
Melatonin Alleviates Liver Apoptosis in Bile Duct Ligation Young Rats


 , ,
, , 
Abstract
:
1. Introduction
2. Results
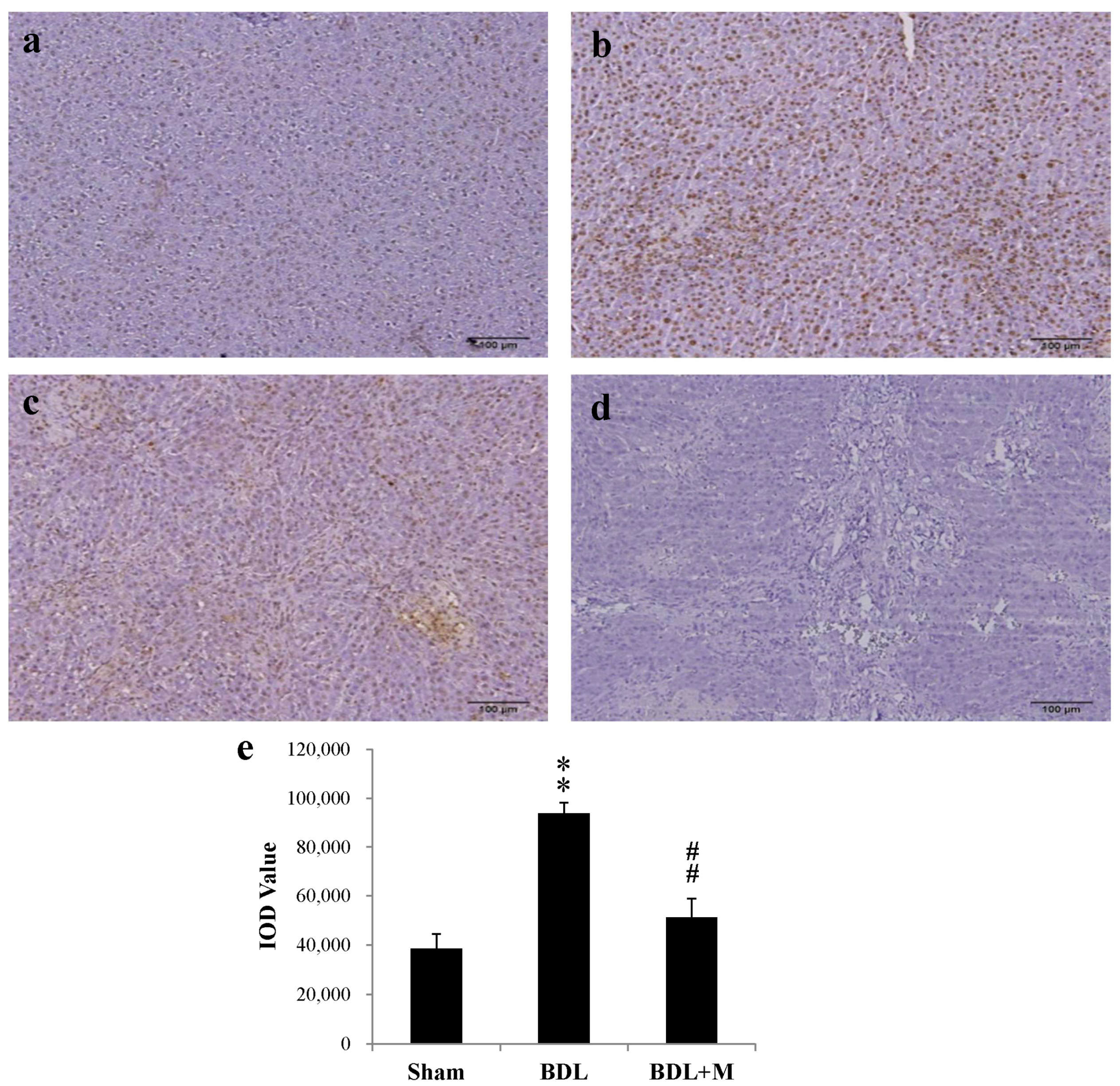
2.1. Bile Duct Ligation (BDL) Resulted in Increased Hepatic Apoptosis
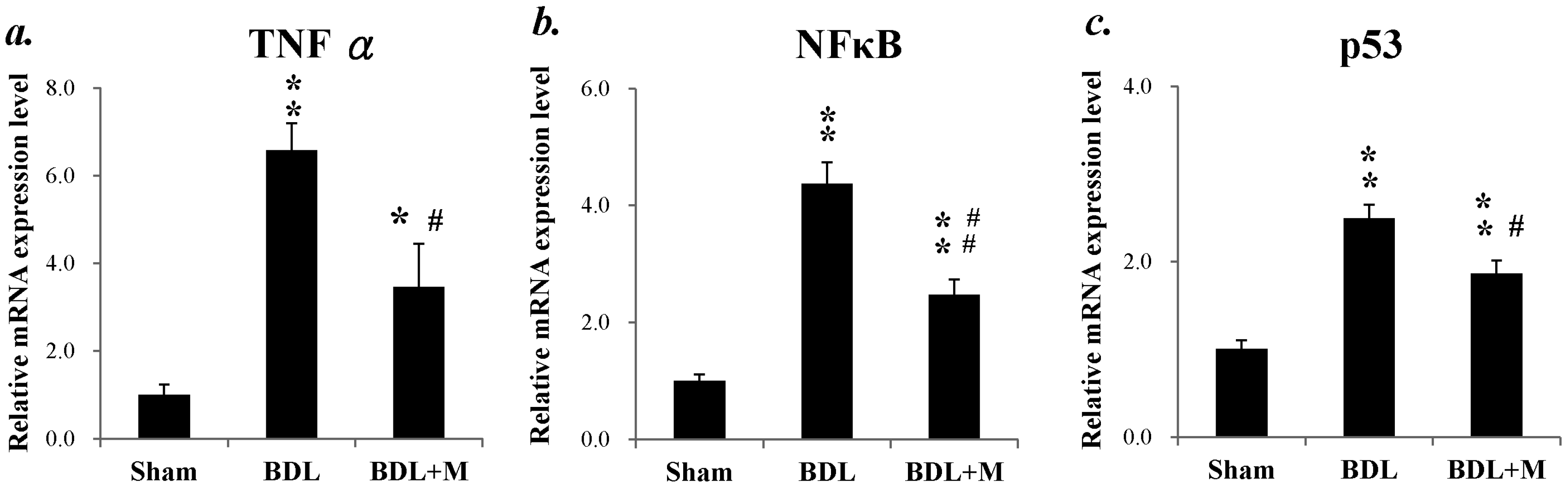
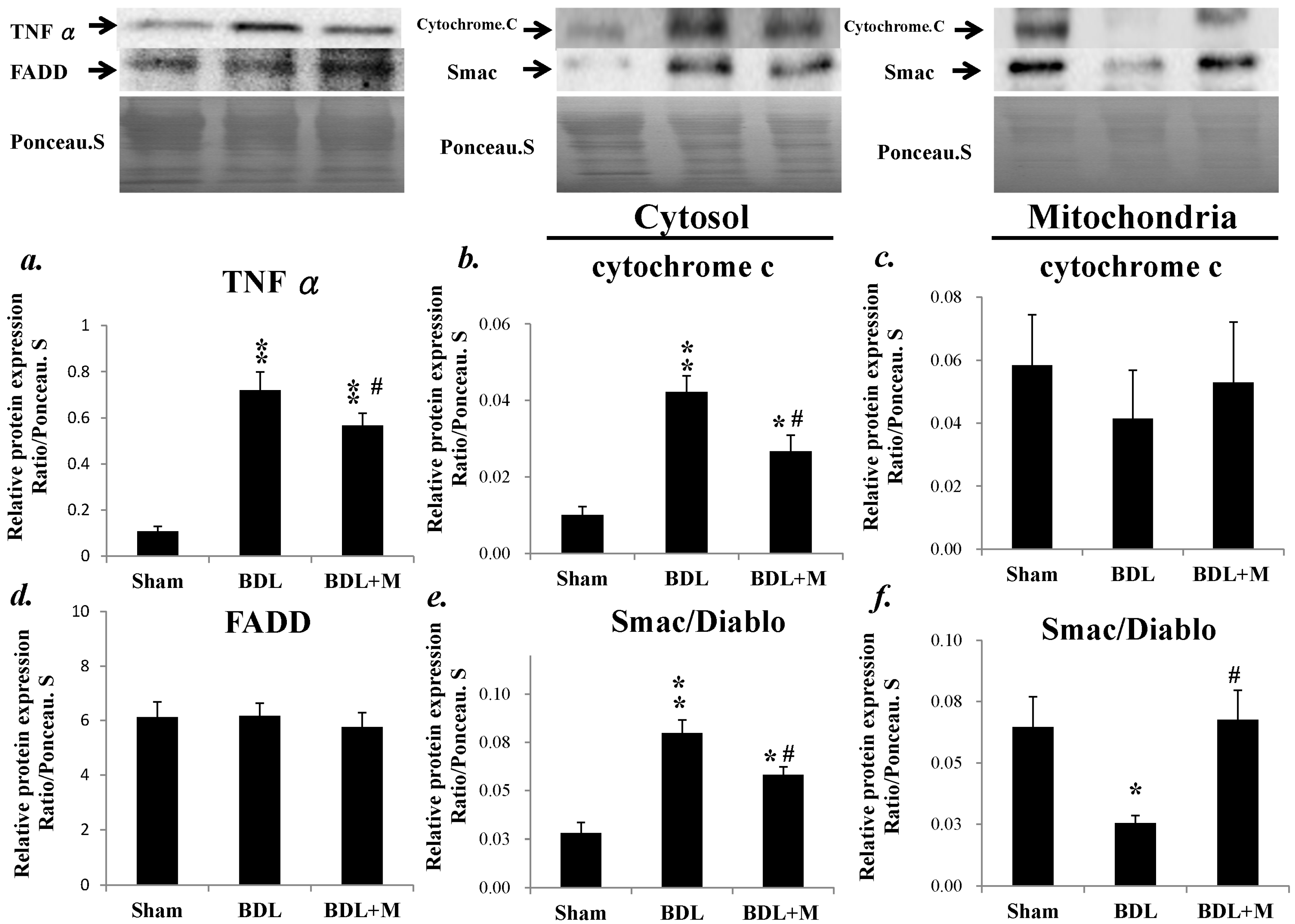
2.2. BDL Increased the Mrna Expression of Proinflammatory Mediators and Melatonin Treatment Altered the Changes
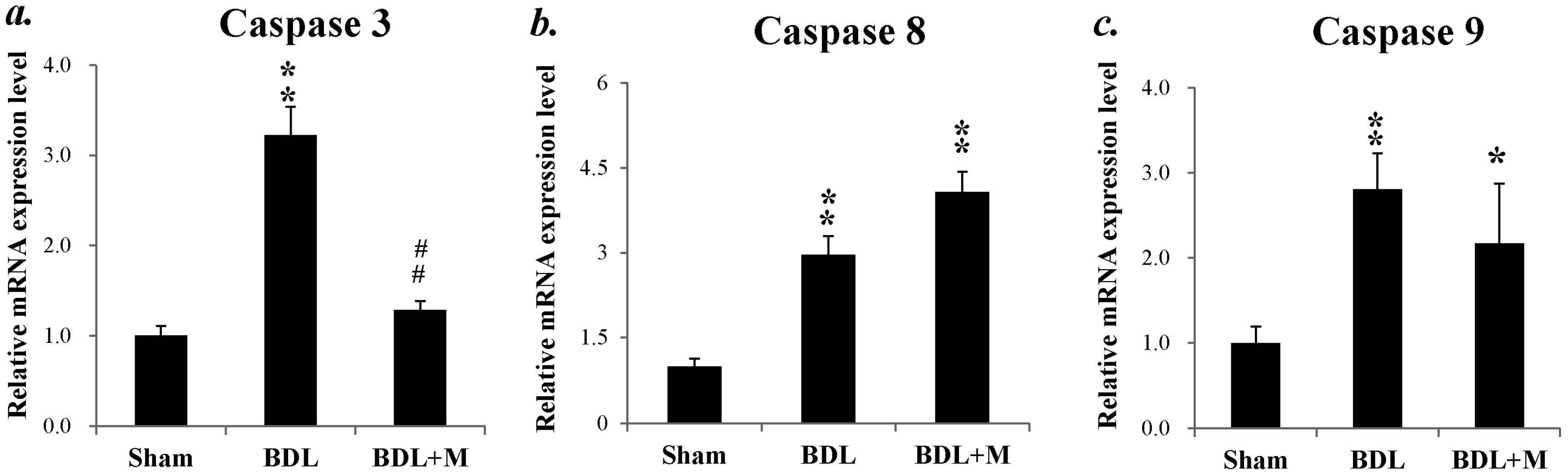
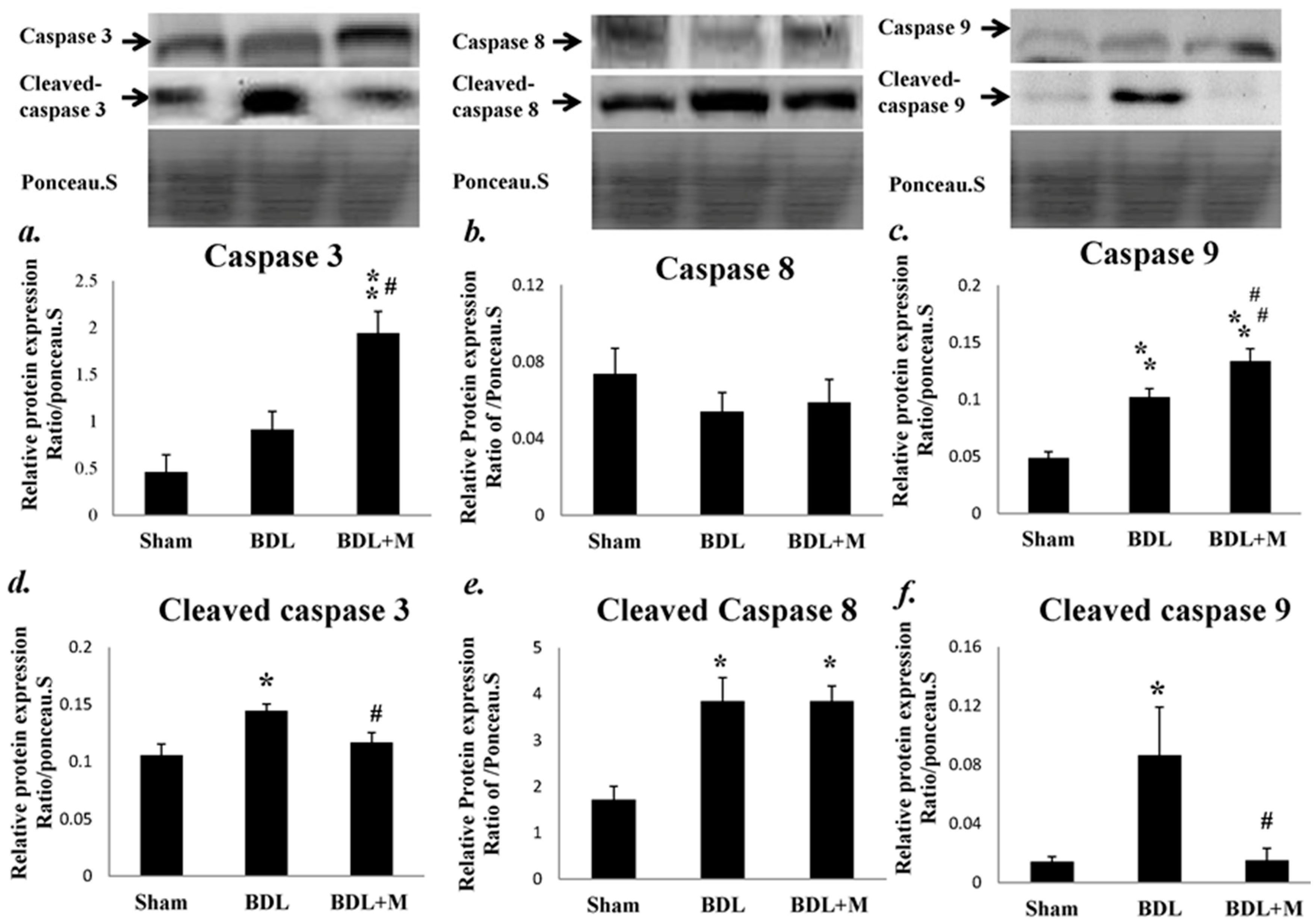
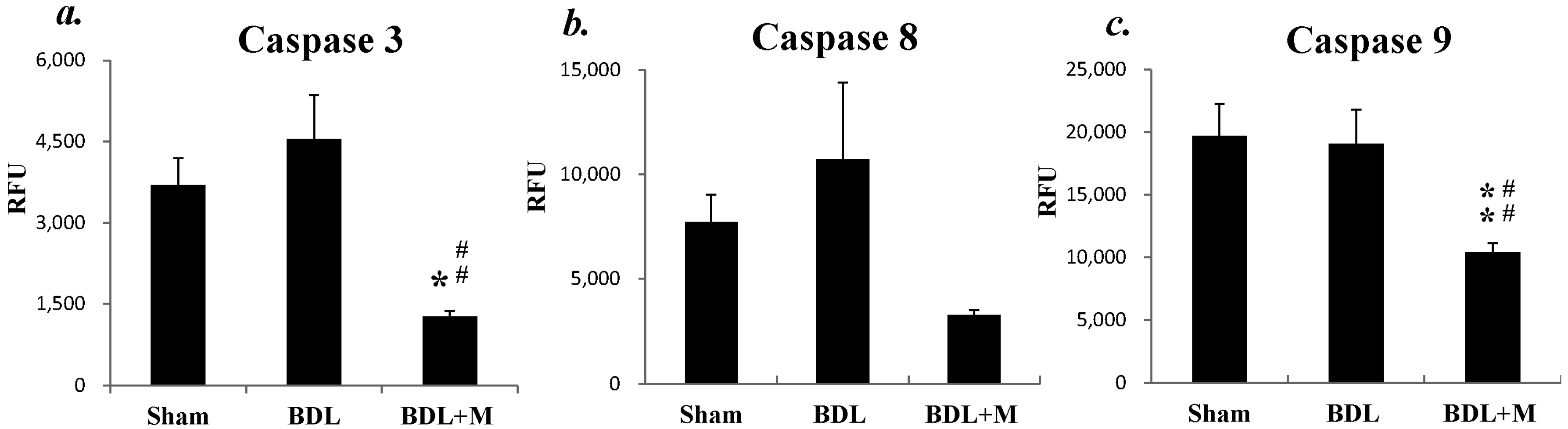
2.3. BDL Induced Liver Apoptosis via the Caspase-Dependent Pathway
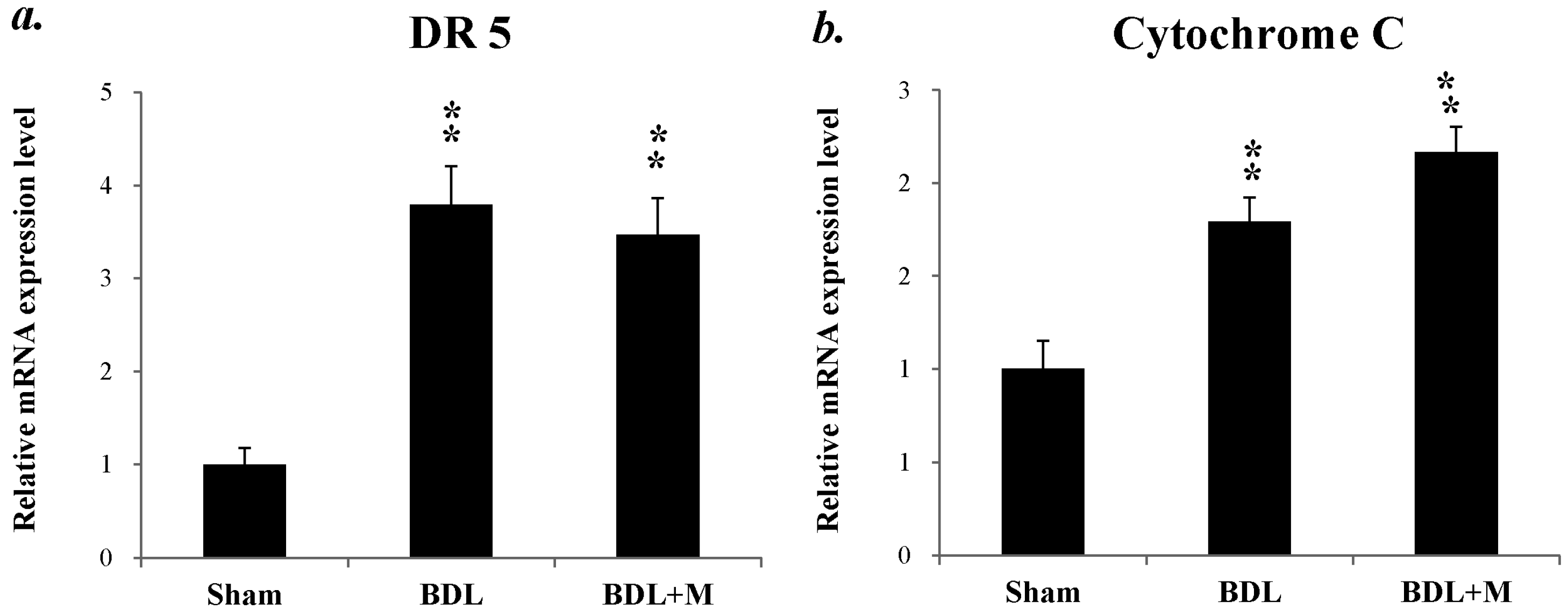
2.4. Melatonin Blocked the Apoptotic Signal Mediation Mainly through the Intrinsic Pathway Induced by BDL
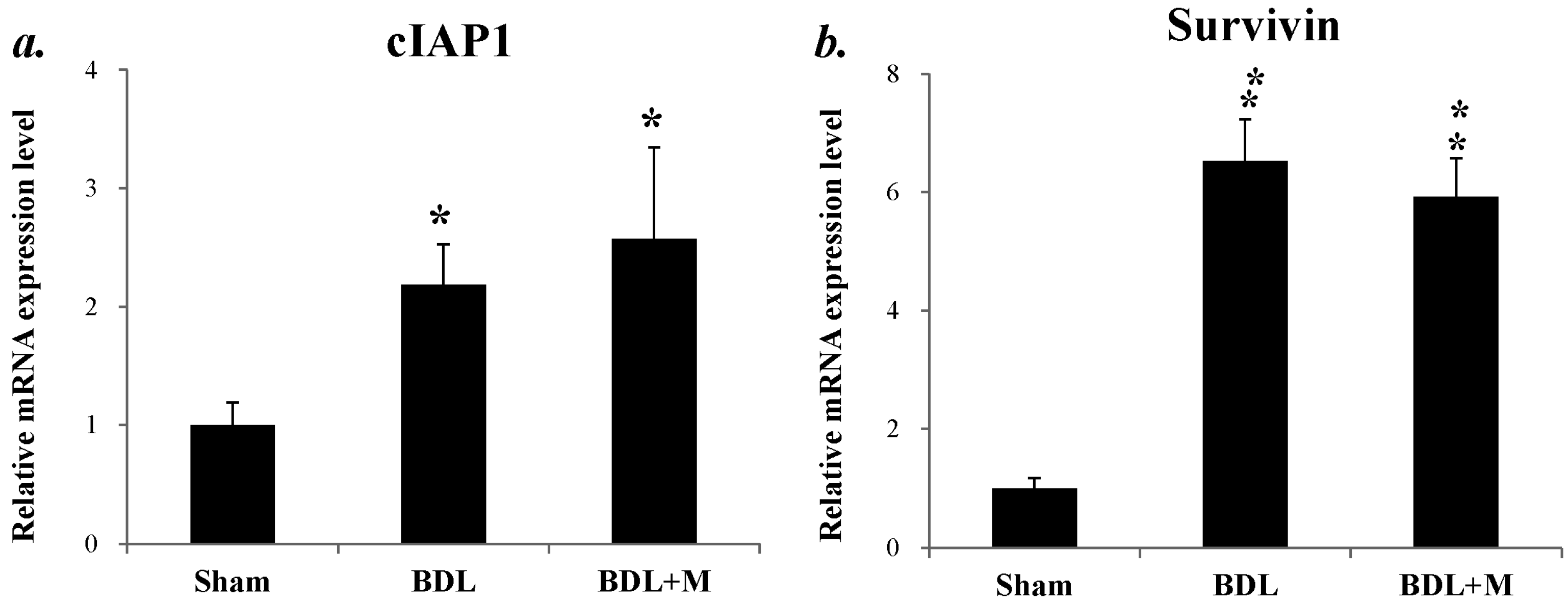
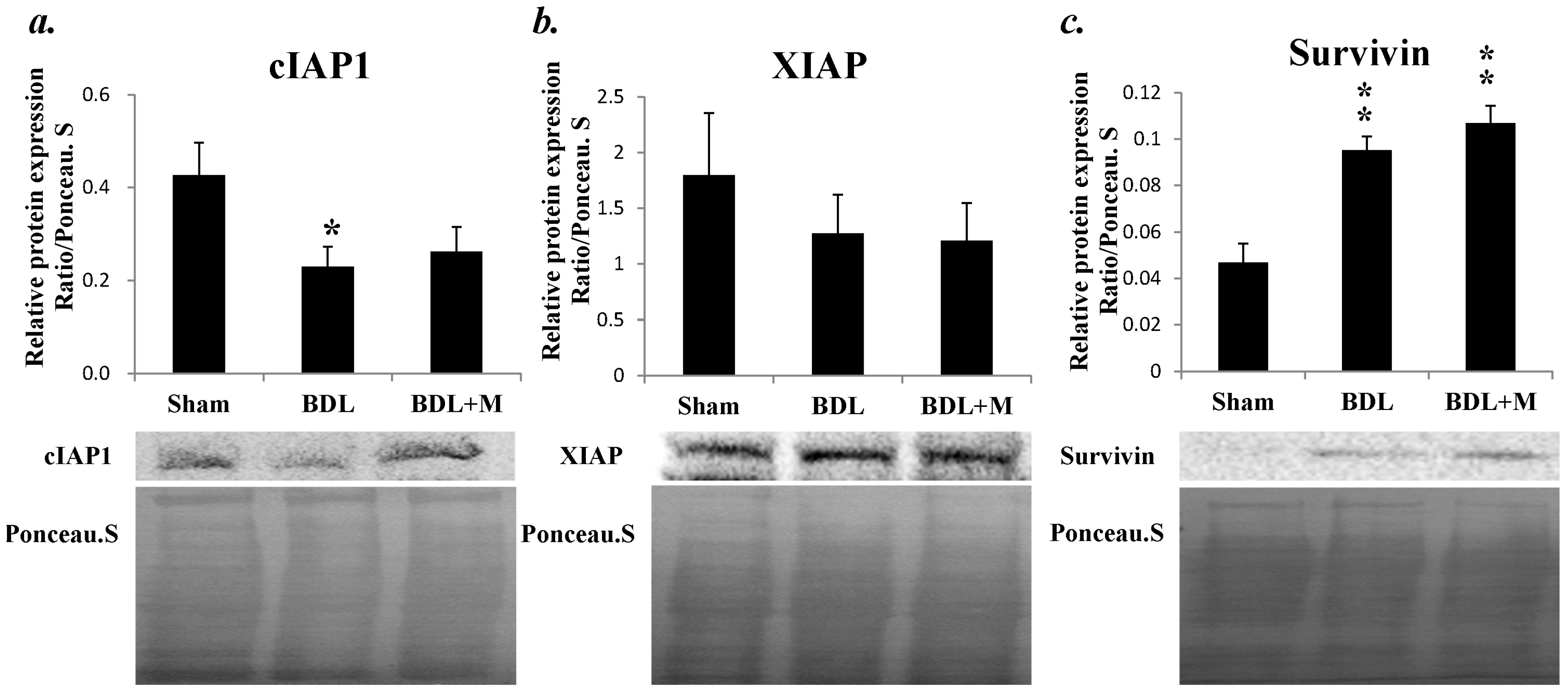
2.5. Anti-Apoptotosis Factors Change in Response to BDL and Melatonin Treatment
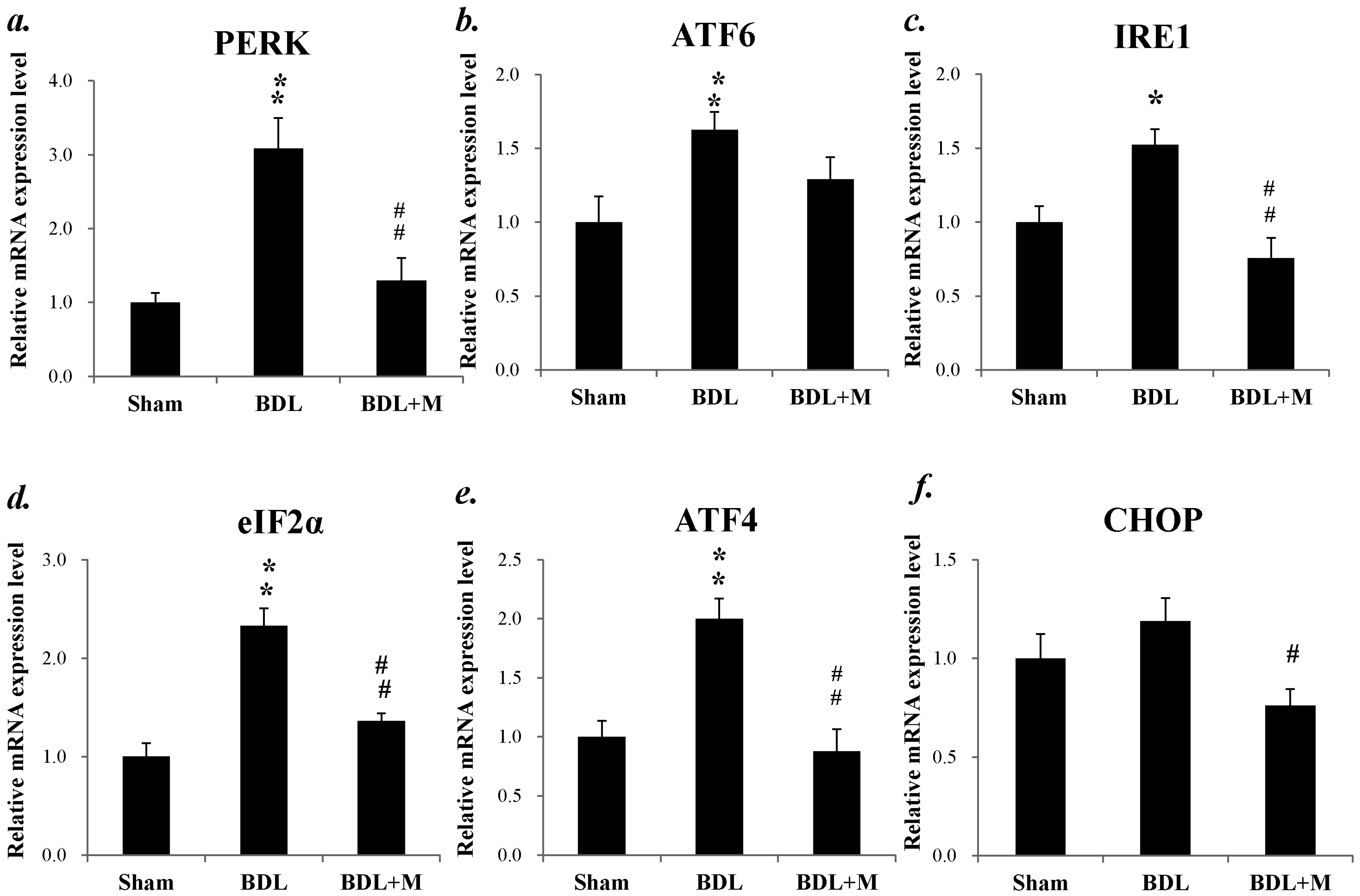
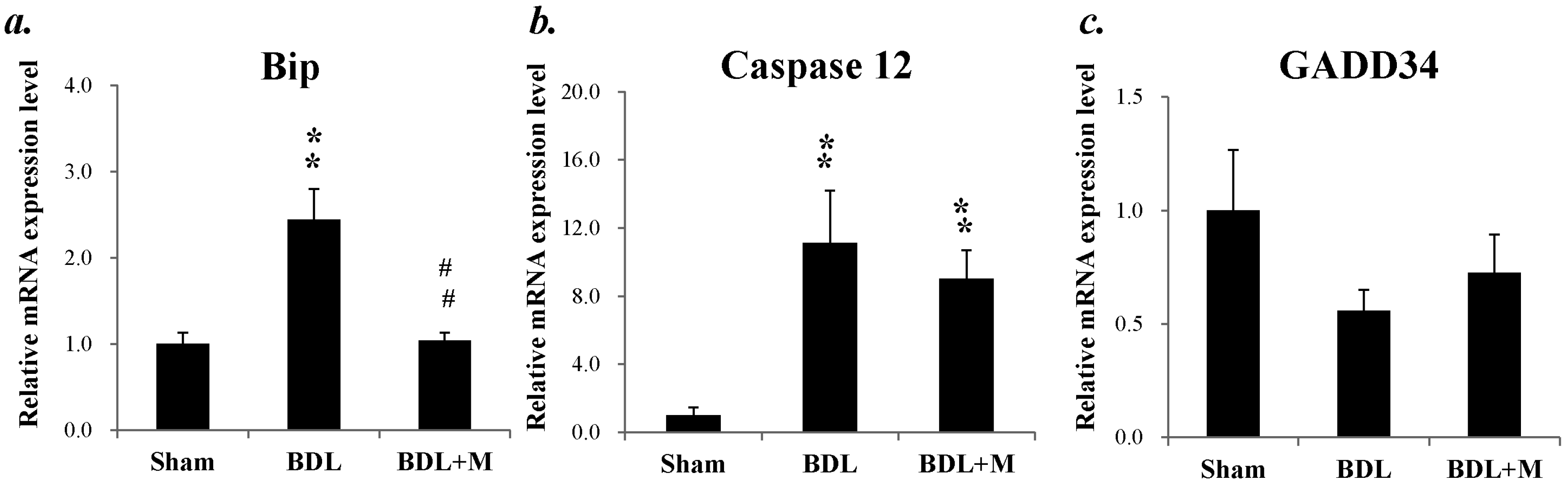
2.6. Endoplasmic Reticulum (ER) Stress in BDL Rat
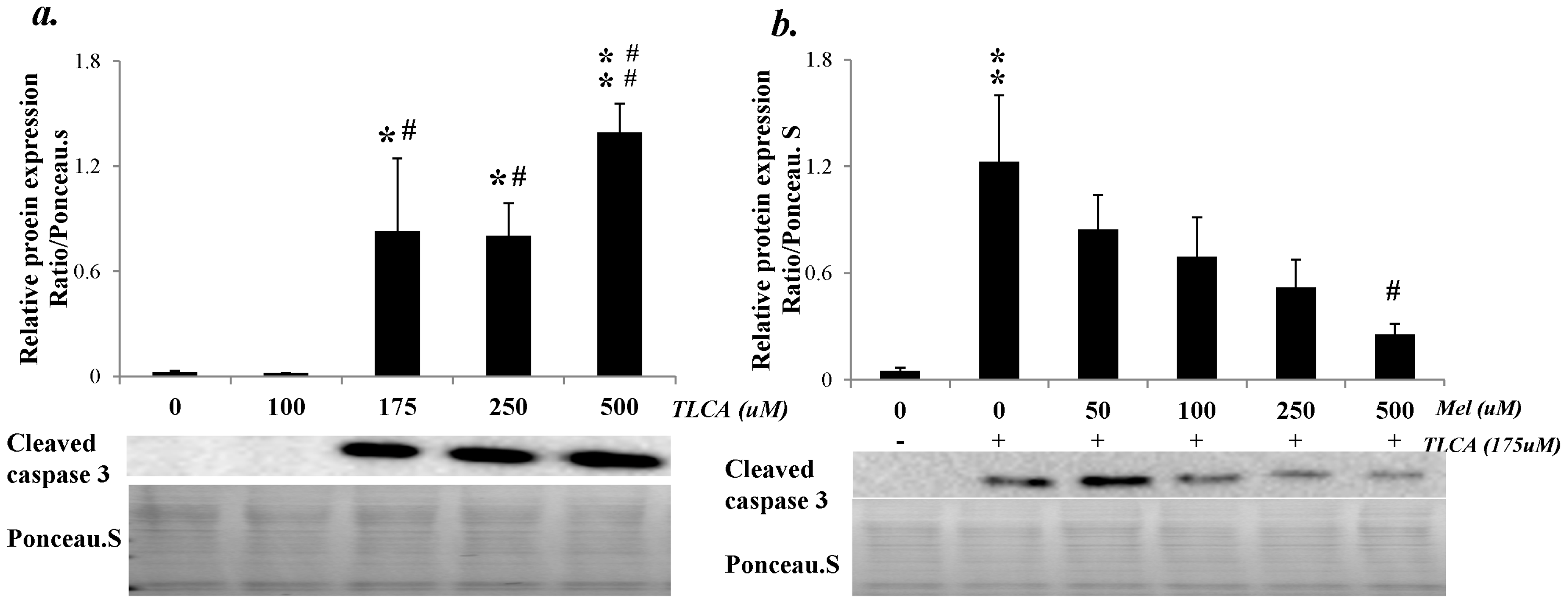
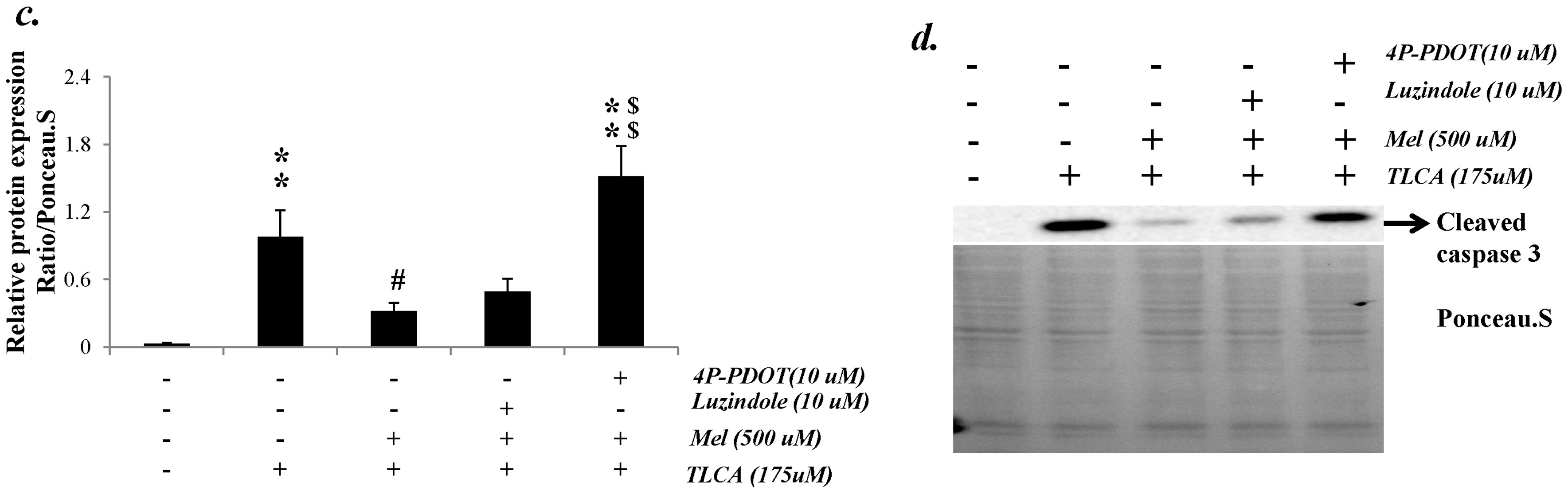
2.7. Melatonin Reduced Cleaved Caspase 3 Expression through the Melatonin 2 (MT2) Receptor
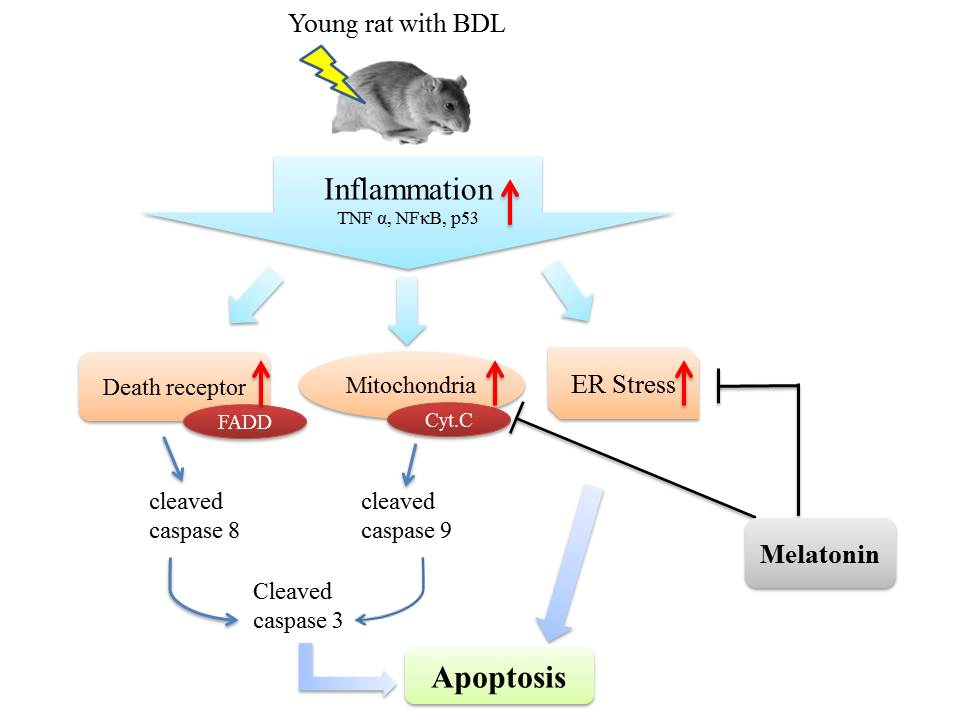
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Experimental Procedures
4.3. Plasma Biochemistry Parameters Measurement
4.4. Liver Histopathology
4.5. Western Blot
4.6. Quantitative Real-Time Polymerase Chain Reaction (PCR) analysis
4.7. HepG2 Cell Culture
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Huang, L.T.; Tiao, M.M.; Tain, Y.L.; Chen, C.C.; Hsieh, C.S. Melatonin ameliorates bile duct ligation-induced systemic oxidative stress and spatial memory deficits in developing rats. Pediatr. Res. 2009, 65, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Celik, T.; Gören, M.Z.; Cinar, K.; Gürdal, H.; Onder, F.O.; Tan, A.; Terzioğlu, B.; Bozdayi, A.M.; Bozkaya, H.; Uzunalimoğlu, O.; et al. Fatigue of cholestasis and the serotoninergic neurotransmitter system in the rat. Hepatology 2005, 41, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Magen, I.; Avraham, Y.; Ackerman, Z.; Vorobiev, L.; Mechoulam, R.; Berry, E.M. Cannabidiol ameliorates cognitive and motor impairments in bile-duct ligated mice via 5-HT1A receptor activation. Br. J. Pharmacol. 2010, 159, 950–957. [Google Scholar] [CrossRef] [PubMed]
- Sheen, J.M.; Huang, L.T.; Hsieh, C.S.; Chen, C.C.; Wang, J.Y.; Tain, Y.L. Bile duct ligation in developing rats: Temporal progression of liver, kidney, and brain damage. J. Pediatr. Surg. 2010, 45, 1650–1658. [Google Scholar] [CrossRef] [PubMed]
- Georgiev, P.; Jochum, W.; Heinrich, S.; Jang, J.H.; Nocito, A.; Dahm, F.; Clavien, P.A. Characterization of time-related changes after experimental bile duct ligation. Br. J. Surg. 2008, 95, 646–656. [Google Scholar] [CrossRef] [PubMed]
- Marques, T.G.; Chaib, E.; da Fonseca, J.H.; Lourenço, A.C.; Silva, F.D.; Ribeiro, M.A., Jr.; Galvão, F.H.; D’Albuquerque, L.A. Review of experimental models for inducing hepatic cirrhosis by bile duct ligation and carbon tetrachloride injection. Acta Cir. Bras. 2012, 27, 589–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.C.; Sheen, J.M.; Tain, Y.L.; Chen, C.C.; Tiao, M.M.; Huang, Y.H.; Hsieh, C.S.; Huang, L.T. Alterations in NADPH oxidase expression and blood-brain barrier in bile duct ligation-treated young rats: Effects of melatonin. Neurochem. Int. 2012, 60, 751–758. [Google Scholar] [CrossRef] [PubMed]
- D’Mello, C.; Le, T.; Swain, M.G. Cerebral microglia recruit monocytes into the brain in response to tumor necrosis factor alpha signaling during peripheral organ inflammation. J. Neurosci. 2009, 29, 2089–2102. [Google Scholar] [CrossRef] [PubMed]
- Sheen, J.M.; Chen, Y.C.; Tain, Y.L.; Huang, L.T. Increased circulatory asymmetric dimethylarginine and multiple organ failure: Bile duct ligation in rat as a model. Int. J. Mol. Sci. 2014, 15, 3989–4006. [Google Scholar] [CrossRef] [PubMed]
- Orellana, M.; Rodrigo, R.; Thielemann, L.; Guajardo, V. Bile duct ligation and oxidative stress in the rat: Effects in liver and kidney. Comp. Biochem. Physiol. C 2000, 126, 105–111. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Aktas, C.; Kanter, M.; Erboga, M.; Mete, R.; Oran, M. Melatonin attenuates oxidative stress, liver damage and hepatocyte apoptosis after bile-duct ligation in rats. Toxicol. Ind. Health 2014, 30, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Eckert, A.; Keil, U.; Kressmann, S.; Schindowski, K.; Leutner, S.; Leutz, S.; Müller, W.E. Effects of EGb 761 Ginkgo biloba extract on mitochondrial function and oxidative stress. Pharmacopsychiatry 2003, 36, 15–23. [Google Scholar]
- Agil, A.; El-Hammadi, M.; Jiménez-Aranda, A.; Tassi, M.; Abdo, W.; Fernández-Vázquez, G.; Reiter, R.J. Melatonin reduces hepatic mitochondrial dysfunction in diabetic obese rats. J. Pineal Res. 2015, 59, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Sarkar, J.; Haldar, C.; Sinha, S. Melatonin Reverses Fas, E2F-1 and Endoplasmic Reticulum Stress Mediated Apoptosis and Dysregulation of Autophagy Induced by the Herbicide Atrazine in Murine Splenocytes. PLoS ONE 2014, 9, e108602. [Google Scholar] [CrossRef] [PubMed]
- Puri, P.; Mirshahi, F.; Cheung, O.; Natarajan, R.; Maher, J.W.; Kellum, J.M.; Sanyal, A.J. Activation and dysregulation of the unfolded protein response in nonalcoholic fatty liver disease. Gastroenterology 2008, 134, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Henkel, A.S.; Elias, M.S.; Green, R.M. Homocysteine supplementation attenuates the unfolded protein response in a murine nutritional model of steatohepatitis. J. Biol. Chem. 2009, 284, 31807–31816. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Kang, H.S.; Lee, J.H.; Park, J.H.; Jung, C.H.; Bae, J.H.; Oh, B.C.; Song, D.K.; Baek, W.K.; Im, S.S. Melatonin ameliorates ER stress-mediated hepatic steatosis through miR-23a in the liver. Biochem. Biophys. Res. Commun. 2015, 458, 462–469. [Google Scholar] [CrossRef] [PubMed]
- San-Miguel, B.; Crespo, I.; Sánchez, D.I.; González-Fernández, B.; Ortiz de Urbina, J.J.; Tuñón, M.J.; González-Gallego, J. Melatonin inhibits autophagy and endoplasmic reticulum stress in mice with carbon tetrachloride-induced fibrosis. J. Pineal Res. 2015, 59, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Manchester, L.C.; Coto-Montes, A.; Boga, J.A.; Andersen, L.P.; Zhou, Z.; Galano, A.; Vriend, J.; Schliess, F.; Görg, B.; Häussinger, D. Pathogenetic interplay between osmotic and oxidative stress: The hepatic encephalopathy paradigm. Biol. Chem. 2006, 387, 1363–1370. [Google Scholar]
- Esteban-Zubero, E.; Alatorre-Jiménez, M.A.; López-Pingarrón, L.; Reyes-Gonzales, M.C.; Almeida-Souza, P.; Cantín-Golet, A.; Ruiz-Ruiz, F.J.; Tan, D.X.; García, J.J.; Reiter, R.J. Melatonin’s role in preventing toxin-related and sepsis-mediated hepatic damage: A review. Pharmacol. Res. 2016, 105, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Acuña Castroviejo, D.; López, L.C.; Escames, G.; López, A.; García, J.A.; Reiter, R.J. Melatonin-mitochondria interplay in health and disease. Curr. Top. Med. Chem. 2011, 11, 221–240. [Google Scholar] [CrossRef] [PubMed]
- Mathes, A.M. Hepatoprotective actions of melatonin: Possible mediation by melatonin receptors. World. J. Gastroenterol. 2010, 16, 6087–6097. [Google Scholar] [CrossRef] [PubMed]
- Tahan, G.; Akin, H.; Aydogan, F.; Ramadan, S.S.; Yapicier, O.; Tarcin, O.; Uzun, H.; Tahan, V.; Zengin, K. Melatonin ameliorates liver fibrosis induced by bile-duct ligation in rats. Can. J. Surg. 2010, 53, 313–318. [Google Scholar] [PubMed]
- Tain, Y.L.; Hsieh, C.S.; Chen, C.C.; Sheen, J.M.; Lee, C.T.; Huang, L.T. Melatonin prevents increased asymmetric dimethylarginine in young rats with bile duct ligation. J. Pineal Res. 2010, 48, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Tiao, M.M.; Huang, L.T.; Chen, C.J.; Sheen, J.M.; Tain, Y.L.; Chen, C.C.; Kuo, H.C.; Huang, Y.H.; Tang, K.S.; Chu, E.W.; et al. Melatonin in the regulation of liver steatosis following prenatal glucocorticoid exposure. BioMed Res. Int. 2014, 2014, 942172. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.Y.; Wei, J.R.; Zhang, R.; Ye, X.L.; Zhu, Y.F.; Li, W.J. Effect of glutamine on caspase-3 mRNA and protein expression in the myocardium of rats with sepsis. Am. J. Med. Sci. 2014, 348, 315–318. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Meng, Z. Expression of caspase and apoptotic signal pathway induced by sulfur dioxide. Environ. Mol. Mutagen. 2010, 51, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Dubocovich, M.L. Melatonin receptors: Are there multiple subtypes? Trends Pharmacol. Sci. 1995, 16, 50–56. [Google Scholar] [CrossRef]
- Morgan, P.J.; Barrett, P.; Howell, H.E.; Helliwell, R. Melatonin receptors: Localization, molecular pharmacology and physiological significance. Neurochem. Int. 1994, 24, 101–146. [Google Scholar] [CrossRef]
- Esrefoglu, M.; Gül, M.; Emre, M.H.; Polat, A.; Selimoglu, M.A. Protective effect of low dose of melatonin against cholestatic oxidative stress after common bile duct ligation in rats. World. J. Gastroenterol. 2005, 11, 1951–1956. [Google Scholar] [CrossRef] [PubMed]
- Emre, M.H.; Polat, A.; Eşrefoğlu, M.; Karabulut, A.B.; Gül, M. Effects of melatonin and acetylsalicylic acid against hepatic oxidative stress after bile duct ligation in rat. Acta Physiol. Hung. 2008, 95, 349–363. [Google Scholar] [CrossRef] [PubMed]
- Guha, M.; Maity, P.; Choubey, V.; Mitra, K.; Reiter, R.J.; Bandyopadhyay, U. Melatonin inhibits free radical-mediated mitochondrial-dependent hepatocyte apoptosis and liver damage induced during malarial infection. J. Pineal Res. 2007, 43, 372–381. [Google Scholar] [CrossRef] [PubMed]
- Padillo, F.J.; Cruz, A.; Navarrete, C.; Bujalance, I.; Briceño, J.; Gallardo, J.I.; Marchal, T.; Caballero, R.; Túnez, I.; Muntané, J.; et al. Melatonin prevents oxidative stress and hepatocyte cell death induced by experimental cholestasis. Free Radic. Res. 2004, 38, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Malhi, H.; Kaufman, R.J. Endoplasmic reticulum stress in liver disease. J. Hepatol. 2011, 54, 795–809. [Google Scholar] [CrossRef] [PubMed]
- Renzi, A.; Glaser, S.; Demorrow, S.; Mancinelli, R.; Meng, F.; Franchitto, A.; Venter, J.; White, M.; Francis, H.; Han, Y.; et al. Melatonin inhibits cholangiocyte hyperplasia in cholestatic rats by interaction with MT1 but not MT2 melatonin receptors. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Tain, Y.L.; Sheen, J.M.; Huang, L.T. Melatonin utility in neonates and children. J. Formos. Med. Assoc. 2012, 111, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, L.L.; Dammer, E.B.; Li, C.B.; Xu, G.; Chen, S.D.; Wang, G. Melatonin for sleep disorders and cognition in dementia: A meta-analysis of randomized controlled trials. Am. J. Alzheimer’s Dis. Other Demen. 2015, 30, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Gonciarz, M.; Gonciarz, Z.; Bielanski, W.; Mularczyk, A.; Konturek, P.C.; Brzozowski, T.; Konturek, S.J. The pilot study of 3-month course of melatonin treatment of patients with nonalcoholic steatohepatitis: Effect on plasma levels of liver enzymes, lipids and melatonin. J. Physiol. Pharmacol. 2010, 61, 705–710. [Google Scholar] [PubMed]
- Celinski, K.; Konturek, P.C.; Slomka, M.; Cichoz-Lach, H.; Brzozowski, T.; Konturek, S.J.; Korolczuk, A. Effects of treatment with melatonin and tryptophan on liver enzymes, parameters of fat metabolism and plasma levels of cytokines in patients with non-alcoholic fatty liver disease—14 months follow up. J. Physiol. Pharmacol. 2014, 65, 75–82. [Google Scholar] [PubMed]
- Tain, Y.L.; Chen, C.C.; Lee, C.T.; Kao, Y.H.; Sheen, J.M.; Yu, H.R.; Huang, L.T. Melatonin regulates l-arginine transport and NADPH oxidase in young rats with bile duct ligation: Role of protein kinase C. Pediatr. Res. 2013, 73, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Tiao, M.M.; Lin, T.K.; Chen, J.B.; Liou, C.W.; Wang, P.W.; Huang, C.C.; Chou, Y.M.; Huang, Y.H.; Chuang, J.H. Dexamethasone decreases cholestatic liver injury via inhibition of intrinsic pathway with simultaneous enhancement of mitochondrial biogenesis. Steroids 2011, 76, 660–666. [Google Scholar] [CrossRef] [PubMed]
- Wen, Q.; Liang, T.; Qin, F.; Wei, J.; He, Q.; Luo, X.; Chen, X.; Zheng, N.; Huang, R. Lyoniresinol 3α-O-β-d-glucopyranoside-mediated hypoglycaemia and its influence on apoptosis-regulatory protein expression in the injured kidneys of streptozotocin-induced mice. PLoS ONE 2013, 8, e81772. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Kao, Y.H.; Hsieh, C.S.; Chen, C.C.; Sheen, J.M.; Lin, I.C.; Huang, L.T. Melatonin blocks oxidative stress-induced increased asymmetric dimethylarginine. Free Radic. Biol. Med. 2010, 49, 1088–1098. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Sham (n = 10) | BDL (n = 10) | BDL + M (n = 10) |
|---|---|---|---|
| AST (IU/L) | 97.8 ± 3.6 | 411.5 ± 21.8 ** | 493.6 ± 40.2 ** |
| ALT (IU/L) | 34.7 ± 1.8 | 108.2 ± 6.7 ** | 99.8 ± 7.8 ** |
| Direct Bilirubin (mg/dL) | 0.17 ± 0.02 | 4.71 ± 0.32 ** | 3.63 ± 0.39 **,# |
| Total bilirubin (mg/dL) | 0.41 ± 0.28 | 5.63 ± 0.38 ** | 5.07 ± 0.50 ** |
| Gene | Forward 5′–3′ | Reverse 5′–3′ |
|---|---|---|
| Caspase-3 | GGCCGACTTCCTGTATGC | GCGCAAAGTGACTGGATG |
| Caspase-8 | ACGATATTGCTGAACGTCTGG | CCGACTGATATGGAAAAGCAG |
| Caspase-9 | GGAAGATCGAGAGACATGCAG | CCGTGACCATTTTCTTAGCAG |
| 18S | GCGATGCGGCGGCGTTAT | AGACTTTGGTTTCCCGGAAGC |
| Cytochrome c | AACCTCCATGGTCTGTTTGG | GTCTGCCCTTTCTCCCTTCT |
| cIAP1 | AGCTTGCAAGTGCTGGATTT | CTCCTGACCCTTCATCCGTA |
| Survivin | TGCAAAGGAGACCAACAACA | AAGCTGGGACAAGTGGCTTA |
| NFκB | GCTTACGGTGGGATTGCATT | GCACAATCTCTAGGCTCGTTTTTAA |
| TNFα | GGCTGCCCCGACTACGT | AGGGCAAGGGCTCTTGATG |
| DR5 | AAATGCTGCTGAAGTGGCT | ACTAATAAAGATCCTCTCGGCTC |
| BiP | GACCACCTATTCCTGCGTCGGT | CGCCAATCAGACGCTCCCCT |
| Caspase12 | GGAAGGTAGGCAAGAGT | GTAGAAGTAGCGTGTCATA |
| GADD34 | TGAATGTTGAGAGAAGAACC | TTGTTTAGAAGTCGCTCTG |
| PERK | GCTTGCTCCCACATCGGATA | TGCGGCAATTCGTCCATCTA |
| IRE1 | TTGACTATGCAGCCTCACTTC | AGTTACCACCAGTCCATCGC |
| eIF2 α | ATAGGCGTTTGACCCCACAA | ATCACATACCTGGGTGGAGC |
| ATF4 | CCTTCGACCAGTCGGGTTTG | CTGTCCCGGAAAAGGCATCC |
| ATF6 | AAGTGAAGAACCATTACTTTATATC | TTTCTGCTGGCTATTTGT |
| p53 | TATGACTTTAGGGCTTGTTA | AGCAACTACCAACCCATTC |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sheen, J.-M.; Chen, Y.-C.; Hsu, M.-H.; Tain, Y.-L.; Huang, Y.-H.; Tiao, M.-M.; Li, S.-W.; Huang, L.-T. Melatonin Alleviates Liver Apoptosis in Bile Duct Ligation Young Rats. Int. J. Mol. Sci. 2016, 17, 1365. https://doi.org/10.3390/ijms17081365
Sheen J-M, Chen Y-C, Hsu M-H, Tain Y-L, Huang Y-H, Tiao M-M, Li S-W, Huang L-T. Melatonin Alleviates Liver Apoptosis in Bile Duct Ligation Young Rats. International Journal of Molecular Sciences. 2016; 17(8):1365. https://doi.org/10.3390/ijms17081365
Chicago/Turabian StyleSheen, Jiunn-Ming, Yu-Chieh Chen, Mei-Hsin Hsu, You-Lin Tain, Ying-Hsien Huang, Mao-Meng Tiao, Shih-Wen Li, and Li-Tung Huang. 2016. "Melatonin Alleviates Liver Apoptosis in Bile Duct Ligation Young Rats" International Journal of Molecular Sciences 17, no. 8: 1365. https://doi.org/10.3390/ijms17081365
APA StyleSheen, J. -M., Chen, Y. -C., Hsu, M. -H., Tain, Y. -L., Huang, Y. -H., Tiao, M. -M., Li, S. -W., & Huang, L. -T. (2016). Melatonin Alleviates Liver Apoptosis in Bile Duct Ligation Young Rats. International Journal of Molecular Sciences, 17(8), 1365. https://doi.org/10.3390/ijms17081365