The Small Heat Shock Protein α-Crystallin B Shows Neuroprotective Properties in a Glaucoma Animal Model

Abstract
:
{kind=link}
{kind=link}
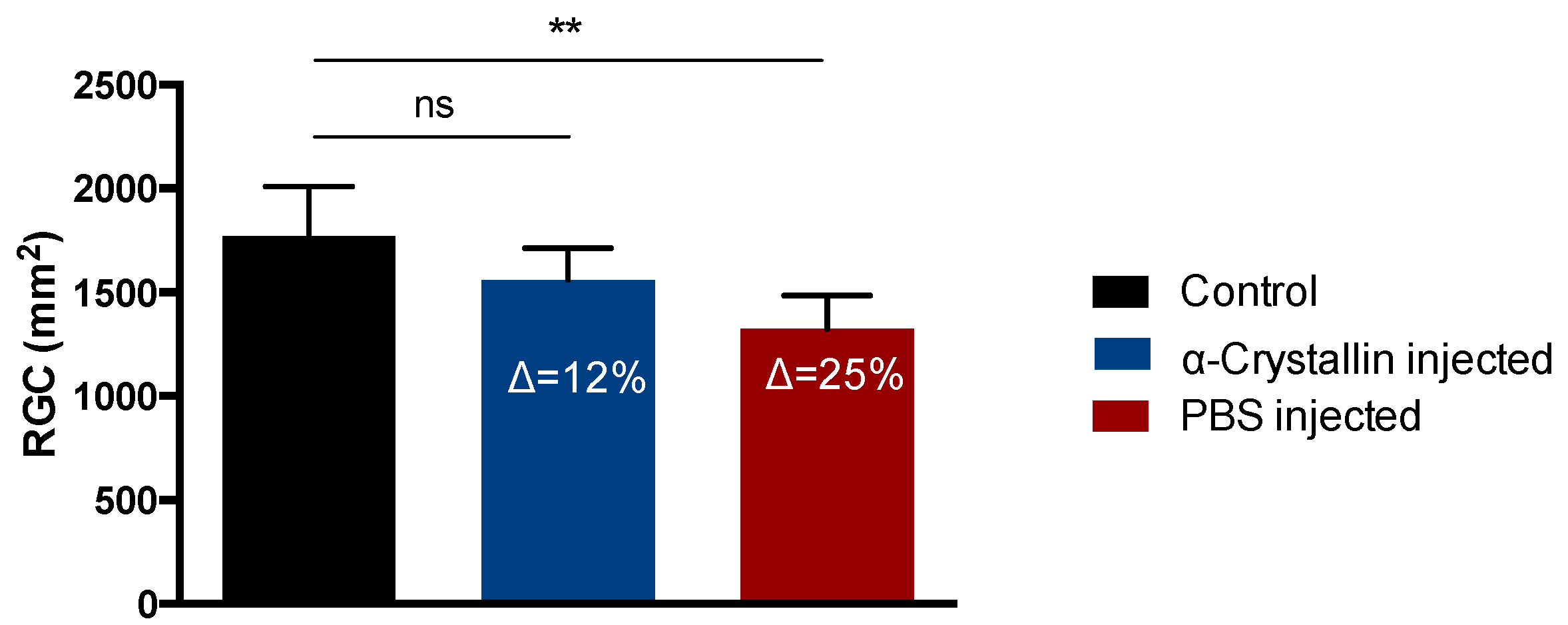{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. IOP Elevation
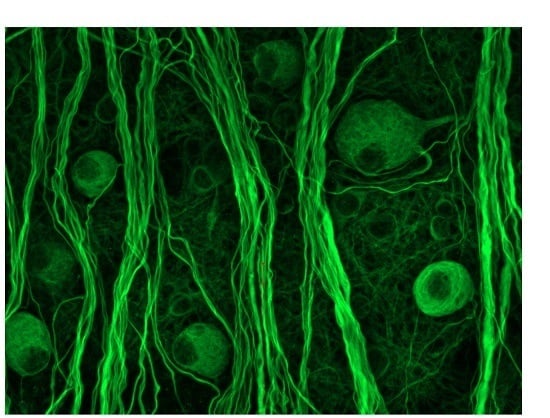
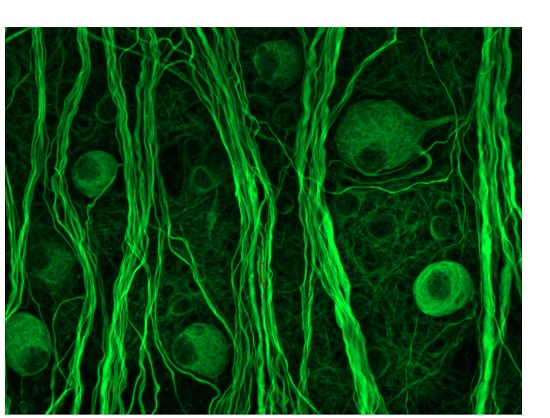
2.2. Morphological Changes
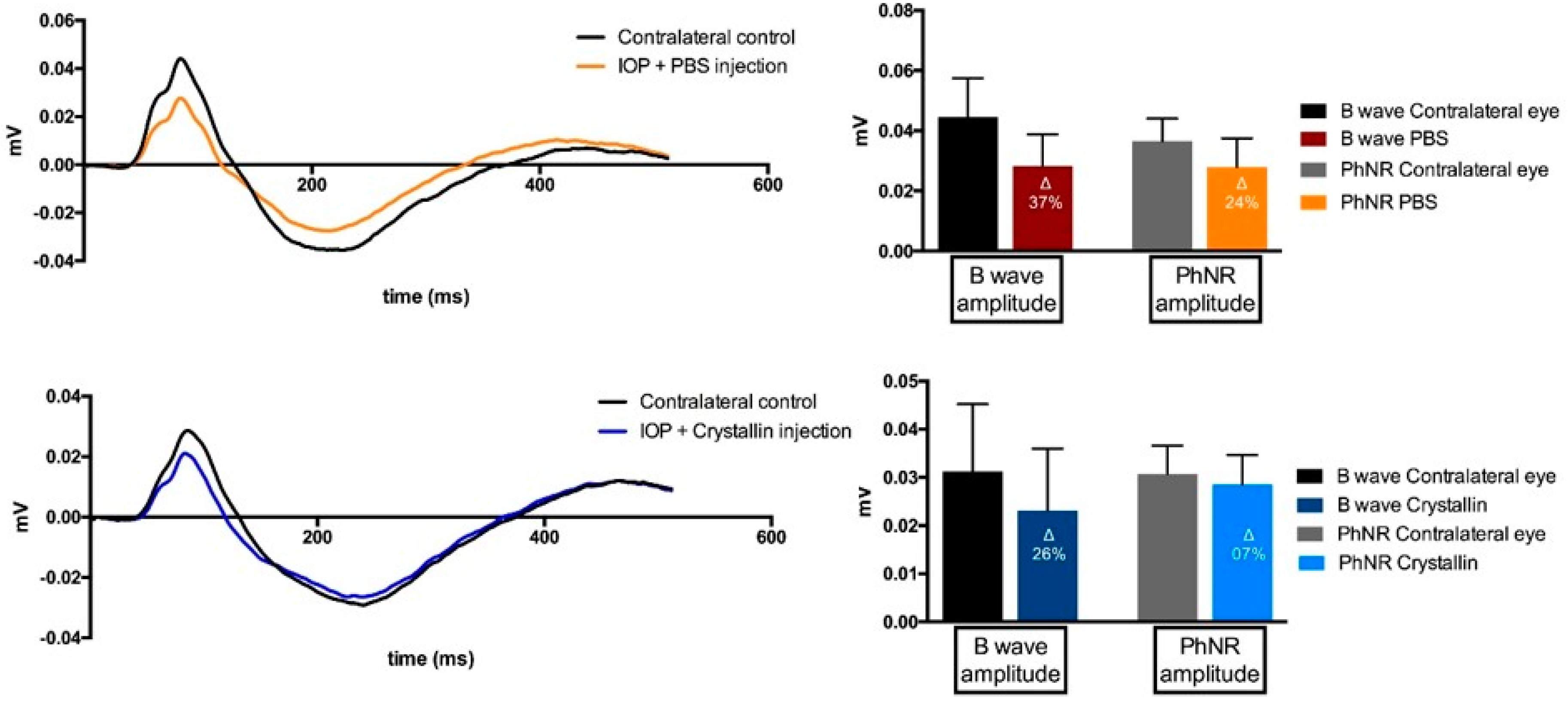
2.3. Functional Changes
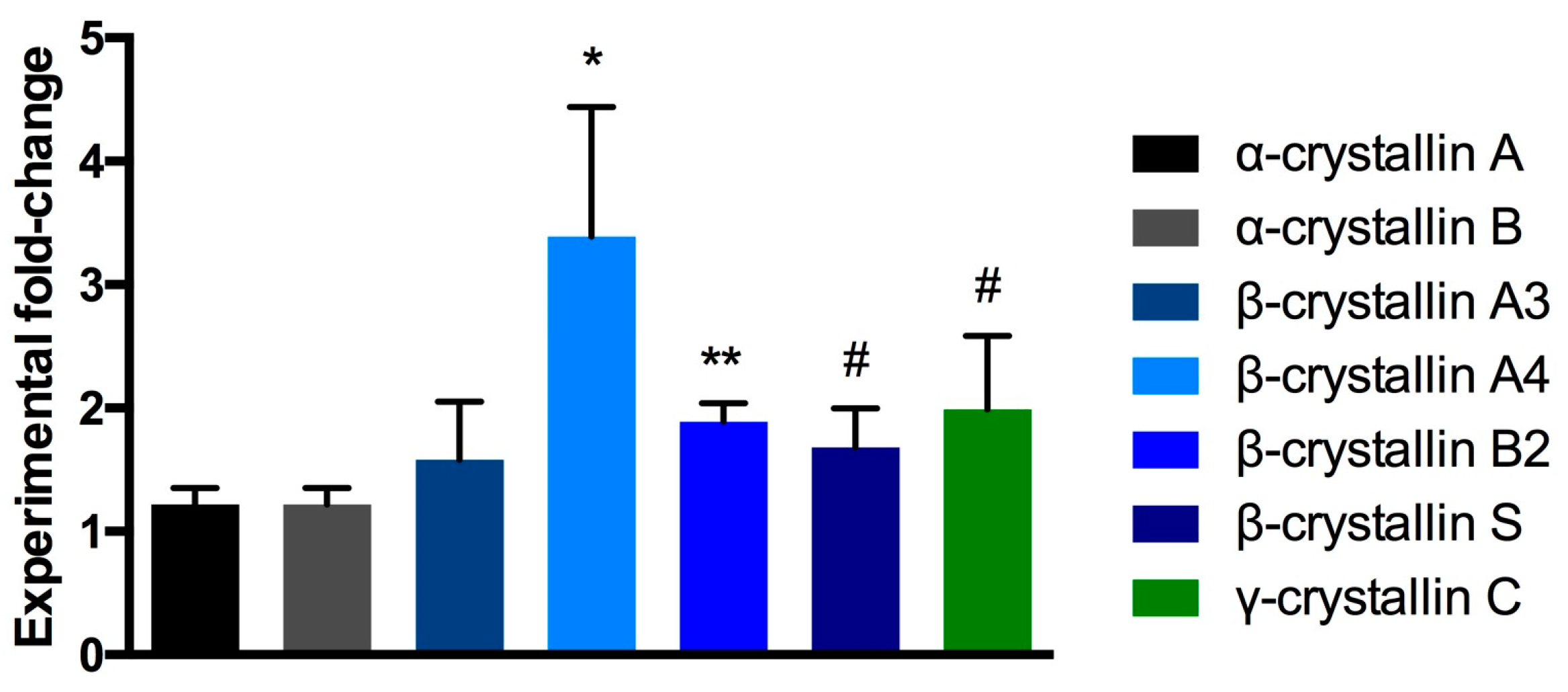
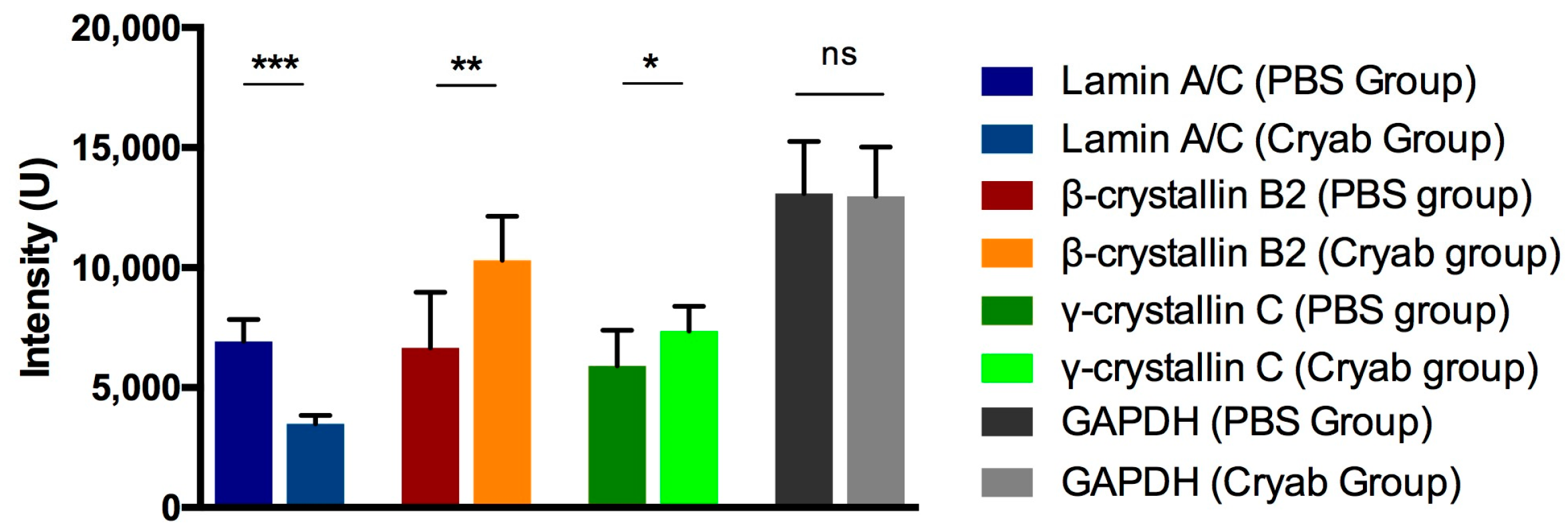
2.4. Proteomic Analysis
3. Discussion
4. Materials and Methods
4.1. Ethical Statement and Animal Experiments
4.2. Episcleral Vein Cauterization
4.3. Intravitreal Injection of α-Crystallin B
4.4. Quantification of Retinal Ganglion Cell Density
4.5. Optical Coherence Tomography
4.6. Ganzfeld Photopic Electroretinogram
4.7. Mass Spectrometry Sample Preparation
4.8. MS Measurement and Data Analysis
4.9. Antibody Microarray
4.10. Bioinformatics Analysis and Proteomic Networks
4.11. Experimental Design and Statistics
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ACN | Acetonitrile |
| AMD | Age-related macular degeneration |
| ARVO | Association of Research in Vision and Ophthalmology |
| BRN3A | Brain-specific homeobox/POU domain protein 3A |
| CNTF | Ciliary neurotrophic factor |
| Cryab | α-crystallin B |
| ERG | Electroretinogram |
| FDR | False discovery rate |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| IL-6 | Interleukin 6 |
| IOP | Intraocular pressure |
| IV | intravitreal |
| LFQ | Label-free quantification |
| MS | Mass-spectrometry |
| PBS | phosphate-buffered saline |
| PhNR | Photopic negative response |
| RGC | Retinal ganglion cell |
| RNFL | Retinal nerve fiber layer |
| RNFLT | Retinal nerve fiber layer thickness |
| sHSP | Small heat shock protein |
| TARC | Translational animal research center |
| uHPLC | Ultra-high pressure liquid chromatography |
| UVA | Ultraviolet A radiation |
| VEGF | Vascular endothelial growth factor |
References
- Weinreb, R.N.; Aung, T.; Medeiros, F.A. The pathophysiology and treatment of glaucoma: A review. JAMA 2014, 311, 1901–1911. [Google Scholar] [CrossRef] [PubMed]
- Jonas, J.B.; Aung, T.; Bourne, R.R.; Bron, A.M.; Ritch, R.; Panda-Jonas, S. Glaucoma. Lancet 2017, 390, 2183–2193. [Google Scholar] [CrossRef]
- Quigley, H.A. Number of people with glaucoma worldwide. Br. J. Ophthalmol. 1996, 80, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Quigley, H.A.; Broman, A.T. The number of people with glaucoma worldwide in 2010 and 2020. Br. J. Ophthalmol. 2006, 90, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Susanna, R., Jr.; De Moraes, C.G.; Cioffi, G.A.; Ritch, R. Why Do People (Still) Go Blind from Glaucoma? Transl. Vis. Sci. Technol. 2015, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.R. Normal-tension glaucoma (low-tension glaucoma). Indian J. Ophthalmol. 2011, 59 (Suppl. S1), 97–101. [Google Scholar] [CrossRef] [PubMed]
- Choplin, N.T. Correlation of asymmetric damage with asymmetric intraocular pressure in normal-tension glaucoma (low-tension glaucoma). Arch. Ophthalmol. 1989, 107, 167–168. [Google Scholar] [CrossRef] [PubMed]
- Abe, R.Y.; Gracitelli, C.P.; Diniz-Filho, A.; Tatham, A.J.; Medeiros, F.A. Lamina Cribrosa in Glaucoma: Diagnosis and Monitoring. Curr. Ophthalmol. Rep. 2015, 3, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; McNally, S.; Kilpatrick, J.I.; Jarvis, S.P.; O’Brien, C.J. Ageing and Ocular Tissue Stiffness in Glaucoma. Surv. Ophthalmol. 2017, in press. [Google Scholar]
- Hall, J.K.; Andrews, A.P.; Walker, R.; Piltz-Seymour, J.R. Association of retinal vessel caliber and visual field defects in glaucoma. Am. J. Ophthalmol. 2001, 132, 855–859. [Google Scholar] [CrossRef]
- Izzotti, A.; Bagnis, A.; Sacca, S.C. The role of oxidative stress in glaucoma. Mutat. Res. 2006, 612, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, O.; Ates, N.A.; Ercan, B.; Muslu, N.; Unlu, A.; Tamer, L.; Atik, U.; Kanık, A. Role of oxidative stress enzymes in open-angle glaucoma. Eye 2005, 19, 580–583. [Google Scholar] [CrossRef] [PubMed]
- Dreyer, E.B.; Zurakowski, D.; Schumer, R.A.; Podos, S.M.; Lipton, S.A. Elevated glutamate levels in the vitreous body of humans and monkeys with glaucoma. Arch. Ophthalmol. 1996, 114, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Cavet, M.E.; Vittitow, J.L.; Impagnatiello, F.; Ongini, E.; Bastia, E. Nitric oxide (NO): An emerging target for the treatment of glaucoma. Investig. Ophthalmol. Vis. Sci. 2014, 55, 5005–5015. [Google Scholar] [CrossRef] [PubMed]
- Anders, F.; Teister, J.; Liu, A.; Funke, S.; Grus, F.H.; Thanos, S.; von Pein, H.D.; Pfeiffer, N.; Prokosch, V. Intravitreal injection of β-crystallin B2 improves retinal ganglion cell survival in an experimental animal model of glaucoma. PLoS ONE 2017, 12, e0175451. [Google Scholar] [CrossRef] [PubMed]
- Piri, N.; Song, M.; Kwong, J.M.; Caprioli, J. Modulation of α and β crystallin expression in rat retinas with ocular hypertension-induced ganglion cell degeneration. Brain Res. 2007, 1141, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhou, Q.; Tang, M.; Fu, N.; Shao, W.; Zhang, S.; Yin, Y.; Zeng, R.; Wang, X.; Hu, G.; et al. Upregulation of αB-crystallin expression in the substantia nigra of patients with Parkinson’s disease. Neurobiol. Aging 2015, 36, 1686–1691. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, S.; Kamps, B.; Boelens, W.C.; Reif, B. αB-crystallin competes with Alzheimer’s disease β-amyloid peptide for peptide-peptide interactions and induces oxidation of Abeta-Met35. FEBS Lett. 2006, 580, 5941–5946. [Google Scholar] [CrossRef] [PubMed]
- Heise, E.A.; Fort, P.E. Impact of diabetes on α-crystallins and other heat shock proteins in the eye. J. Ocul. Biol. Dis. Infor. 2011, 4, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Kannan, R.; Sreekumar, P.G.; Hinton, D.R. α Crystallins in the retinal pigment epithelium and implications for the pathogenesis and treatment of age-related macular degeneration. Biochim. Biophys. Acta 2016, 1860, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Derham, B.K.; Harding, J.J. α-Crystallin as a molecular chaperone. Prog. Retin. Eye Res. 1999, 18, 463–509. [Google Scholar] [CrossRef]
- Garcia-Valenzuela, E.; Shareef, S.; Walsh, J.; Sharma, S.C. Programmed cell death of retinal ganglion cells during experimental glaucoma. Exp. Eye Res. 1995, 61, 33–44. [Google Scholar] [CrossRef]
- Wilsey, L.; Gowrisankaran, S.; Cull, G.; Hardin, C.; Burgoyne, C.F.; Fortune, B. Comparing three different modes of electroretinography in experimental glaucoma: Diagnostic performance and correlation to structure. Doc. Ophthalmol. Adv. Ophthalmol. 2017, 134, 111–128. [Google Scholar] [CrossRef] [PubMed]
- Bayer, A.U.; Neuhardt, T.; May, A.C.; Martus, P.; Maag, K.P.; Brodie, S.; Lütjen-Drecoll, E.; Podos, S.M.; Mittag, T. Retinal morphology and ERG response in the DBA/2NNia mouse model of angle-closure glaucoma. Investig. Ophthalmol. Vis. Sci. 2001, 42, 1258–1265. [Google Scholar]
- Bayer, A.U.; Danias, J.; Brodie, S.; Maag, K.P.; Chen, B.; Shen, F.; Podos, S.M.; Mittag, T.W. Electroretinographic abnormalities in a rat glaucoma model with chronic elevated intraocular pressure. Exp. Eye Res. 2001, 72, 667–677. [Google Scholar] [CrossRef] [PubMed]
- Nork, T.M.; Ver Hoeve, J.N.; Poulsen, G.L.; Nickells, R.W.; Davis, M.D.; Weber, A.J.; Sarks, S.H.; Lemley, H.L.; Millecchia, L.L. Swelling and loss of photoreceptors in chronic human and experimental glaucomas. Arch. Ophthalmol. 2000, 118, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, M.; Rodriguez, F.D.; Sharma, S.C.; Vecino, E. Immunohistochemical changes in rat retinas at various time periods of elevated intraocular pressure. Mol. Vis. 2009, 15, 2696–2709. [Google Scholar] [PubMed]
- Liu, J.P.; Schlosser, R.; Ma, W.Y.; Dong, Z.; Feng, H.; Lui, L.; Huang, X.Q.; Li, D.W. Human αA- and αB-crystallins prevent UVA-induced apoptosis through regulation of PKCα, RAF/MEK/ERK and AKT signaling pathways. Exp. Eye Res. 2004, 79, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Losiewicz, M.K.; Fort, P.E. Diabetes impairs the neuroprotective properties of retinal α-crystallins. Investig. Ophthalmol. Vis. Sci. 2011, 52, 5034–5042. [Google Scholar] [CrossRef] [PubMed]
- Masilamoni, J.G.; Vignesh, S.; Kirubagaran, R.; Jesudason, E.P.; Jayakumar, R. The neuroprotective efficacy of α-crystallin against acute inflammation in mice. Brain Res. Bull. 2005, 67, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Ousman, S.S.; Tomooka, B.H.; van Noort, J.M.; Wawrousek, E.F.; O’Connor, K.C.; Hafler, D.A.; Sobel, R.A.; Robinson, W.H.; Steinman, L. Protective and therapeutic role for αB-crystallin in autoimmune demyelination. Nature 2007, 448, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Doss, E.W.; Ward, K.A.; Koretz, J.F. Investigation of the ‘fines’ hypothesis of primary open-angle glaucoma: The possible role of α-crystallin. Ophthalmic Res. 1998, 30, 142–156. [Google Scholar] [CrossRef] [PubMed]
- Chiu, K.; Zhou, Y.; Yeung, S.C.; Lok, C.K.; Chan, O.O.; Chang, R.C.; So, K.F.; Chiu, J.F. Up-regulation of crystallins is involved in the neuroprotective effect of wolfberry on survival of retinal ganglion cells in rat ocular hypertension model. J. Cell. Biochem. 2010, 110, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Thanos, S.; Bohm, M.R.; Zu Hörste, M.M.; Prokosch-Willing, V.; Hennig, M.; Bauer, D.; Heiligenhaus, A. Role of crystallins in ocular neuroprotection and axonal regeneration. Prog. Retin. Eye Res. 2014, 42, 145–161. [Google Scholar] [CrossRef] [PubMed]
- Bohm, M.R.; Prokosch, V.; Bruckner, M.; Pfrommer, S.; Melkonyan, H.; Thanos, S. βB2-Crystallin Promotes Axonal Regeneration in the Injured Optic Nerve in Adult Rats. Cell Transplant. 2015, 24, 1829–1844. [Google Scholar] [CrossRef] [PubMed]
- Bohm, M.R.; Pfrommer, S.; Chiwitt, C.; Bruckner, M.; Melkonyan, H.; Thanos, S. Crystallin-β-b2-overexpressing NPCs support the survival of injured retinal ganglion cells and photoreceptors in rats. Investig. Ophthalmol. Vis. Sci. 2012, 53, 8265–8279. [Google Scholar] [CrossRef] [PubMed]
- Fischer, D.; Hauk, T.G.; Muller, A.; Thanos, S. Crystallins of the β/γ-superfamily mimic the effects of lens injury and promote axon regeneration. Mol. Cell. Neurosci. 2008, 37, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Muller, A.; Hauk, T.G.; Leibinger, M.; Marienfeld, R.; Fischer, D. Exogenous CNTF stimulates axon regeneration of retinal ganglion cells partially via endogenous CNTF. Mol. Cell. Neurosci. 2009, 41, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Pucci, S.; Mazzarelli, P.; Missiroli, F.; Regine, F.; Ricci, F. Neuroprotection: VEGF, IL-6, and clusterin: The dark side of the moon. Prog. Brain Res. 2008, 173, 555–573. [Google Scholar] [PubMed]
- Samejima, K.; Svingen, P.A.; Basi, G.S.; Kottke, T.; Mesner, P.W., Jr.; Stewart, L.; Durrieu, F.; Poirier, G.G.; Alnemri, E.S.; Champoux, J.J.; et al. Caspase-mediated cleavage of DNA topoisomerase I at unconventional sites during apoptosis. J. Biol. Chem. 1999, 274, 4335–4340. [Google Scholar] [CrossRef] [PubMed]
- Okinaga, T.; Kasai, H.; Tsujisawa, T.; Nishihara, T. Role of caspases in cleavage of lamin A/C and PARP during apoptosis in macrophages infected with a periodontopathic bacterium. J. Med. Microbiol. 2007, 56, 1399–1404. [Google Scholar] [CrossRef] [PubMed]
- Anders, F.; Teister, J.; Funke, S.; Pfeiffer, N.; Grus, F.; Solon, T.; Prokosch, V. Proteomic profiling reveals crucial retinal protein alterations in the early phase of an experimental glaucoma model. Graefe’s Arch. Clin. Exp. Ophthalmol. 2017, 255, 1395–1407. [Google Scholar] [CrossRef] [PubMed]
- Burke, B. Lamins and apoptosis: A two-way street? J. Cell Biol. 2001, 153, F5–F7. [Google Scholar] [CrossRef] [PubMed]
- Broers, J.L.; Bronnenberg, N.M.; Kuijpers, H.J.; Schutte, B.; Hutchison, C.J.; Ramaekers, F.C. Partial cleavage of A-type lamins concurs with their total disintegration from the nuclear lamina during apoptosis. Eur. J. Cell Biol. 2002, 81, 677–691. [Google Scholar] [CrossRef] [PubMed]
- Cross, T.; Griffiths, G.; Deacon, E.; Sallis, R.; Gough, M.; Watters, D.; Lord, J.M. PKC-delta is an apoptotic lamin kinase. Oncogene 2000, 19, 2331–2337. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, A.S.; Sridhar Rao, K.; Rangaraj, N.; Parnaik, V.K.; Mohan Rao, C. Heat stress-induced localization of small heat shock proteins in mouse myoblasts: Intranuclear lamin A/C speckles as target for αB-crystallin and Hsp25. Exp. Cell Res. 2004, 299, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, D.; Dvoriantchikova, G.; Nathanson, L.; McKinnon, S.J.; Shestopalov, V.I. Microarray analysis of gene expression in adult retinal ganglion cells. FEBS Lett. 2006, 580, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Springelkamp, H.; Iglesias, A.I.; Cuellar-Partida, G.; Amin, N.; Burdon, K.P.; van Leeuwen, E.M.; Gharahkhani, P.; Mishra, A.; van der Lee, S.J.; Hewitt, A.W.; et al. ARHGEF12 influences the risk of glaucoma by increasing intraocular pressure. Hum. Mol. Genet. 2015, 24, 2689–2699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, U.R.; Madden, B.J.; Charlesworth, M.C.; Fautsch, M.P. Proteome analysis of human aqueous humor. Investig. Ophthalmol. Vis. Sci. 2010, 51, 4921–4931. [Google Scholar] [CrossRef] [PubMed]
- Bonnemann, C.G.; Thompson, T.G.; van der Ven, P.F.; Goebel, H.H.; Warlo, I.; Vollmers, B.; Reimann, J.; Herms, J.; Gautel, M.; Takada, F.; et al. Filamin C accumulation is a strong but nonspecific immunohistochemical marker of core formation in muscle. J. Neurol. Sci. 2003, 206, 71–78. [Google Scholar] [CrossRef]
- Hagemann, T.L.; Boelens, W.C.; Wawrousek, E.F.; Messing, A. Suppression of GFAP toxicity by αB-crystallin in mouse models of Alexander disease. Hum. Mol. Genet. 2009, 18, 1190–1199. [Google Scholar] [CrossRef] [PubMed]
- Prokai-Tatrai, K.; Xin, H.; Nguyen, V.; Szarka, S.; Blazics, B.; Prokai, L.; Koulen, P. 17β-estradiol eye drops protect the retinal ganglion cell layer and preserve visual function in an in vivo model of glaucoma. Mol. Pharm. 2013, 10, 3253–3261. [Google Scholar] [CrossRef] [PubMed]
- Funke, S.; Perumal, N.; Beck, S.; Gabel-Scheurich, S.; Schmelter, C.; Teister, J.; Gerbig, C.; Gramlich, O.W.; Pfeiffer, N.; Grus, F.H. Glaucoma related Proteomic Alterations in Human Retina Samples. Sci. Rep. 2016, 6, 29759. [Google Scholar] [CrossRef] [PubMed]
- Shareef, S.R.; Garcia-Valenzuela, E.; Salierno, A.; Walsh, J.; Sharma, S.C. Chronic ocular hypertension following episcleral venous occlusion in rats. Exp. Eye Res. 1995, 61, 379–382. [Google Scholar] [CrossRef]
- Nadal-Nicolas, F.M.; Jimenez-Lopez, M.; Sobrado-Calvo, P.; Nieto-Lopez, L.; Canovas-Martinez, I.; Salinas-Navarro, M.; Vidal-Sanz, M.; Agudo, M. Brn3a as a marker of retinal ganglion cells: Qualitative and quantitative time course studies in naive and optic nerve-injured retinas. Investig. Ophthalmol. Vis. Sci. 2009, 50, 3860–3868. [Google Scholar] [CrossRef] [PubMed]
- Rappsilber, J.; Mann, M.; Ishihama, Y. Protocol for micro-purification, enrichment, pre-fractionation and storage of peptides for proteomics using StageTips. Nat. Protoc. 2007, 2, 1896–1906. [Google Scholar] [CrossRef] [PubMed]
- Bluhm, A.; Casas-Vila, N.; Scheibe, M.; Butter, F. Reader interactome of epigenetic histone marks in birds. Proteomics 2016, 16, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Vizcaino, J.A.; Csordas, A.; Del-Toro, N.; Dianes, J.A.; Griss, J.; Lavidas, I.; Mayer, G.; Perez-Riverol, Y.; Reisinger, F.; Ternent, T.; et al. 2016 update of the PRIDE database and its related tools. Nucleic Acids Res. 2016, 44, D447–D456. [Google Scholar] [CrossRef] [PubMed]







© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anders, F.; Liu, A.; Mann, C.; Teister, J.; Lauzi, J.; Thanos, S.; Grus, F.H.; Pfeiffer, N.; Prokosch, V. The Small Heat Shock Protein α-Crystallin B Shows Neuroprotective Properties in a Glaucoma Animal Model. Int. J. Mol. Sci. 2017, 18, 2418. https://doi.org/10.3390/ijms18112418
Anders F, Liu A, Mann C, Teister J, Lauzi J, Thanos S, Grus FH, Pfeiffer N, Prokosch V. The Small Heat Shock Protein α-Crystallin B Shows Neuroprotective Properties in a Glaucoma Animal Model. International Journal of Molecular Sciences. 2017; 18(11):2418. https://doi.org/10.3390/ijms18112418
Chicago/Turabian StyleAnders, Fabian, Aiwei Liu, Carolina Mann, Julia Teister, Jasmin Lauzi, Solon Thanos, Franz H. Grus, Norbert Pfeiffer, and Verena Prokosch. 2017. "The Small Heat Shock Protein α-Crystallin B Shows Neuroprotective Properties in a Glaucoma Animal Model" International Journal of Molecular Sciences 18, no. 11: 2418. https://doi.org/10.3390/ijms18112418
APA StyleAnders, F., Liu, A., Mann, C., Teister, J., Lauzi, J., Thanos, S., Grus, F. H., Pfeiffer, N., & Prokosch, V. (2017). The Small Heat Shock Protein α-Crystallin B Shows Neuroprotective Properties in a Glaucoma Animal Model. International Journal of Molecular Sciences, 18(11), 2418. https://doi.org/10.3390/ijms18112418