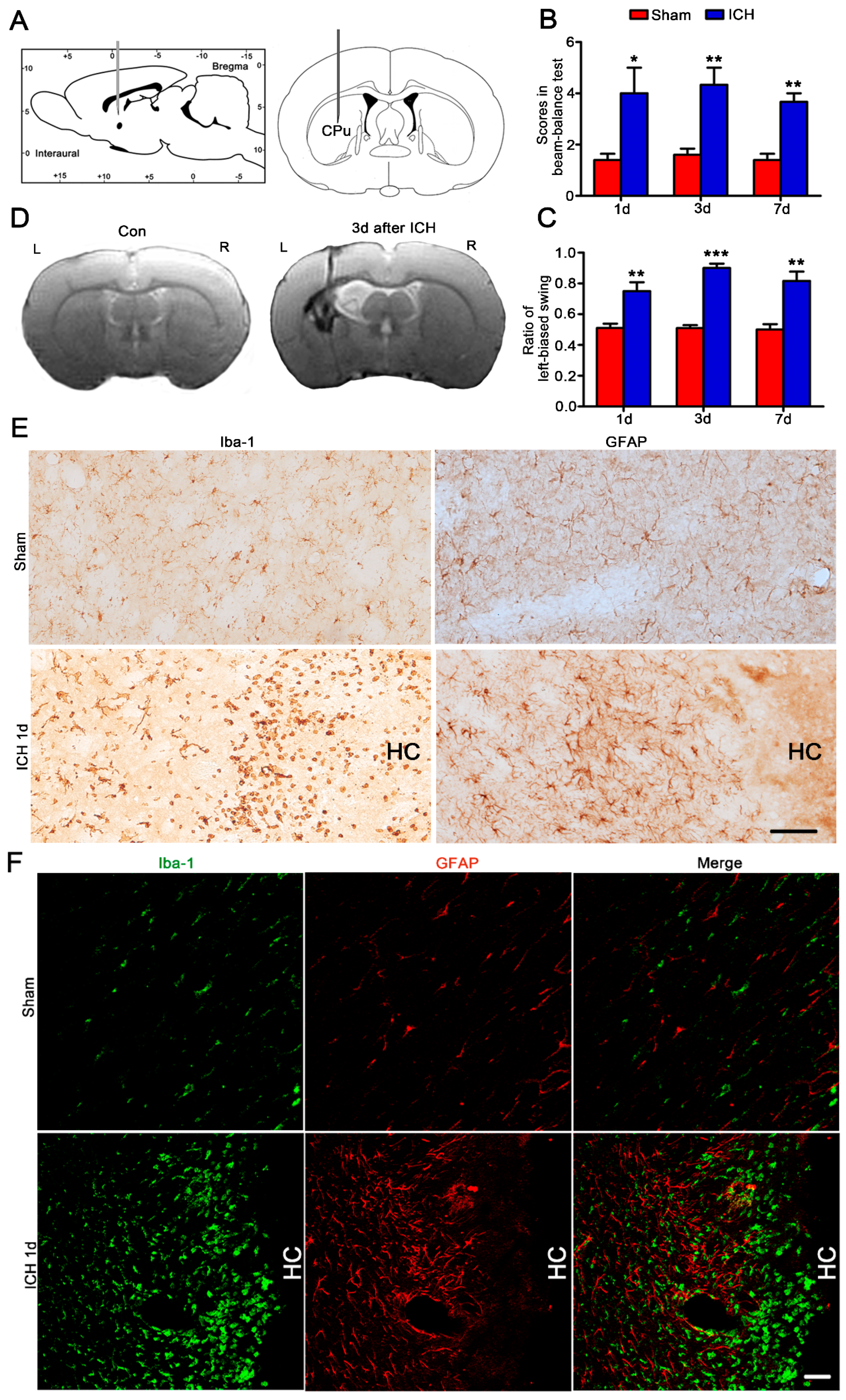
2.1. Increased and Accumulated Microglia/Macrophages and Astrocytes in the Hemorrhagic CPU of Rats
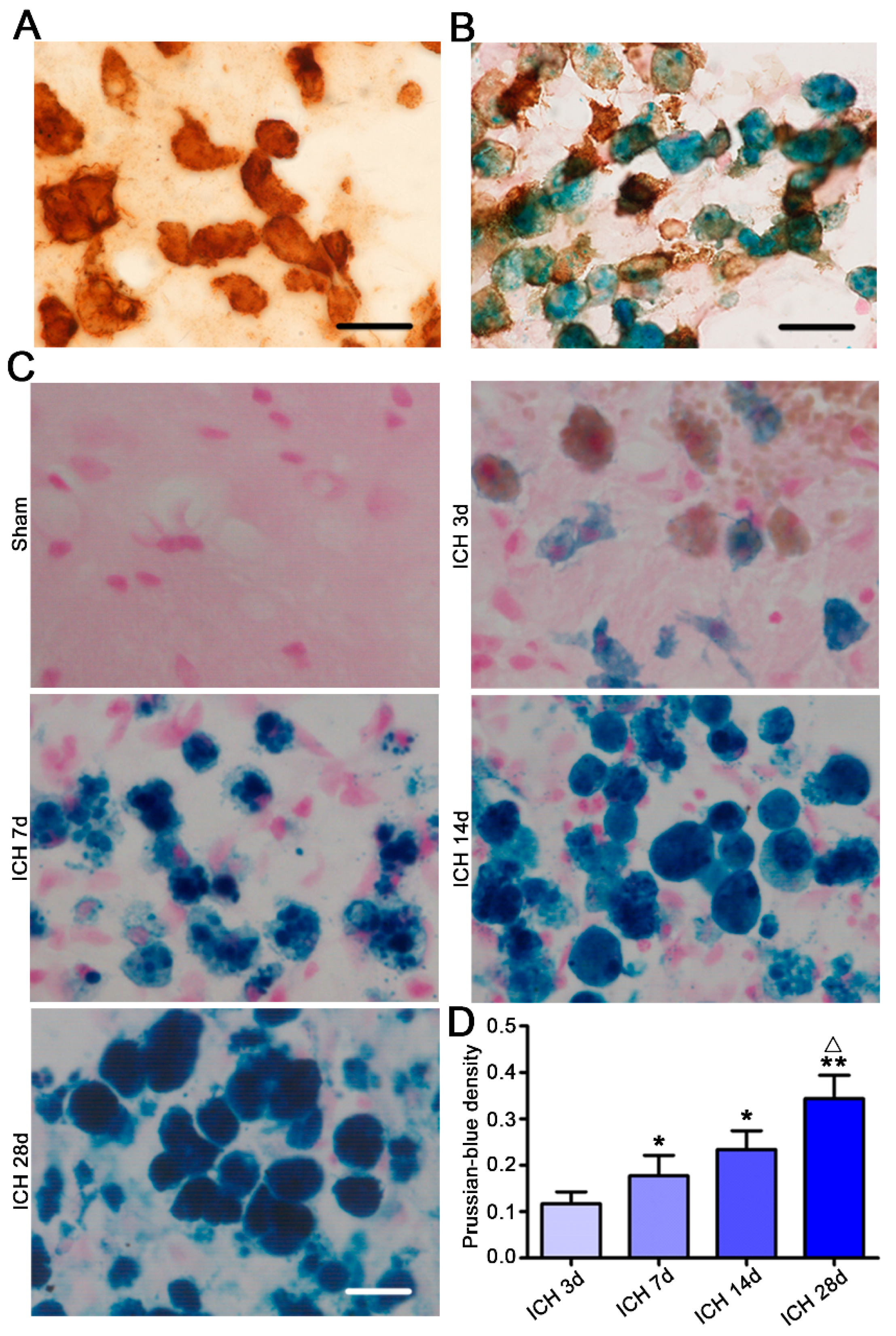
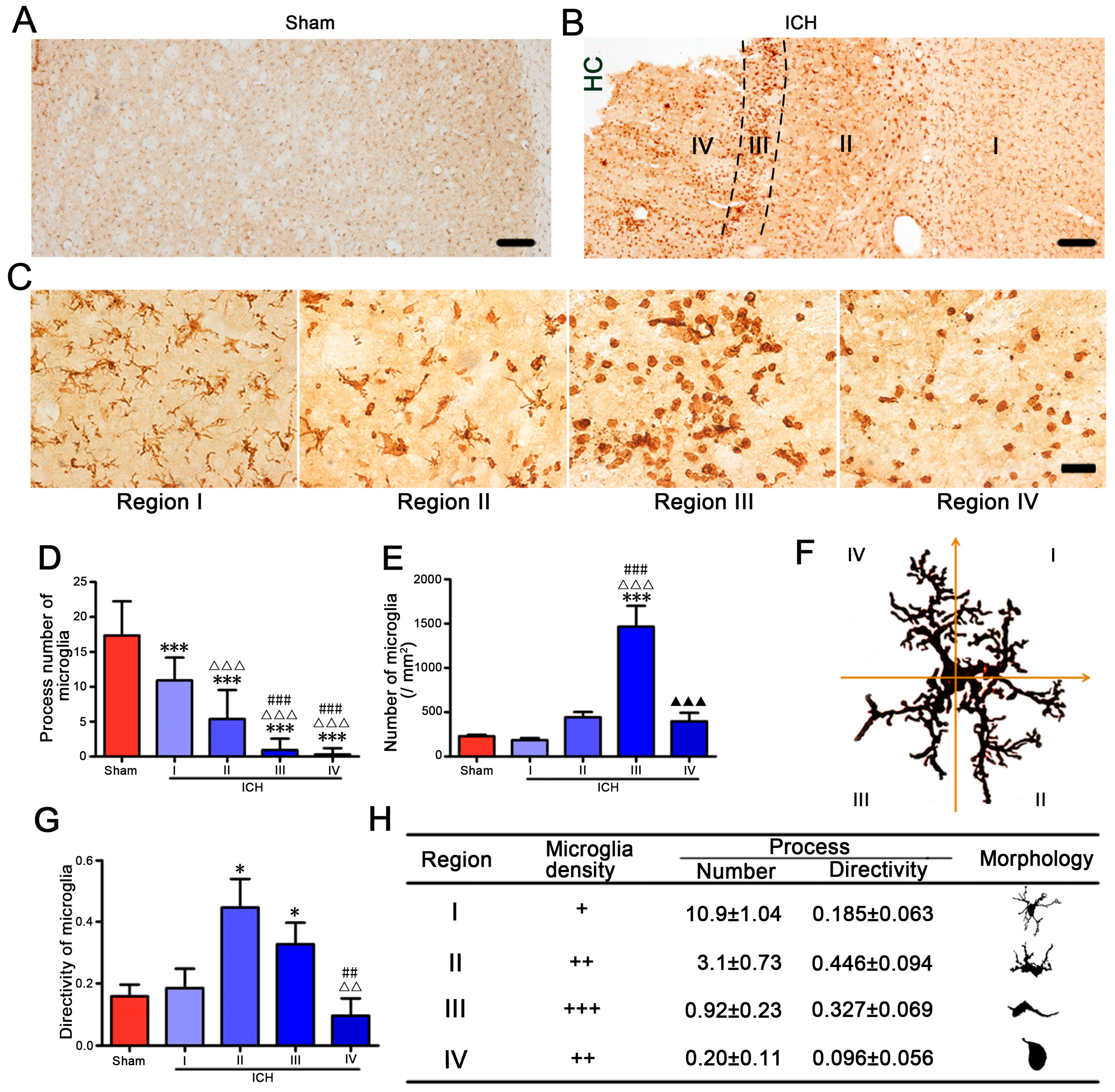
2.2. Microglia/Macrophages Nearest to the Hemorrhagic Center Were Ameboid and Prussian Blue Positive
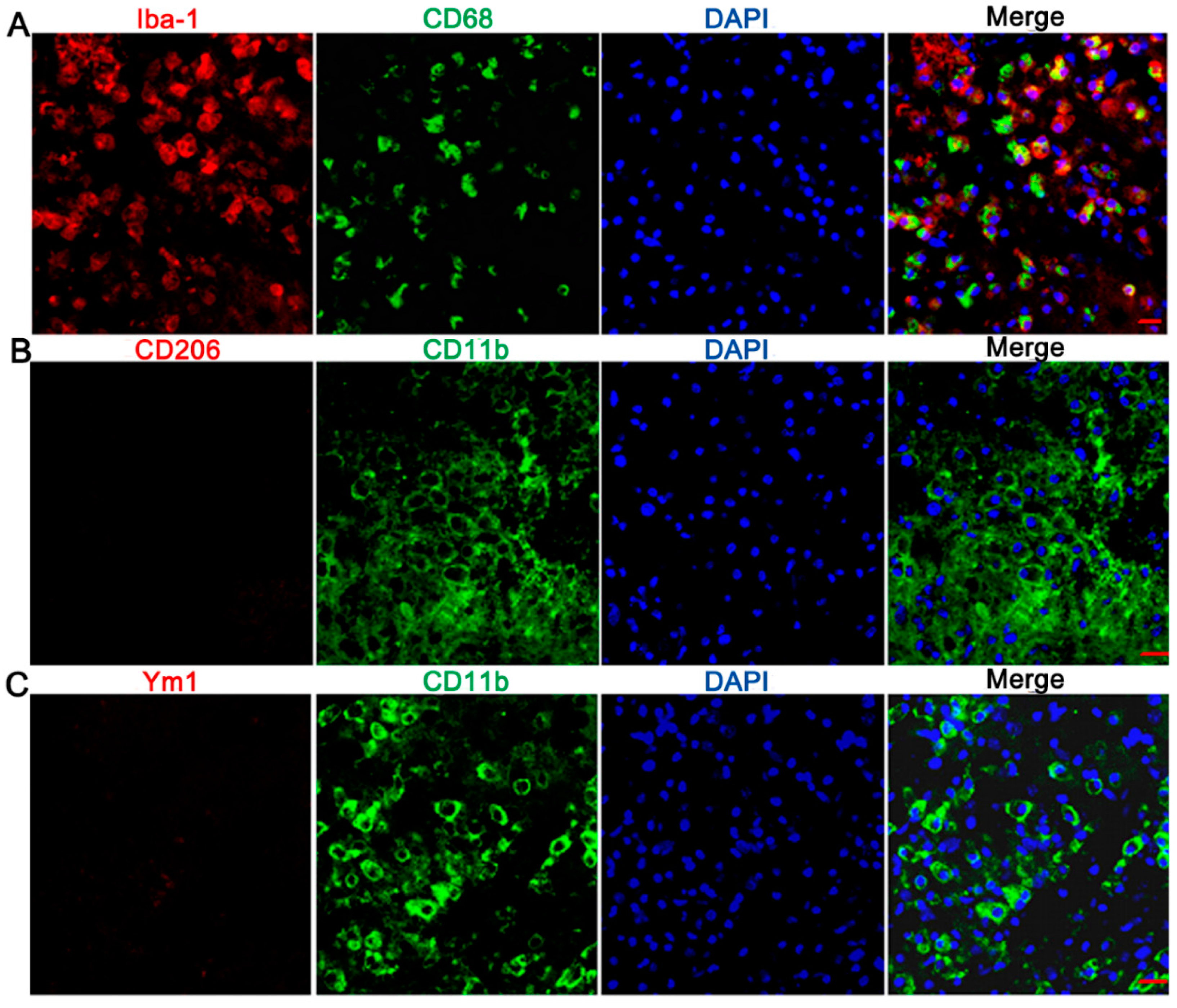
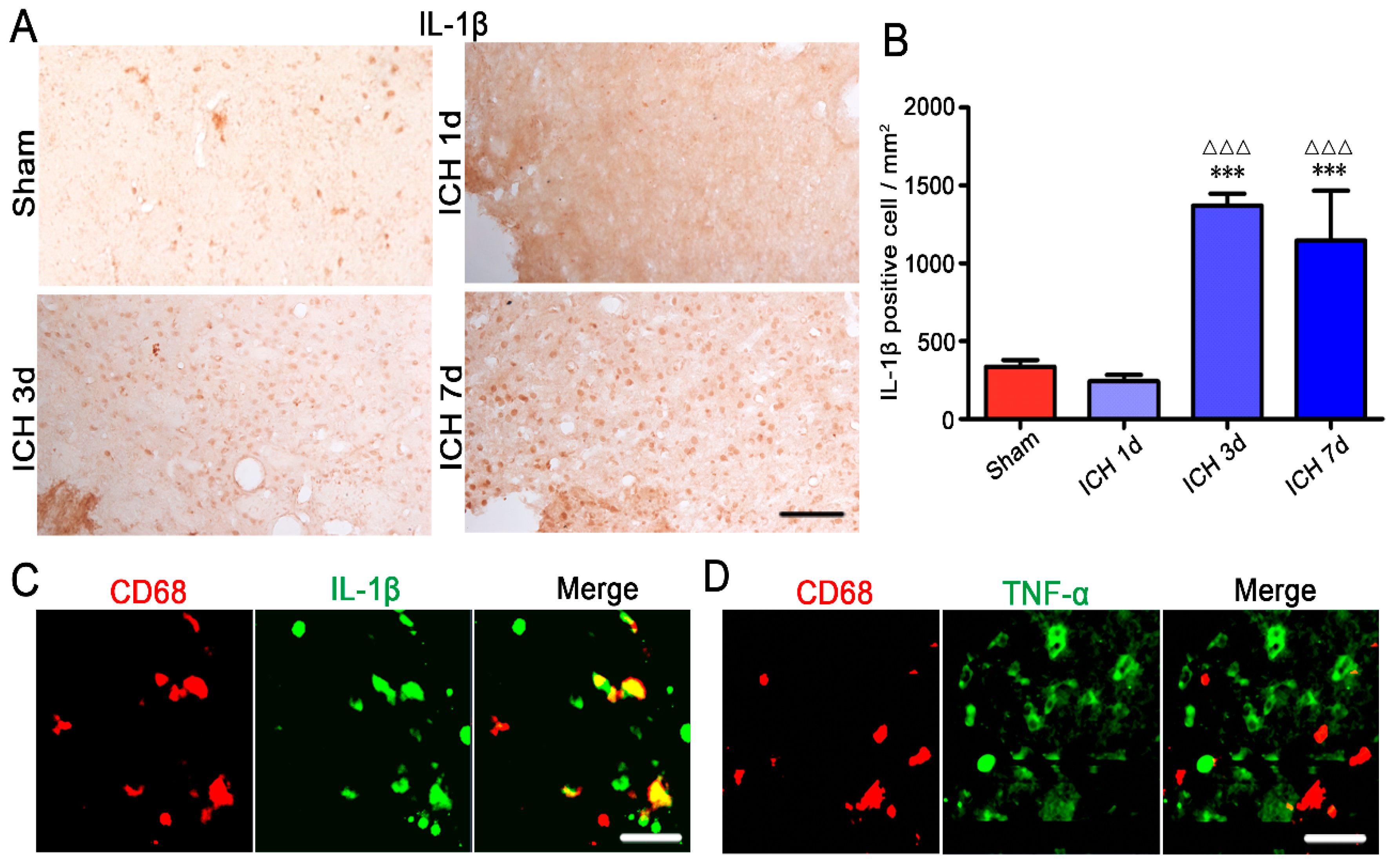
2.3. Ameboid Microglia/Macrophages Presented CD68 and IL-1β Positive
2.4. High Morphologic Plasticity of Microglia/Macrophages in Hemorrhagic CPu
3. Discussion
4. Materials and Methods
4.1. Antibodies and Chemicals
4.2. Animals and Stereotactic Injections of CPus
4.3. Behavior Tests
4.4. Magnetic Resonance Imaging (MRI)
4.5. Morphological Techniques on Brain Slices
4.5.1. Immunohistochemical Staining
4.5.2. Double-Label Immunofluorescence Staining
4.5.3. Prussian Blue Staining
4.6. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Feigin, V.L.; Lawes, C.M.; Bennett, D.A.; Barker-Collo, S.L.; Parag, V. Worldwide stroke incidence and early case fatality reported in 56 population-based studies: A systematic review. Lancet Neurol. 2009, 8, 355–369. [Google Scholar] [CrossRef]
- Krishnamurthi, R.V.; Moran, A.E.; Forouzanfar, M.H.; Bennett, D.A.; Mensah, G.A.; Lawes, C.M.; Barker-Collo, S.; Connor, M.; Roth, G.A.; Sacco, R.; et al. Global Burden of Diseases, Injuries, and Risk Factors 2010 Study Stroke Expert Group, The global burden of hemorrhagic stroke: A summary of findings from the GBD 2010 study. Glob. Heart 2014, 9, 101–106. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, M.J.; Xavier, D.; Liu, L.; Zhang, H.; Chin, S.L.; Rao-Melacini, P.; Rangarajan, S.; Islam, S.; Pais, P.; McQueen, M.J.; et al. Risk factors for ischaemic and intracerebral haemorrhagic stroke in 22 countries (the Interstroke study): A case-control study. Lancet 2010, 376, 112–123. [Google Scholar] [CrossRef]
- Mendelow, A.D.; Gregson, B.A.; Mitchell, P.M.; Murray, G.D.; Rowan, E.N.; Gholkar, A.R.; STICH II Investigators. Surgical trial in lobar intracerebral haemorrhage (Stich II) protocol. Trials 2011, 12, 124. [Google Scholar] [CrossRef] [PubMed]
- Kuramatsu, J.B.; Huttner, H.B.; Schwab, S. Advances in the management of intracerebral hemorrhage. J. Neural Transm. 2013, 120, S35–S41. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Garton, T.; Keep, R.F.; Hua, Y.; Xi, G. Microglia/Macrophage Polarization After Experimental Intracerebral Hemorrhage. Transl. Stroke Res. 2015, 6, 407–409. [Google Scholar] [CrossRef] [PubMed]
- Schilling, M.; Besselmann, M.; Müller, M.; Strecker, J.K.; Ringelstein, E.B.; Kiefer, R. Predominant phagocytic activity of resident microglia over hematogenous macrophages following transient focal cerebral ischemia: An investigation using green fluorescent protein transgenic bone marrow chimeric mice. Exp. Neurol. 2005, 196, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Melinda, E.L.; Michelle, L.B. Microglial activation and chronic neurodegeneration. Neurotheapeutics 2010, 7, 354–365. [Google Scholar] [Green Version]
- Aguzzi, A.; Barres, B.A.; Bennett, M.L. Microglia: Scapegoat, saboteur, or something else? Science 2013, 339, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Colton, C.A. Heterogeneity of microglial activation in the innate immune response in the brain. J. Neuroimmune Pharmacol. 2009, 4, 399–418. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.A. Age-Related Neuroinflammatory Changes Negatively Impact on Neuronal Function. Front. Aging Neurosci. 2009, 1, 6. [Google Scholar] [CrossRef] [PubMed]
- Loane, D.J.; Kumar, A. Microglia in the TBI brain: The good, the bad, and the dysregulated. Exp. Neurol. 2016, 275, 316–327. [Google Scholar] [CrossRef] [PubMed]
- Febinger, H.Y.; Thomasy, H.E.; Pavlova, M.N.; Ringgold, K.M.; Barf, P.R.; George, A.M.; Grillo, J.N.; Bachstetter, A.D.; Garcia, J.A.; Cardona, A.E. Time-Dependent effects of CX3CR1 in a mouse model of mild traumatic brain injury. J. Neuroinflamm. 2015, 12, 154. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, G.A.; Mun-Bryce, S.; Wesley, M.; Kornfeld, M. Collagenase-induced intracerebral hemorrhage in rats. Stroke 1990, 21, 801–807. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S. Alternative activation of macrophages. Nat. Rev. Immunol. 2003, 3, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Bedi, S.S.; Smith, P.; Hetz, R.A.; Xue, H.S.; Cox, C.S. Immunomagnetic enrichment and flow cytometric characterization of mouse microglia. J. Neurosci. Meth. 2013, 219, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Miron, V.E.; Boyd, A.; Zhao, J.W.; Yuen, T.J.; Ruckh, J.M.; Shadrach, J.L.; Wijngaarden, P.; Wagers, A.J.; Williams, A.; Franklin, R.J.M.; et al. M2 microglia and macrophages drive oligodendrocyte differentiation during CNS remyelination. Nat. Neurosci. 2013, 16, 1211–1218. [Google Scholar] [CrossRef] [PubMed]
- Bowyer, J.F.; Sarkar, S.; Tranter, K.M.; Hanig, J.P.; Miller, D.B.; O’Callaghan, J.P. Vascular-directed responses of microglia produced by methamphetamine exposure: Indirect evidence that microglia are involved in vascular repair? J. Neuroinflamm. 2016, 13. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhang, J.; Hu, X.; Zhang, L.; Mao, L.; Jiang, X.; Liou, A.K.; Leak, R.K.; Gao, Y.; Chen, J. Microglia/macrophage polarization dynamics in white matter after traumatic brain injury. J. Cereb. Blood. Flow Metab. 2013, 33, 1864–1874. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Yang, S.S.; Zeng, P; Zhang, T.; Liu, Y.; Wang, J.; Tian, Q. Effect of emodin on experimental intracerebral hemorrhage in rats. Unpublished. 2016. [Google Scholar]
- MacLellan, C.L.; Silasi, G.; Poon, C.C.; Edmundson, C.L.; Buist, R.; Peeling, J.; Colbourne, F. Intracerebral hemorrhage models in rat: Comparing collagenase to blood infusion. J. Cereb. Blood Flow. Metab. 2008, 28, 516–525. [Google Scholar] [CrossRef] [PubMed]
- MacLellan, C.L.; Silasi, G.; Auriat, A.M.; Colbourne, F. Rodent models of intracerebral hemorrhage. Stroke 2010, 41, S95–S98. [Google Scholar] [CrossRef] [PubMed]
- Bullock, R.; Mendelow, A.D.; Teasdale, G.M.; Graham, D.I.; McCulloch, J. Intracranial haemorrhage induced at arterial pressure in the rat. Part 1: Description of technique, ICP changes and neuropathological findings. Neurol. Res. 1984, 6, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, Z.; Lu, H.; Yang, Q.; Wu, H.; Wang, J. Microglial polarization and inflammatory mediators after intracerebral hemorrhage. Mol. Neurobiol. 2016, in press. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Doré, S. Heme oxygenase-1 exacerbates early brain injury after intracerebral haemorrhage. Brain 2007, 130, 1643–1652. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; del Bigio, M.R. Intracerebral injection of autologous whole blood in rats: Time course of inflammation and cell death. Neurosci. Lett. 2000, 283, 230–232. [Google Scholar] [CrossRef]
- Wang, J. Preclinical and clinical research on inflammation after intracerebral hemorrhage. Prog. Neurobiol. 2010, 92, 463–477. [Google Scholar] [CrossRef] [PubMed]
- Sansing, L.H.; Harris, T.H.; Welsh, F.A.; Kasner, S.E.; Hunter, C.A.; Kariko, K. Toll-like receptor 4 contributes to poor outcome after intracerebral hemorrhage. Ann. Neurol. 2011, 70, 646–656. [Google Scholar] [CrossRef] [PubMed]
- Kreutzberg, G.W. Microglia: A sensor for pathological events in the CNS. Trends Neurosci. 1996, 19, 312–318. [Google Scholar] [CrossRef]
- Ginhoux, F.; Greter, M.; Leboeuf, M.; Nandi, S.; See, P.; Gokhan, S.; Mehler, M.F.; Conway, S.J.; Ng, L.G.; Stanley, E.R.; et al. Fate mapping analysis reveals that adult microglia derive from primitive macrophages. Science 2010, 330, 841–845. [Google Scholar] [CrossRef] [PubMed]
- Aronowski, J.; Zhao, X. Molecular pathophysiology of cerebral hemorrhage secondary brain injury. Stroke 2011, 42, 1781–1786. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 5th ed.; Academic Press: Sydney, Australia, 2004; p. 68, Figure 28. [Google Scholar]
- Zausinger, S.; Hungerhuber, E.; Baethmann, A.; Reulen, H.; Schmid-Elsaesser, R. Neurological impairment in rats after transient middle cerebral artery occlusion: A comparative study under various treatment paradigms. Brain Res. 2000, 863, 94–105. [Google Scholar] [CrossRef]
- Borlongan, C.V.; Sanberg, P.R. Elevated body swing test: A new behavioral parameter for rats with 6-hydroxydopamine-induced hemiparkinsonism. J. Neurosci. 1995, 15, 5372–5378. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Type | Dilution | Source |
|---|---|---|---|
| Iba-1 | Poly- | 1:200 for IHC 1:200 for IF | Wako (Osaka, Japan) |
| CD68 | Mono- | 1:100 for IF | Abcam (Cambridge, UK) |
| GFAP | Mono- | 1:200 for IHC 1:200 for IF | Abcam |
| IL-1β | Poly- | 1:100 for IHC 1:50 for IF | Santa Cruz Biotechnology (Delaware Ave Santa Cruz, CA, USA) |
| TNF-α | Poly- | 1:50 for IF | Millipore (Billerica, MA, USA) |
| CD11b | Mono- | 1:50 for IF | Bio-Rad (Hercules, CA, USA) |
| Ym1 | Poly- | 1:100 for IF | Stem Cell Technologies (Vancouver, BC, Canada) |
| CD206 | Poly- | 1:100 for IF | R&D Systems (Minneapolis, MN, USA) |
| goat anti-mouse IgG (H + L) | Rhodamine Red-X conjugated | 1:1000 for IF | Invitrogen (Carlsbad, CA, USA) |
| goat anti-mouse IgG (H + L) | Oregon Green 488 conjugated | 1:1000 for IF | Invitrogen |
| goat anti-rabbit IgG (H + L) | Rhodamine Red-X conjugated | 1:1000 for IF | Invitrogen |
| goat anti-rabbit IgG (H + L) | Oregon Green 488 conjugated | 1:1000 for IF | Invitrogen |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.-S.; Lin, L.; Liu, Y.; Wang, J.; Chu, J.; Zhang, T.; Ning, L.-N.; Shi, Y.; Fang, Y.-Y.; Zeng, P.; et al. High Morphologic Plasticity of Microglia/Macrophages Following Experimental Intracerebral Hemorrhage in Rats. Int. J. Mol. Sci. 2016, 17, 1181. https://doi.org/10.3390/ijms17071181
Yang S-S, Lin L, Liu Y, Wang J, Chu J, Zhang T, Ning L-N, Shi Y, Fang Y-Y, Zeng P, et al. High Morphologic Plasticity of Microglia/Macrophages Following Experimental Intracerebral Hemorrhage in Rats. International Journal of Molecular Sciences. 2016; 17(7):1181. https://doi.org/10.3390/ijms17071181
Chicago/Turabian StyleYang, Shu-Sheng, Li Lin, Yue Liu, Jie Wang, Jiang Chu, Teng Zhang, Lin-Na Ning, Yan Shi, Ying-Yan Fang, Peng Zeng, and et al. 2016. "High Morphologic Plasticity of Microglia/Macrophages Following Experimental Intracerebral Hemorrhage in Rats" International Journal of Molecular Sciences 17, no. 7: 1181. https://doi.org/10.3390/ijms17071181
APA StyleYang, S. -S., Lin, L., Liu, Y., Wang, J., Chu, J., Zhang, T., Ning, L. -N., Shi, Y., Fang, Y. -Y., Zeng, P., Wang, J. -Z., Qiu, M. -Y., & Tian, Q. (2016). High Morphologic Plasticity of Microglia/Macrophages Following Experimental Intracerebral Hemorrhage in Rats. International Journal of Molecular Sciences, 17(7), 1181. https://doi.org/10.3390/ijms17071181