
Exposure to Zinc Sulfate Results in Differential Effects on Olfactory Sensory Neuron Subtypes in Adult Zebrafish

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
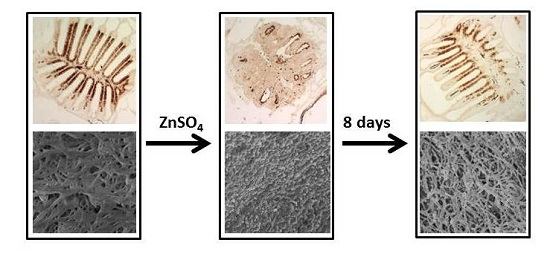
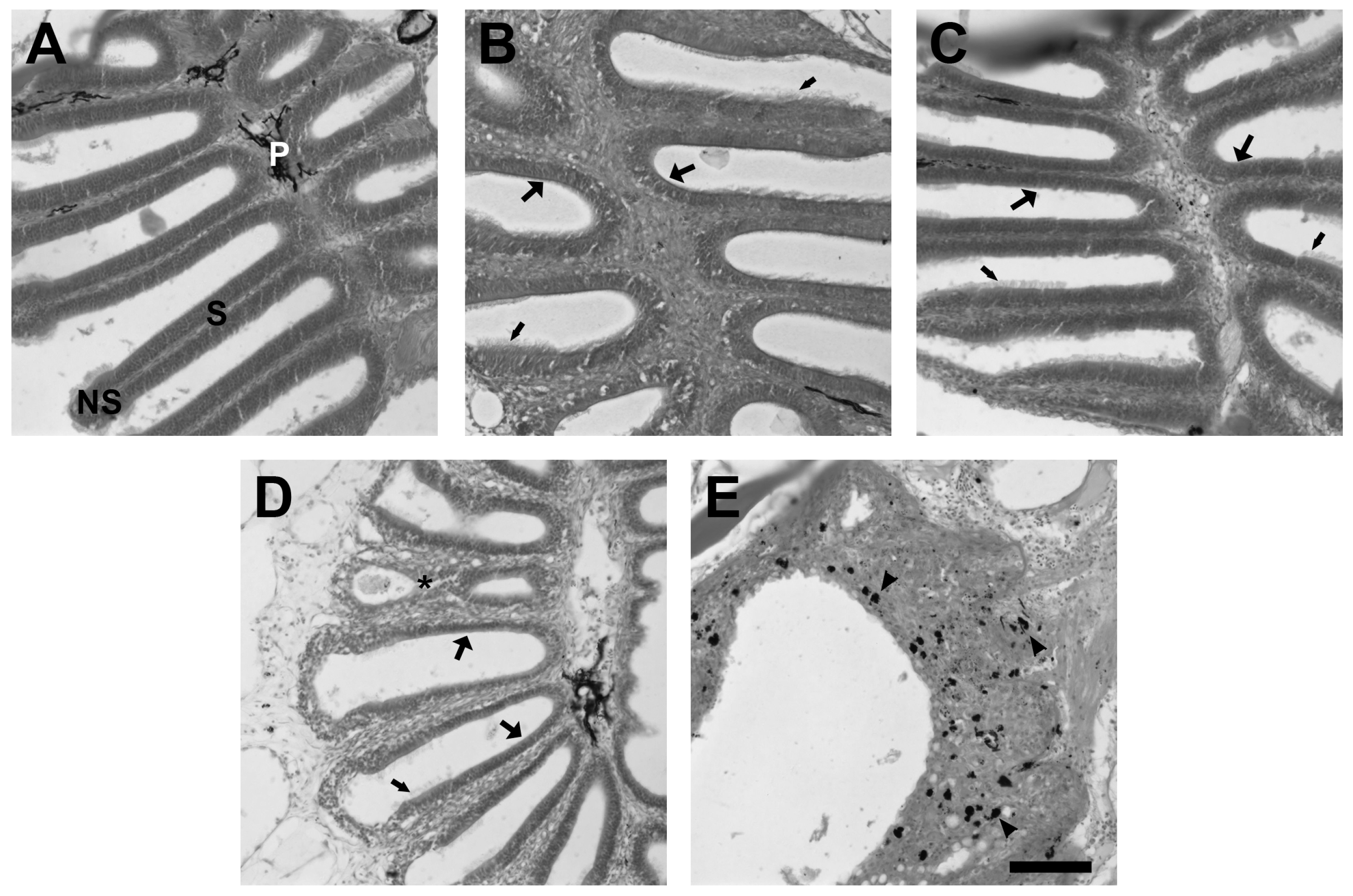
2.1. Time Course of Degeneration and Regeneration of the Zebrafish Olfactory Epithelium Following Exposure to Zinc Sulfate
2.2. Ultrastructural Analysis of Epithelial Surface with Scanning Electron Microscopy
2.3. Effects on Olfactory-Mediated Behavior
3. Discussion
4. Materials and Methods
4.1. Exposure to Zinc Sulfate
4.2. Tissue Processing
4.3. Histology and Immunohistochemistry
4.4. Optical Density Measurements
4.5. Scanning Electron Microscopy
4.6. Odor-Driven Behavior Assay
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Schwob, J.E. Neural regeneration and the peripheral olfactory system. Anat. Rec. 2002, 269, 33–49. [Google Scholar] [CrossRef] [PubMed]
- Zippel, H.P. Regeneration in the peripheral and central olfactory system: A review of morphological, physiological and behavioral aspects. J. Hirnforsch. 1993, 34, 207–229. [Google Scholar] [PubMed]
- Zupanc, G.K.H. Neurogenesis and neuronal regeneration in the adult fish brain. J. Comp. Physiol. A 2006, 192, 649–670. [Google Scholar] [CrossRef] [PubMed]
- Tierney, K.B.; Baldwin, D.H.; Hara, T.J.; Ross, P.S.; Scholz, N.L.; Kennedy, C.J. Olfactory toxicity in fishes. Aquat. Toxicol. 2010, 96, 2–26. [Google Scholar] [CrossRef] [PubMed]
- Cancalon, P. Degeneration and regeneration of olfactory cells induced by ZnSO4 and other chemicals. Tissue Cell 1982, 14, 717–733. [Google Scholar] [CrossRef] [PubMed]
- Becker, C.G.; Becker, T. Adult zebrafish as a model for successful central nervous system regeneration. Restor. Neurol. Neurosci. 2008, 26, 71–80. [Google Scholar] [PubMed]
- Hill, A.J.; Teraoka, H.; Heideman, W.; Peterson, R.E. Zebrafish as a model vertebrate for investigating chemical toxicity. Toxicol. Sci. 2005, 86, 6–19. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-C.; Lu, P.-N.; Huang, H.-L.; Chu, C.; Li, H.-P.; Tsai, H.-J. Zebrafish transgenic line huORFZ is an effective living bioindicator for detecting environmental toxicants. PLoS ONE 2014, 9, e90160. [Google Scholar] [CrossRef] [PubMed]
- Bettini, S.; Lazzari, M.; Ferrando, S.; Gallus, L.; Franceschini, V. Histopathological analysis of the olfactory epithelium of zebrafish (Danio rerio) exposed to sublethal doses of urea. J. Anat. 2016, 228, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Byrd, C.A.; Brunjes, P.C. Organization of the olfactory systems in the adult zebrafish: Histological, immunohistochemical, and quantitative analysis. J. Comp. Neurol. 1995, 385, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Hansen, A.; Zeiske, E. The peripheral olfactory organ of the zebrafish, Danio rerio: An ultrastructural study. Chem. Senses 1998, 23, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Hansen, A.; Zielinski, B. Diversity in the olfactory epithelium of bony fishes: Development, lamellar arrangement, sensory neuron cell types and transduction components. J. Neurocytol. 2005, 34, 183–208. [Google Scholar] [CrossRef] [PubMed]
- Koide, T.; Miyasaka, N.; Morimoto, K.; Asakawa, K.; Urasaki, A.; Kawakami, K.; Yoshihara, Y. Olfactory neural circuitry for attraction to amino acids revealed by transposon-mediated gene trap approach in zebrafish. Proc. Natl. Acad. Sci. USA 2009, 106, 9884–9889. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Mack, J.A.; Souren, M.; Yaksi, E.; Higashijima, S.; Mione, M.; Fetcho, J.R.; Friedrich, R.W. Early development of functional spatial maps in the zebrafish olfactory bulb. J. Neurosci. 2005, 25, 5784–5795. [Google Scholar] [CrossRef] [PubMed]
- Yaksi, E.; Judkewitz, B.; Friedrich, R.W. Topological reorganization of odor representations in the olfactory bulb. PLoS Biol. 2007, 5, e178. [Google Scholar] [CrossRef] [PubMed]
- Lipschitz, D.; Michel, W. Amino acid odorants stimulate microvillar sensory neurons. Chem. Senses 2002, 27, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Hamdani, E.; Doving, K.B. Specific projections of the sensory crypt cells in the olfactory system in crucian carp, Carassius carassius. Chem. Senses 2006, 31, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Hamdani, E.H.; Lastein, S.; Gregersen, F.; Doving, K.B. Seasonal variations in olfactory sensory neurons—Fish sensitivity to sex pheromones explained? Chem. Senses 2008, 33, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, T.; Byrd-Jacobs, C. Rapid degeneration and regeneration of the zebrafish olfactory epithelium after Triton X-100 application. Chem. Senses 2010, 35, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Paskin, T.R.; Byrd-Jacobs, C.A. Reversible deafferentation of the adult zebrafish olfactory bulb affects glomerular distribution and olfactory-mediated behavior. Behav. Brain Res. 2012, 235, 293–301. [Google Scholar] [CrossRef] [PubMed]
- White, E.J.; Kounelis, S.K.; Byrd-Jacobs, C.A. Plasticity of glomeruli and olfactory-mediated behavior in zebrafish following detergent lesioning of the olfactory epithelium. Neuroscience 2015, 284, 622–631. [Google Scholar] [CrossRef] [PubMed]
- Castro, A.; Becerra, M.; Manso, M.J.; Anadon, R. Calretinin immunoreactivity in the brain of the zebrafish, Danio rerio: Distribution and comparison with some neuropeptides and neurotransmitter-synthesizing enzymes. I. Olfactory organ and forebrain. J. Comp. Neurol. 2006, 494, 435–459. [Google Scholar] [CrossRef] [PubMed]
- Gayoso, J.A.; Castro, A.; Anadon, R.; Manso, M.J. Differential bulbar and extrabulbar projections of diverse olfactory receptor neuron populations in the adult zebrafish (Danio rerio). J. Comp. Neurol. 2011, 519, 247–276. [Google Scholar] [CrossRef] [PubMed]
- Paskin, T.R.; Iqbal, T.R.; Byrd-Jacobs, C.A. Olfactory bulb recovery following reversible deafferentation with repeated detergent application in the adult zebrafish. Neuroscience 2011, 196, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Braubach, O.R.; Fine, A.; Croll, R.P. Distribution and functional organization of glomeruli in the olfactory bulbs of zebrafish (Danio rerio). J. Comp. Neurol. 2012, 520, 2317–2339. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, D.; Mandal, D.K. Mercuric chloride induced toxicity responses in the olfactory epithelium of Labeo rohita (Hamilton): A light and electron microscopy study. Fish Physiol. Biochem. 2014, 40, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Kalueff, A.V.; Gebhardt, M.; Stewart, A.M.; Cachat, J.M.; Brimmer, M.; Chawla, J.S.; Craddock, C.; Kyzar, E.J.; Roth, A.; Landsman, S.; et al. Towards a comprehensive catalog of zebrafish behavior 1.0 and beyond. Zebrafish 2013, 10, 70–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultz, E.W.; Gebhardt, L.P. The use of zinc sulfate solution for the prevention of poliomyelitis in man. JAMA 1938, 110, 2024. [Google Scholar] [CrossRef]
- Tisdall, F.F.; Brown, A.; Defries, R.D. Persistent anosmia following zinc sulfate nasal spraying. J. Pediatr. 1938, 13, 60–62. [Google Scholar] [CrossRef]
- Jafek, B.W.; Linschoten, M.R.; Murrow, B.W. Anosmia after intranasal zinc gluconate use. Am. J. Rhinol. 2004, 18, 137–141. [Google Scholar] [PubMed]
- Lim, J.H.; Davis, G.E.; Wang, Z.; Li, V.; Wu, Y.; Rue, T.C.; Storm, D.R. Zicam-induced damage to mouse and human nasal tissue. PLoS ONE 2009, 4, e7647. [Google Scholar] [CrossRef] [PubMed]
- Matulionis, D.H. Ultrastructural study of mouse olfactory epithelium following destruction by ZnSO4 and its subsequent regeneration. Am. J. Anat. 1975, 142, 67–90. [Google Scholar] [CrossRef] [PubMed]
- Burd, G.D. Morphological study of the effects of intranasal zinc sulfate irrigation on the mouse olfactory epithelium and olfactory bulb. Microsc. Res. Tech. 1993, 24, 195–213. [Google Scholar] [CrossRef] [PubMed]
- Slotnick, B.; Glover, P.; Bodyak, N. Does intranasal application of zinc sulfate produce anosmia in the rat? Behav. Neurosci. 2000, 114, 814–829. [Google Scholar] [CrossRef] [PubMed]
- McBride, K.; Slotnick, B.; Margolis, F. Does intranasal application of zinc sulfate produce anosmia in the mouse? Chem. Senses 2003, 28, 659–670. [Google Scholar] [CrossRef] [PubMed]
- Yovanovich, C.A.M.; Jungblut, L.D.; Heer, T.; Pozzi, A.G.; Paz, D.A. Amphibian larvae and zinc sulphate: A suitable model to study the role of brain-derived neurotrophic factor (BDNF) in the neuronal turnover of the olfactory epithelium. Cell Tissue Res. 2009, 336, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Frontera, J.L.; Raices, M.; Cervino, A.S.; Pozzi, A.G.; Paz, D.A. Neural regeneration dynamics of Xenopus laevis olfactory epithelium after zinc sulfate-induced damage. J. Chem. Neuroanat. 2016, 77, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Saucier, D.; Astic, L. Morpho-functional alterations in the olfactory system of rainbow trout (Oncorhynchus mykiss) and possible acclimation in response to long-lasting exposure to low levels of copper. Comp. Biochem. Physiol. A Physiol. 1995, 112, 273–284. [Google Scholar] [CrossRef]
- Roy, D.; Ghosh, D.; Mandal, D.K. Cadmium induced histopathology in the olfactory epithelium of a snakehead fish, Channa punctatus (Bloch). Int. J. Aquat. Biol. 2013, 1, 221–227. [Google Scholar]
- Cancalon, P. Regeneration of three populations of olfactory axons as a function of temperature. Dev. Brain Res. 1983, 9, 265–278. [Google Scholar] [CrossRef]
- Zielinski, B.; Hara, T.J. Ciliated and microvillar receptor cells degenerate and then differentiate in the olfactory epithelium of rainbow trout following olfactory nerve section. Microsc. Res. Tech. 1992, 23, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Graziadei, P.P.C.; Monti-Graziadei, G.A. Neurogenesis and neuron regeneration in the olfactory system of mammals. I. Morphological aspects of differentiation and structural organization of the olfactory sensory neurons. J. Neurocytol. 1979, 8, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Starcevic, S.L.; Muruganandam, A.; Mutus, B.; Zielinski, B.S. Glutathione in the olfactory mucosa of rainbow trout (Oncorhyncus mykiss). Chem. Senses 1993, 18, 57–65. [Google Scholar] [CrossRef]
- Starcevic, S.L.; Zielinski, B.S. Immunohistochemical localization of glutathione S-transferase pi in rainbow trout olfactory receptor neurons. Neurosci. Lett. 1995, 183, 175–178. [Google Scholar] [CrossRef]
- Chen, W.-Y.; John, J.A.C.; Lin, C.-H.; Chang, C.-Y. Expression pattern of metallothionein, MTF-1 nuclear translocation, and its DNA-binding activity in zebrafish (Danio rerio) induced by zinc and cadmium. Environ. Toxicol. Chem. 2007, 26, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Carr, V.M.; Ring, G.; Youngentob, S.L.; Schwob, J.E.; Farbman, A.I. Altered epithelial density and expansion of bulbar projections of a discrete HSP70 immunoreactive subpopulation of rat olfactory receptor neurons in reconstituting olfactory epithelium following exposure to methyl bromide. J. Comp. Neurol. 2004, 469, 475–493. [Google Scholar] [CrossRef] [PubMed]
- Hegg, C.C.; Lucero, M.T. Purinergic receptor antagonists inhibit odorant-induced heat shock protein 25 induction in mouse olfactory epithelium. Glia 2006, 53, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Matz, C.J.; Krone, P.H. Cell death, stress-responsive transgene activation, and deficits in the olfactory system of larval zebrafish following cadmium exposure. Environ. Sci. Technol. 2007, 41, 5143–5148. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Gallagher, E.P. Role of Nrf2 antioxidant defense in mitigating cadmium-induced oxidative stress in the olfactory system of zebrafish. Toxicol. Appl. Pharmacol. 2013, 266, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Vinnikov, Y.A. Glycocalyx of receptor cell membranes. Chem. Senses 1986, 11, 243–257. [Google Scholar] [CrossRef]
- Foster, J.D.; Getchell, M.L.; Getchell, T.V. Ultrastructural localization of sialylated glycoconjugates in cells of the salamander olfactory mucosa using lectin cytochemistry. Cell Tissue Res. 1992, 267, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Youngentob, S.L.; Schwob, J.E. Changes in odorant quality perception following methyl bromide induced lesions of the olfactory epithelium. Chem. Senses 1997, 22, 830–831. [Google Scholar]
- Kolmakov, N.N.; Hubbard, P.C.; Lopes, O.; Canario, A.V.M. Effect of acute copper sulfate exposure on olfactory responses to amino acids and pheromones in goldfish (Carassius auratus). Environ. Sci. Technol. 2009, 43, 8393–8399. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.R.; Gallagher, E.P. Effects of cadmium on olfactory mediated behaviors and molecular biomarkers in coho salmon (Oncorhynchus kisutch). Aquat. Toxicol. 2013, 140, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, D.H.; Sandahl, J.F.; Labenia, J.S.; Scholz, N.L. Sublethal effects of copper on coho salmon: Impacts on nonoverlapping receptor pathways in the peripheral olfactory nervous system. Environ. Toxicol. Chem. 2003, 22, 2266–2274. [Google Scholar] [CrossRef] [PubMed]





© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hentig, J.T.; Byrd-Jacobs, C.A. Exposure to Zinc Sulfate Results in Differential Effects on Olfactory Sensory Neuron Subtypes in Adult Zebrafish. Int. J. Mol. Sci. 2016, 17, 1445. https://doi.org/10.3390/ijms17091445
Hentig JT, Byrd-Jacobs CA. Exposure to Zinc Sulfate Results in Differential Effects on Olfactory Sensory Neuron Subtypes in Adult Zebrafish. International Journal of Molecular Sciences. 2016; 17(9):1445. https://doi.org/10.3390/ijms17091445
Chicago/Turabian StyleHentig, James T., and Christine A. Byrd-Jacobs. 2016. "Exposure to Zinc Sulfate Results in Differential Effects on Olfactory Sensory Neuron Subtypes in Adult Zebrafish" International Journal of Molecular Sciences 17, no. 9: 1445. https://doi.org/10.3390/ijms17091445
APA StyleHentig, J. T., & Byrd-Jacobs, C. A. (2016). Exposure to Zinc Sulfate Results in Differential Effects on Olfactory Sensory Neuron Subtypes in Adult Zebrafish. International Journal of Molecular Sciences, 17(9), 1445. https://doi.org/10.3390/ijms17091445