
β-Ketoacyl-acyl Carrier Protein Synthase I (KASI) Plays Crucial Roles in the Plant Growth and Fatty Acids Synthesis in Tobacco


Abstract
:
1. Introduction
2. Results
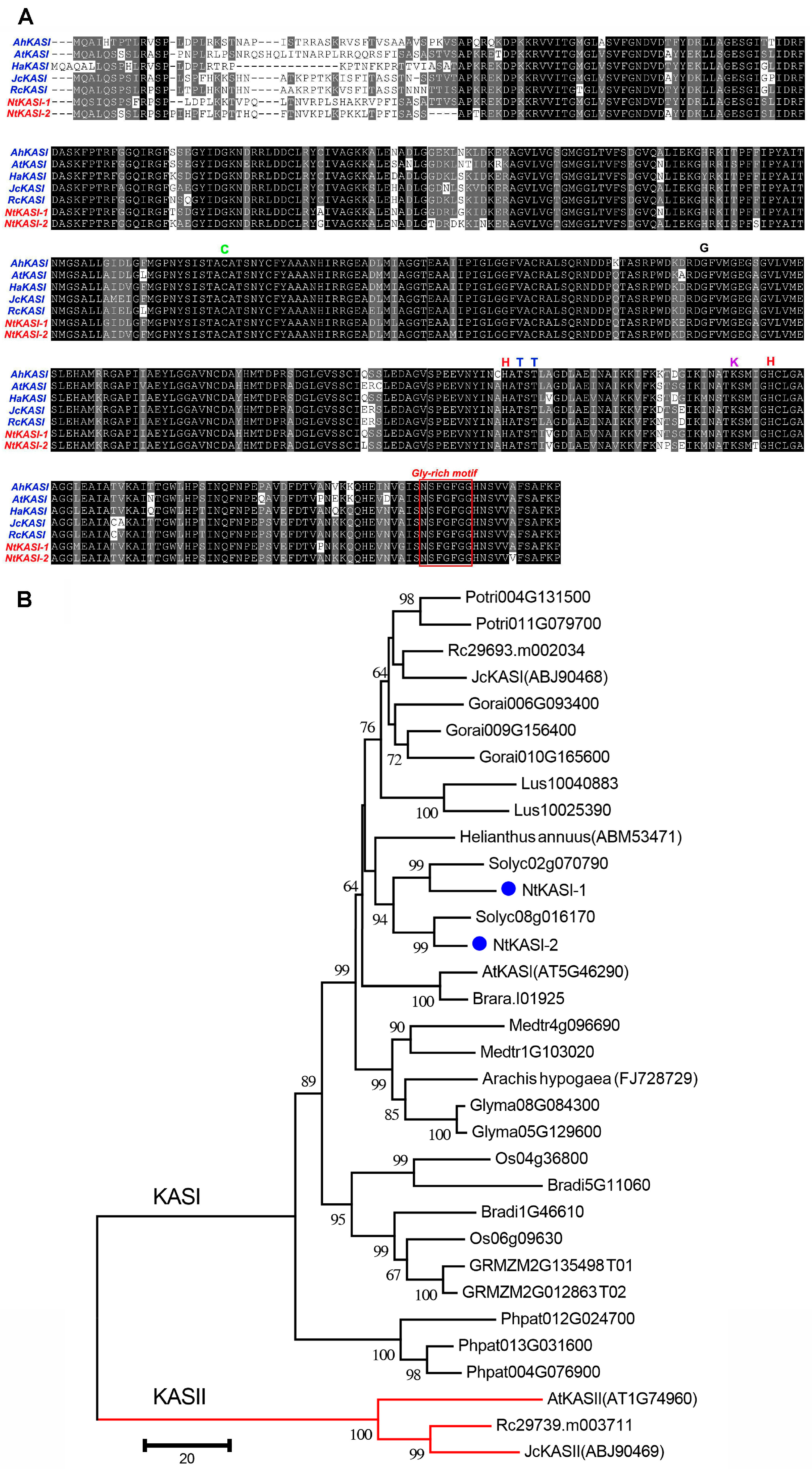
2.1. Identification of 3-Ketoacyl-ACP Synthase I Gene in Tobacco
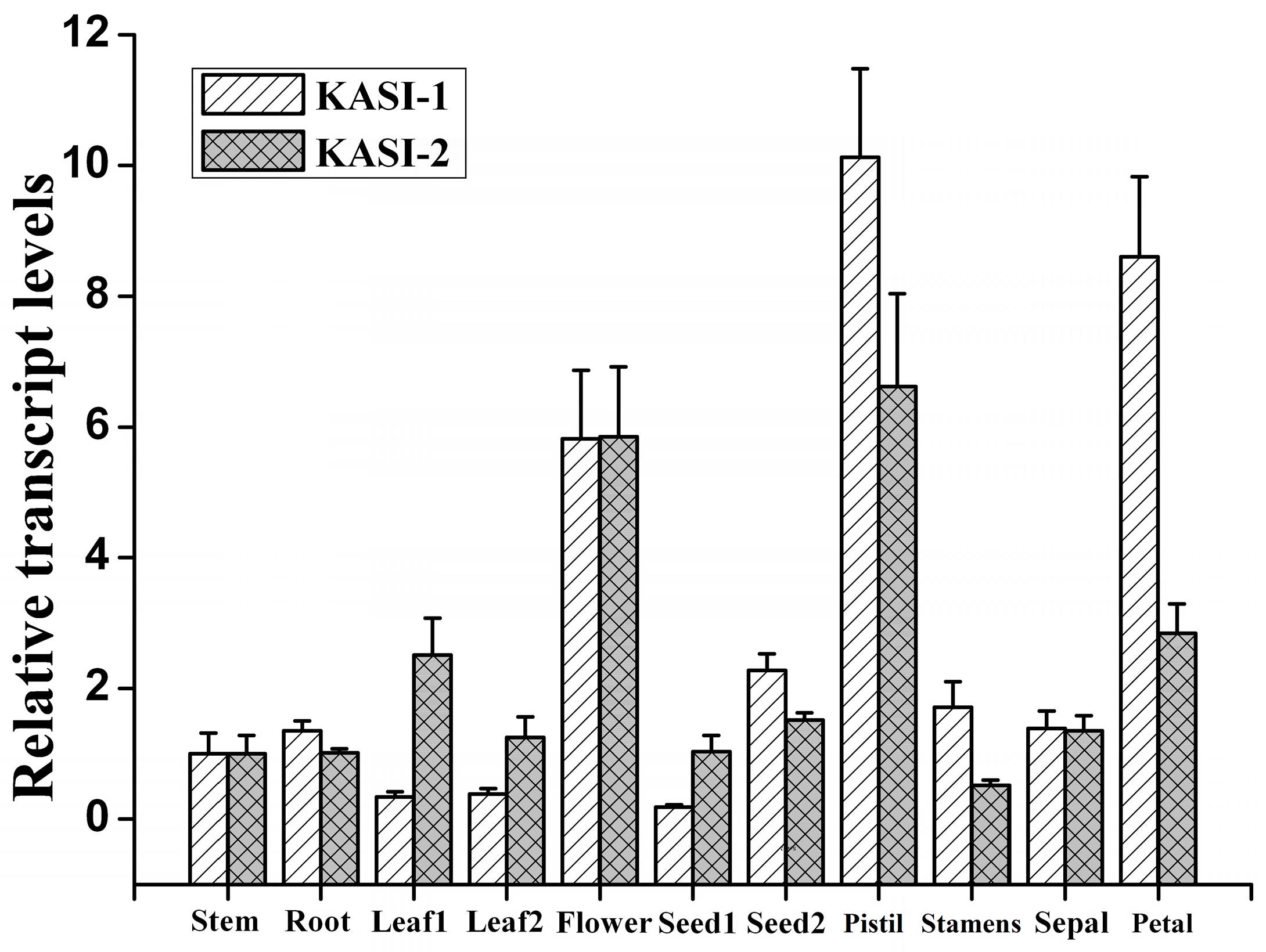
2.2. Expression Patterns of NtKASI-1 and NtKASI-2
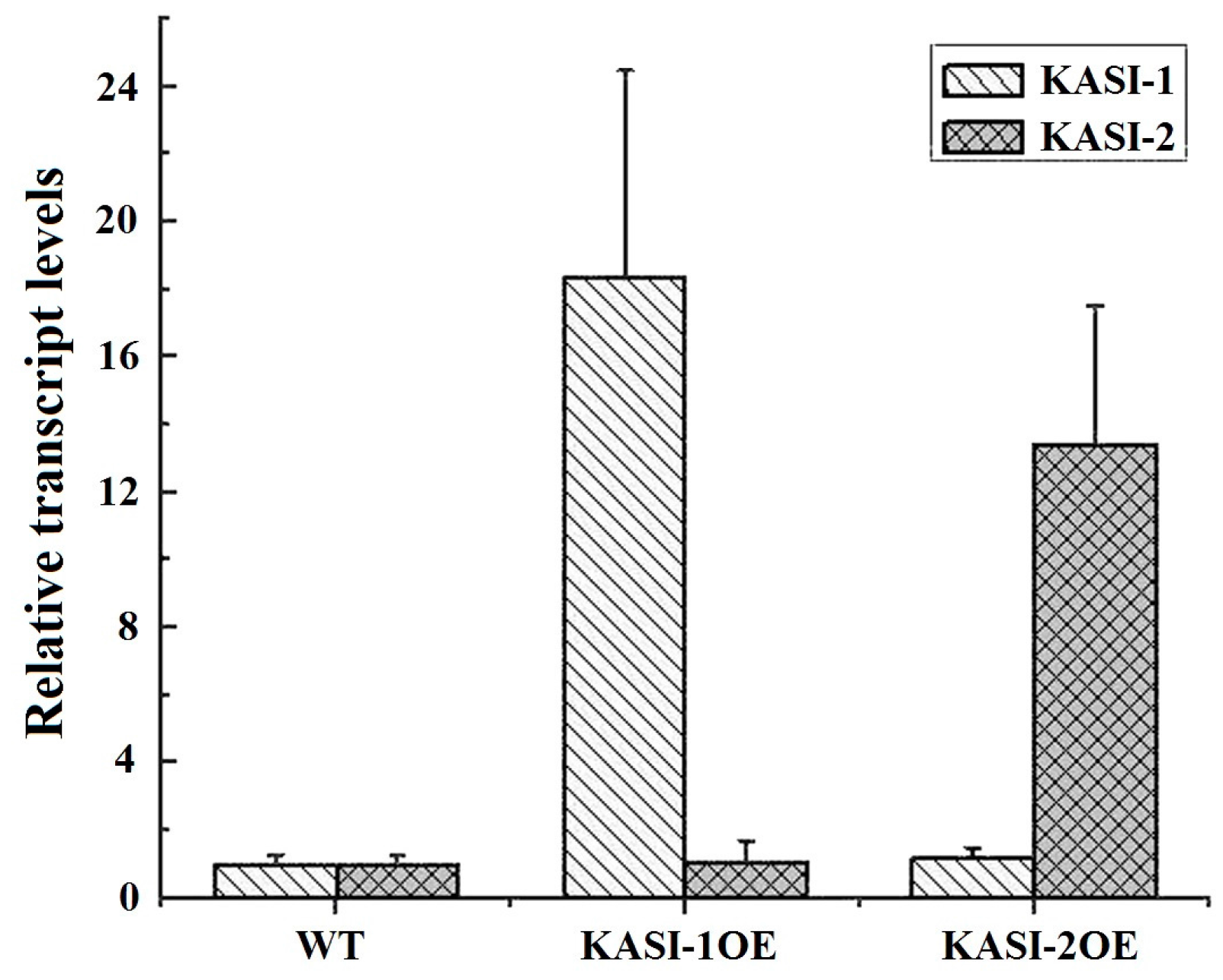
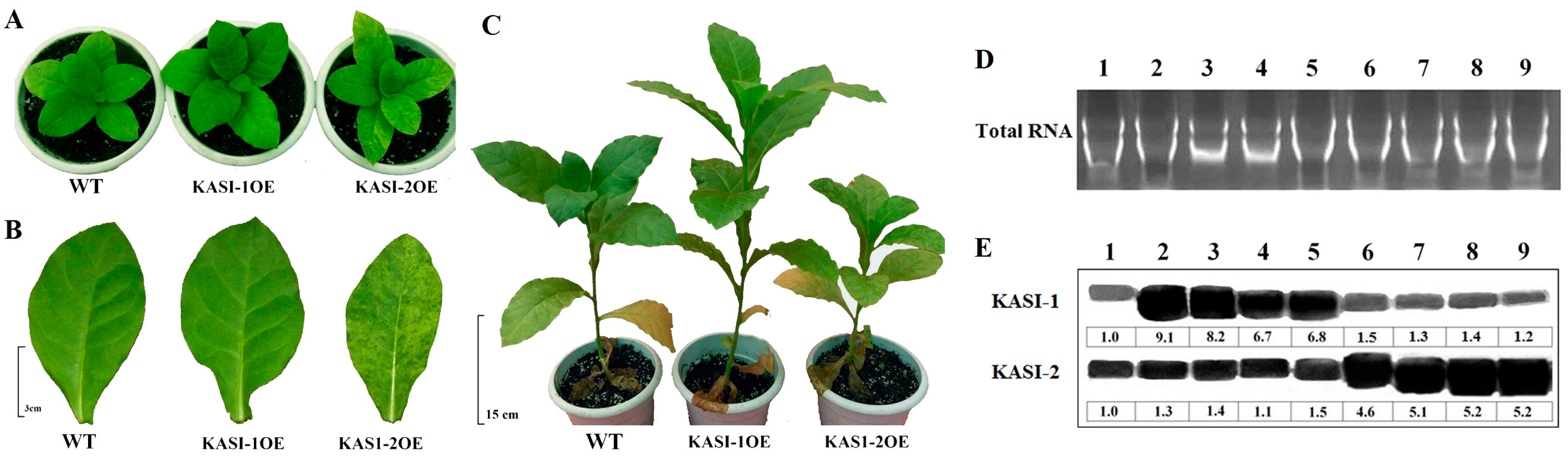
2.3. Phenotypes of Over-Expression of NtKASI Genes in Tobacco
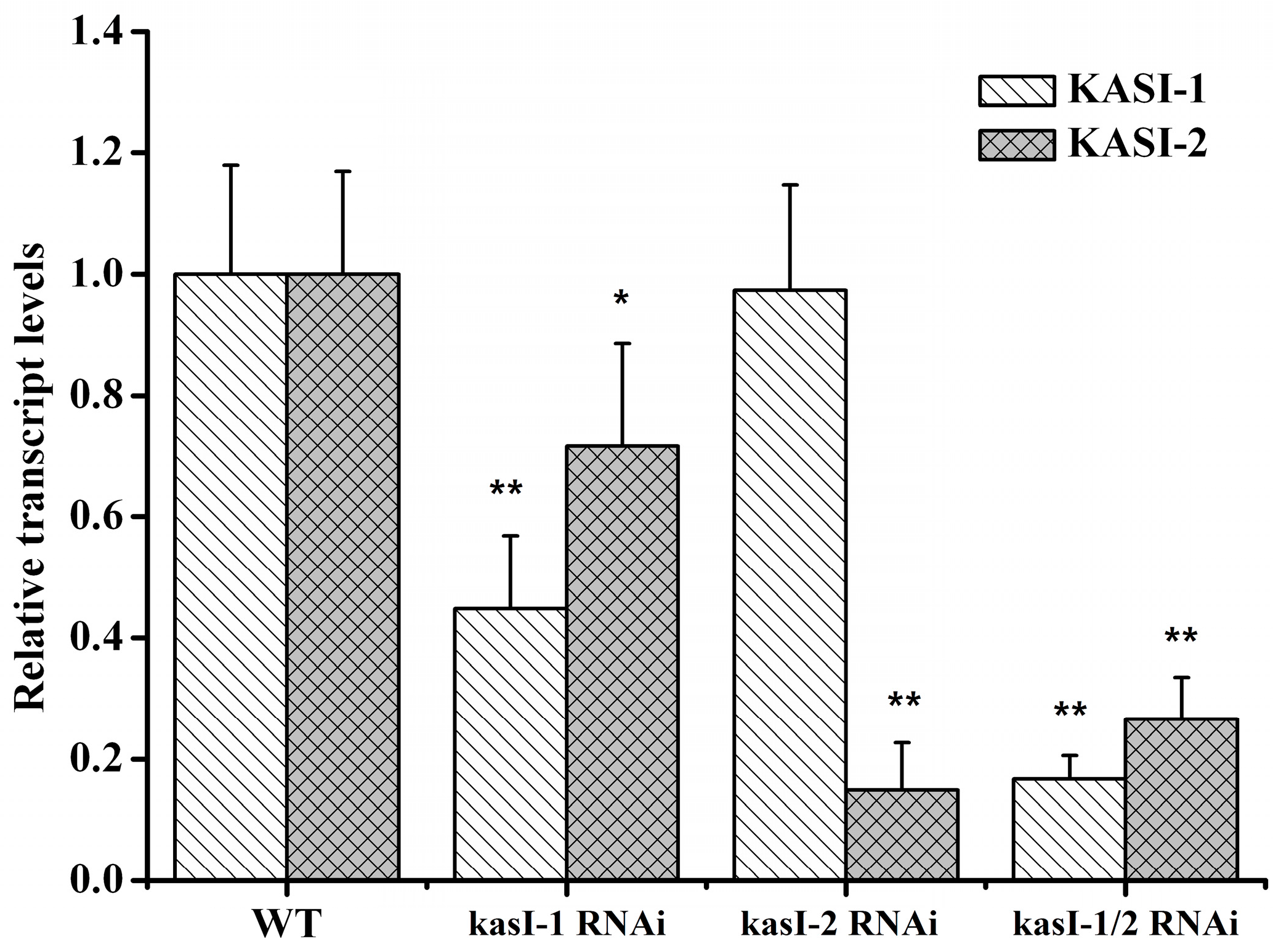
2.4. Silencing of NtKASI Genes in Tobacco
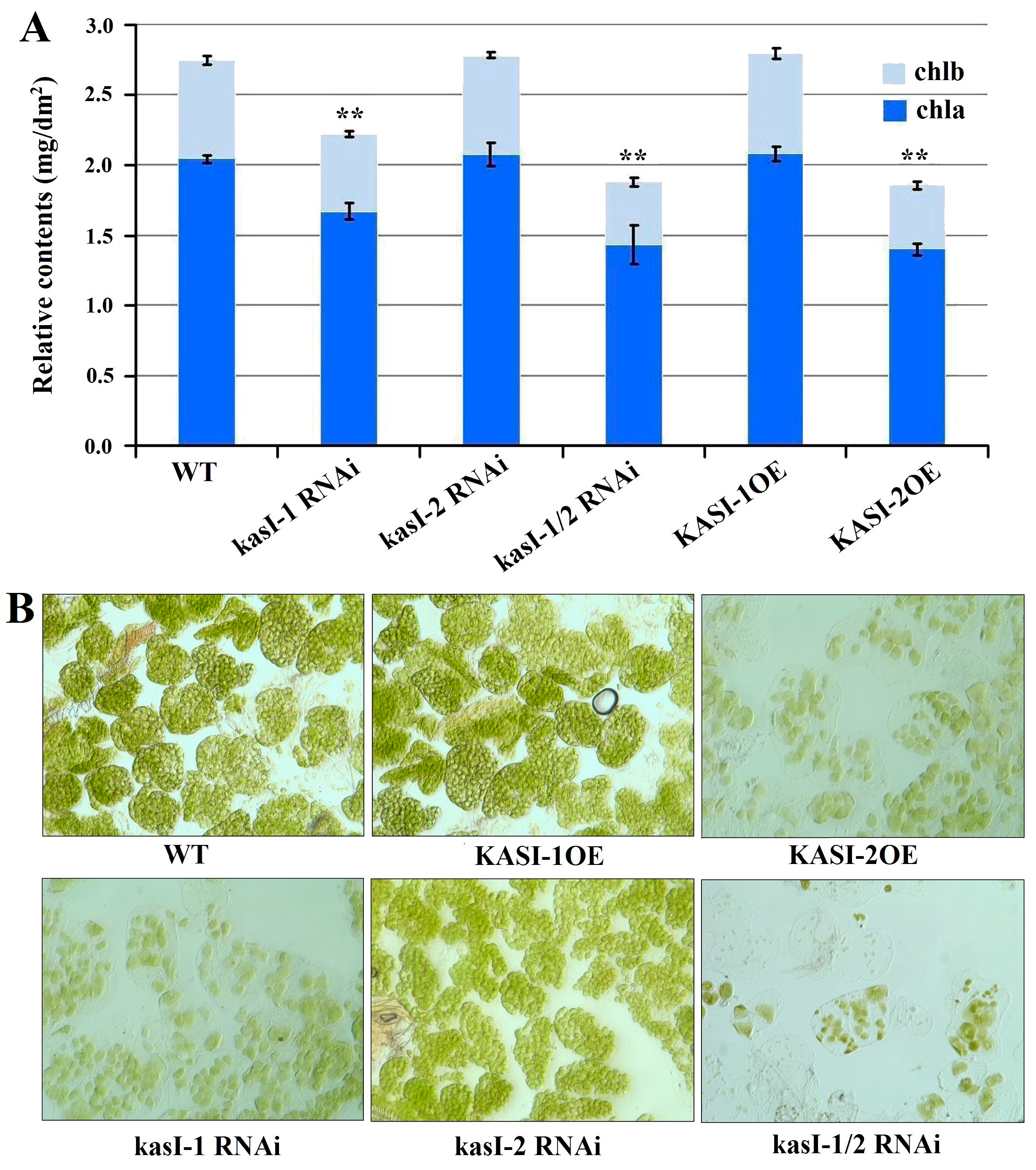
2.5. NtKASI Genes Affected the Chloroplast Development
2.6. NtKASI Genes Change Fatty Acids Composition in Tobacco Leaf
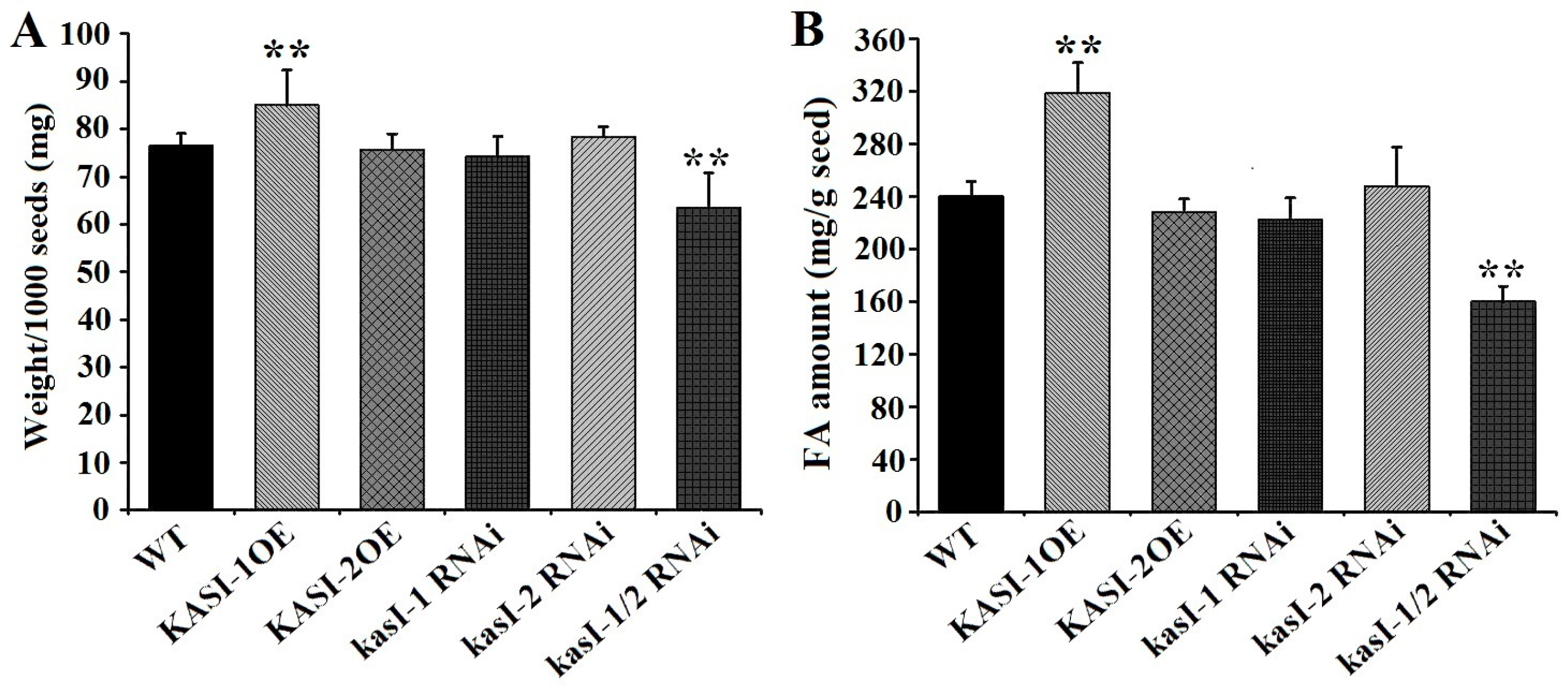
2.7. NtKASI Genes Affect the Seed Weight and Lipid Content
3. Discussion
4. Materials and Methods
4.1. Isolation and Sequence Analysis of Tobacco KASI Genes
4.2. Vector Construction and Transformation
4.3. Expression Analyses of KASI Genes in Wild-Type and Transgenic Tobacco
4.4. Morphological Observation of Chloroplast and Measurement of Chlorophylls
4.5. Seed Weight Determination and Lipid Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| FA | Fatty Acid |
| ACCase | Acetyl-CoA Carboxylase |
| CoA | Coenzyme A |
| ACP | Acyl Carrier Protein |
| KAS | 3-Ketoacyl-Acyl-Carrier Protein Synthase |
| TAG | Triacylglycerol |
| GC | Gas Chromatographic |
References
- Bonaventure, G.; Salas, J.J.; Pollard, M.R.; Ohlrogge, J.B. Disruption of the FATB gene in Arabidopsis demonstrates an essential role of saturated fatty acids in plant growth. Plant Cell 2003, 15, 1020–1033. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.Q.; Rowland, O.; Kunst, L. Disruptions of the Arabidopsis enoyl-CoA reductase gene reveal an essential role for very-long-chain fatty acid synthesis in cell expansion during plant morphogenesis. Plant Cell 2005, 17, 1467–1481. [Google Scholar] [CrossRef] [PubMed]
- Roudier, F.; Gissot, L.; Beaudoin, F.; Haslam, R.; Michaelson, L.; Marion, J.; Molino, D.; Lima, A.; Bach, L.; Morin, H.; et al. Very-long-chain fatty acids are involved in polar auxin transport and developmental patterning in Arabidopsis. Plant Cell 2010, 22, 364–375. [Google Scholar] [CrossRef] [PubMed]
- Durrett, T.P.; Benning, C.; Ohlrogge, J. Plant triacylglycerols as feedstocks for the production of biofuels. Plant J. 2008, 54, 593–607. [Google Scholar] [CrossRef] [PubMed]
- Dyer, J.M.; Stymne, S.; Green, A.G.; Carlsson, A.S. High value oil from plants. Plant J. 2008, 54, 640–655. [Google Scholar] [CrossRef] [PubMed]
- Gornicki, P.; Haselkorn, R. Wheat acetyl-CoA carboxylase. Plant Mol. Biol. 1993, 22, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Jackowski, S.; Rock, C.O. Acetoacetyl-acyl carrier protein synthase, a potential regulator of fatty acid biosynthsis in bacteria. J. Biol. Chem. 1987, 262, 7927–7931. [Google Scholar] [PubMed]
- Shimakata, T.; Stumpf, P.K. Purification and characterizations of beta-Ketacayl-[acyl-carrier-protein] reductase, beta-hydroxyacyl-[acyl-carrier-protein] dehydarse, and enoyl-[acyl-carrier-protein] reductase from Spinacia oleracea leaves. Arch. Biochem. Biophys. 1982, 218, 77–91. [Google Scholar] [CrossRef]
- Chi, X.Y.; Chen, M.N.; Yang, Q.L.; He, Y.N.; Pan, L.J.; Gao, Y.; Yu, S.L. Isolation and expression analysis of a β-ketoacyl-acyl carrier protein synthaseⅠ gene from Arachis hypogaea L. Legume Genom. Genet. 2010, 1, 11–17. [Google Scholar]
- Wu, G.Z.; Xue, H.W. Arabidopsis β-ketoacyl-[acyl carrier protein] synthase I is crucial for fatty acid synthesis and plays a role in chloroplast division and embryo development. Plant Cell 2010, 22, 3726–3744. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Li, J.; Zhang, L.; Wu, P.Z.; Chen, Y.P.; Li, M.R.; Jiang, H.W.; Wu, G.J. Cloning and characterization of a β-Ketoacyl-acyl carrier protein Synthase II from Jatropha curcas. J. Plant Physiol. 2012, 169, 816–824. [Google Scholar] [CrossRef] [PubMed]
- González-Mellado, D.; von Wettstein-Knowles, P.; Garcés, R.; Martínez-Force, E. The role of β-ketoacyl-acyl carrier protein synthase III in the condensation steps of fatty acid biosynthesis in sunflower. Planta 2010, 231, 1277–1289. [Google Scholar] [CrossRef] [PubMed]
- Stoll, C.; Lühs, W.; Zarhloul, M.K.; Friedt, W. Genetic modification of saturated fatty acids in oilseed rape (Brassica napus). Eur. J. Lipid Sci. Technol. 2005, 107, 244–248. [Google Scholar] [CrossRef]
- Schutt, B.S.; Abbadi, A.; Loddenkotter, B.; Brummel, M.; Spener, F. β-ketoacyl- acyl carrier protein synthase IV: A key enzyme for regulation of medium-chain fatty acid synthesis in Cuphea lanceolata seeds. Planta 2002, 215, 847–854. [Google Scholar] [CrossRef] [PubMed]
- Jaworski, J.G.; Clough, R.C.; Barnum, S.R. A cerulenin insensitive short chain 3–ketoacyl–acyl carrier protein synthase in Spinacia oleracea leaves. Plant Physiol. 1989, 90, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Clough, R.C.; Matthis, A.L.; Barnum, S.R.; Jaworski, J.G. Purification and characterization of 3-ketoacyl-acyl carrier protein synthase III from spinach: A condensing enzyme utilizing acetyl-coenzyme-A to initiate fatty acid synthesis. J. Biol. Chem. 1992, 267, 20992–20998. [Google Scholar] [PubMed]
- Murphy, D.J. Plant Lipids: Biology, Utilization and Manipulation; Blackwell Pubishing: Oxford, UK, 2005. [Google Scholar]
- Dehesh, K.; Edwards, P.; Fillatti, J.; Slabaugh, M.; Byrne, J. KAS IV: A 3-ketoacyl-ACP synthase from Cuphea sp. is a medium chain specific condensing enzyme. Plant J. 1998, 15, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Yasuno, R.; von Wettstein-Knowles, P.; Wada, H. Identification and molecular characterization of the β-ketoacyl-[acyl carrier protein] synthase component of the Arabidopsis mitochondrial fatty acid synthase. J. Biol. Chem. 2004, 279, 8242–8251. [Google Scholar] [CrossRef] [PubMed]
- Olsen, J.G.; Rasmussen, A.V.; von Wettstein–Knowles, P.; Henriksen, A. Structure of the mitochondrial beta-ketoacyl-[acyl carrier protein] synthase from Arabidopsis and its role in fatty acid synthesis. FEBS Lett. 2004, 577, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Tai, H.; Jaworski, J.G. 3-ketoacyl-acyl carrier protein synthase III from spinach (Spinacia olerecea) is not similar to other condensing enzyme of fatty acid synthase. Plant Physiol. 1993, 103, 1361–1367. [Google Scholar] [CrossRef] [PubMed]
- Tai, H.; Post-Beiyyenmiller, D.; Jaworski, J.G. Cloning of a cDNA encoding 3-ketoacyl-acyl carrier protein synthase III from Arabidopsis. Plant Physiol. 1994, 106, 801–802. [Google Scholar] [CrossRef] [PubMed]
- Slabaugh, M.B.; Tai, H.; Jaworski, J.G.; Knapp, S.J. cDNA clones encoding β-ketoacyl-acyl carrier protein synthase III from Cuphea wrightii. Plant Physiol. 1995, 108, 443–444. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Post-Beittenmiller, D. Molecular cloning of a cDNA encoding 3-ketoacyl-acyl carrier protein synthase III from leek. Gene 1996, 182, 45–52. [Google Scholar] [CrossRef]
- Jones, A.L.; Gane, A.M.; Herbert, D.; Willey, D.L.; Rutter, A.J.; Kille, P.; Dancer, J.E.; Harwood, J.L. β-Ketoacyl-acyl carrier protein synthase III from pea (Pisum sativum L.): Properties, inhibition by a novel thiolactomycin analogue and isolation of a cDNA clone encoding the enzyme. Planta 2003, 216, 752–761. [Google Scholar] [PubMed]
- Ye, J.; Qu, J.; Bui, H.T.; Chua, N.H. Rapid analysis of Jatropha curcas gene functions by virus-induced gene silencing. Plant Biotechnol. J. 2009, 7, 964–976. [Google Scholar] [CrossRef] [PubMed]
- Siggaard-Andersen, M.; Kauppinen, S.; von Wettstein-Knowles, P. Primary structure of a cerulenin-binding beta-ketoacyl-[acyl carrier protein] synthase from barley chloroplasts. Proc. Natl. Acad. Sci. USA 1991, 88, 4114–4118. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.; Lin, L.; Zhang, B.; Xiang, X.; Wu, J.; Pan, Z.; Zhu, S. OsKASI: A β-ketoacyl-[acyl carrier protein] synthase I: is involved in root development in rice (Oryza sativa L.). Planta 2015, 242, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Vanhercke, T.; EI Tahchy, A.; Liu, Q.; Zhou, X.R.; Shrestha, P.; Divi, U.K.; Ral, J.P.; Mansour, M.P.; Nichols, P.D.; James, C.N.; et al. Metabolic engineering of biomass for high energy density: Oilseed-like triacylglycerol yields from plant leaves. Plant Biotechnol. J. 2014, 12, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Andrianov, V.; Borisjuk, N.; Pogrebnyak, N.; Brinker, A.; Dixon, J.; Spitsin, S.; Flynn, J.; Matyszczuk, P.; Andryszak, K.; Laurelli, M.; et al. Tobacco as a production platform for biofuel: Overexpression of Arabidopsis DGAT and LEC2 genes increases accumulation and shifts the composition of lipids in green biomass. Plant Biotechnol. J. 2010, 8, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Usta, N. Use of tobacco seed oil methyl ester in a turbocharged indirect injection diesel engine. Biomass Bioenerg. 2005, 28, 77–86. [Google Scholar] [CrossRef]
- Zhang, L.; Joshi, A.K.; Hofmann, J.; Schweizer, E.; Smith, S. Cloning; expression; and characterization of the human mitochondrial β-ketoacyl synthase. J. Biol. Chem. 2005, 280, 12422–12429. [Google Scholar] [CrossRef] [PubMed]
- Kachroo, A.; Lapchyk, L.; Fukushige, H.; Hildebrand, D.; Klessig, D.; Kachroo, P. Plastidial fatty acid signaling modulates salicylic acid and jasmonic acid mediated defense pathways in the Arabidopsis ssi2 mutant. Plant Cell 2003, 15, 2952–2965. [Google Scholar] [CrossRef] [PubMed]
- Kachroo, A.; Venugopal, S.C.; Lapchyk, L.; Falcone, D.; Hildebrand, D.; Kachroo, P. Oleic acid levels regulated by glycerolipid metabolism modulate defense gene expression in Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 5152–5157. [Google Scholar] [CrossRef] [PubMed]
- Nandi, A.; Krothapalli, K.; Buseman, C.M.; Li, M.; Welti, R.; Enyedi, A.; Shah, J. Arabidopsis sfd mutants affect plastidic lipid composition and suppress dwarfing; cell death; and the enhanced disease resistance phenotypes resulting from the deficiency of a fatty acid desaturase. Plant Cell 2003, 15, 2383–2398. [Google Scholar] [CrossRef] [PubMed]
- Ohlrogge, J.; Browse, J. Lipid biosynthesis. Plant Cell 1995, 7, 957–970. [Google Scholar] [CrossRef] [PubMed]
- Somerville, C.; Browse, J.; Jaworski, J.G.; Ohlrogge, J. Lipids. In Biochemistry & Molecular Biology of Plants; Buchanan, B.B., Gruissem, W., Jones, R.L., Eds.; Amercian Society of Plant Physiologists: Rockville, MD, USA, 2000; pp. 456–527. [Google Scholar]
- Qin, Y.M.; Hu, C.Y.; Pang, Y.; Kastaniotis, A.J.; Hiltunen, J.K.; Zhu, Y.X. Saturated very-long-chain fatty acids promote cotton fiber and Arabidopsis cell elongation by activating ethylene biosynthesis. Plant Cell 2007, 19, 3692–3704. [Google Scholar] [CrossRef] [PubMed]
- Post-Beittenmiller, D.; Jaworski, J.G.; Ohlrogge, J.B. In vivo pools of free and acylated acyl carrier proteins in spinach. Evidence for sites of regulation of fatty acid biosynthesis. J. Biol. Chem. 1991, 266, 1858–1865. [Google Scholar] [PubMed]
- Post-Beittenmiller, D.; Roughan, R.G.; Ohlrogge, J.B. Regulation of plant fatty acid biosynthesis: Analysis of acyl-CoA and acyl-ACP substrate pools in spinach and pea chloroplasts. Plant Physiol. 1992, 100, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Lee, H.; Kang, S.; Park, W.J. Fatty acid desaturases, polyunsaturated fatty acid regulation and biotechnological advances. Nutrients 2016, 8, 23. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Lopez, N.; Usher, S.; Sayanova, O.V.; Napier, J.A.; Haslam, R.P. Modifying the lipid content and composition of plant seeds: Engineering the production of LC–PUFA. Appl. Microbiol. Biotechnol. 2015, 99, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.H.; Yang, T.Q.; Wang, R.L.; Liu, A.Z. Characterisation of DGAT1 and DGAT2 from Jatropha curcas and their functions in storage lipid biosynthesis. Funct. Plant. Biol. 2014, 41, 321–329. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. ClustalW and ClustalX version 2. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood; evolutionary distance; and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Songkumarn, P.; Liu, J.; Wang, G.L. A versatile zero background T-vector system for gene cloning and functional genomics. Plant Physiol. 2009, 150, 1111–1121. [Google Scholar] [CrossRef] [PubMed]
- Horsch, R.B.; Fry, J.E.; Hoffmann, N.; Eicholz, D.; Rogers, S.G.; Fraley, R.T. A simple and general method for transferring genes into plants. Science 1985, 227, 1229–1231. [Google Scholar]
- Pyke, K.A.; Leech, R.M. Rapid image analysis screening procedure for identifying chloroplast number mutants in mesophyll cells of Arabidopsis thaliana (L.) Heynh. Plant Physiol. 1991, 96, 1193–1195. [Google Scholar] [CrossRef] [PubMed]
- Arnon, D.I. Copper enzymes in isolation chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Bligh, E.G.; Dyer, W.J. A rapid method for total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Maisonneuve, S.; Bessoule, J.J.; Lessire, R.; Delseny, M.; Roscoe, T.J. Expression of rapeseed microsomal lysophosphatidic acid acyltransferase isozymes enhances seed oil content in Arabidopsis. Plant Physiol. 2010, 152, 670–684. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FA Species | WT | KASI-1OE | KASI-2OE | kasI-1 RNAi | kasI-2 RNAi | kasI-1/2 RNAi |
|---|---|---|---|---|---|---|
| 10C | 6.07 ± 0.48 | 4.66 ± 0.66 | 4.33 ± 0.65 | 6.56 ± 0.40 | 5.29 ± 0.28 | 4.39 ± 0.41 |
| 12C | 3.31 ± 0.35 | 2.77 ± 0.35 | 2.44 ± 0.51 | 3.13 ± 0.75 | 3.34 ± 0.25 | 11.06 ± 0.61 |
| 14C | 1.71 ± 0.56 | 1.67 ± 0.54 | 2.43 ± 0.60 | 1.81 ± 0.67 | 2.34 ± 0.11 | 1.51 ± 0.20 |
| Total | 11.09 | 9.10 | 9.20 | 11.50 | 10.97 | 16.96 ** |
| 16C | 11.07 ± 1.30 | 13.66 ± 1.83 | 11.66 ± 1.08 | 11.43 ± 0.18 | 11.32 ± 0.49 | 11.97 ± 2.38 |
| 16C1 | 2.80 ± 0.16 | 2.47 ± 0.70 | 2.08 ± 0.24 | 2.93 ± 0.13 | 2.56 ± 0.34 | 1.16 ± 0.31 |
| 16C2 | 8.88 ± 1.44 | 8.34 ± 0.12 | 8.72 ± 0.50 | 9.02 ± 0.14 | 8.66 ± 1.20 | 6.51 ± 0.23 |
| 16C3 | 2.14 ± 0.22 | 3.32 ± 0.30 | 2.90 ± 0.27 | 2.26 ± 0.16 | 2.05 ± 0.52 | 1.10 ± 0.10 |
| 18C | 2.12 ± 0.22 | 3.76 ± 0.32 | 2.98 ± 0.28 | 2.63 ± 0.15 | 2.45 ± 0.14 | 3.07 ± 0.49 |
| 18C1 | 3.73 ± 0.13 | 4.73 ± 0.24 | 4.10 ± 0.20 | 4.19 ± 0.14 | 4.06 ± 0.19 | 4.51 ± 0.43 |
| 18C2 | 11.48 ± 1.26 | 10.49 ± 0.34 | 12.57 ± 0.25 | 12.76 ± 0.16 | 12.36 ± 0.90 | 14.04 ± 3.55 |
| 18C3 | 43.38 ± 1.34 | 43.50 ± 1.11 | 44.35 ± 1.31 | 38.63 ± 1.95 | 43.86 ± 1.29 | 35.98 ± 2.52 |
| Total | 85.60 | 90.27 * | 89.36 | 83.85 | 87.32 | 78.34 ** |
| 20C | 2.74 ± 0.51 | 1.45 ± 0.60 ** | 1.43 ± 0.22 ** | 5.55 ± 0.19 ** | 1.71 ± 0.61 * | 4.7 ± 0.75 ** |
| US/S | 2.7 | 2.6 | 3.0 | 2.2 | 2.7 | 1.7 |
| FA content | 0.96 ± 0.13 | 1.03 ± 0.09 | 0.76 ± 0.05 ** | 0.83 ± 0.03 * | 0.88 ± 0.07 | 0.76 ± 0.07 ** |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, T.; Xu, R.; Chen, J.; Liu, A. β-Ketoacyl-acyl Carrier Protein Synthase I (KASI) Plays Crucial Roles in the Plant Growth and Fatty Acids Synthesis in Tobacco. Int. J. Mol. Sci. 2016, 17, 1287. https://doi.org/10.3390/ijms17081287
Yang T, Xu R, Chen J, Liu A. β-Ketoacyl-acyl Carrier Protein Synthase I (KASI) Plays Crucial Roles in the Plant Growth and Fatty Acids Synthesis in Tobacco. International Journal of Molecular Sciences. 2016; 17(8):1287. https://doi.org/10.3390/ijms17081287
Chicago/Turabian StyleYang, Tianquan, Ronghua Xu, Jianghua Chen, and Aizhong Liu. 2016. "β-Ketoacyl-acyl Carrier Protein Synthase I (KASI) Plays Crucial Roles in the Plant Growth and Fatty Acids Synthesis in Tobacco" International Journal of Molecular Sciences 17, no. 8: 1287. https://doi.org/10.3390/ijms17081287
APA StyleYang, T., Xu, R., Chen, J., & Liu, A. (2016). β-Ketoacyl-acyl Carrier Protein Synthase I (KASI) Plays Crucial Roles in the Plant Growth and Fatty Acids Synthesis in Tobacco. International Journal of Molecular Sciences, 17(8), 1287. https://doi.org/10.3390/ijms17081287