Impact of Unsaturated Fatty Acids on Cytokine-Driven Endothelial Cell Dysfunction



Abstract
:1. Introduction
2. Results
2.1. Fatty Acid Composition
2.2. Cell Viability
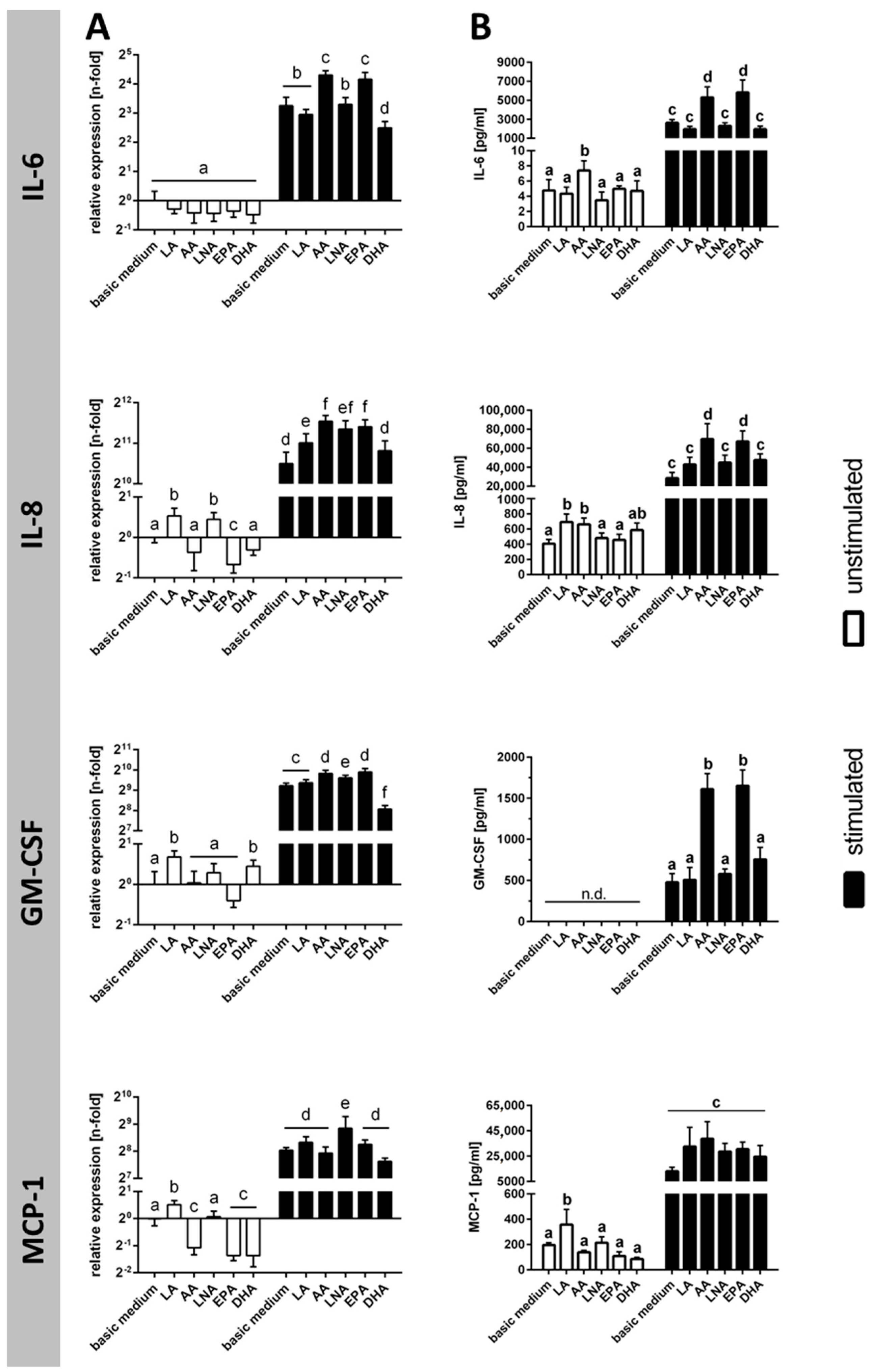
2.3. Cytokines
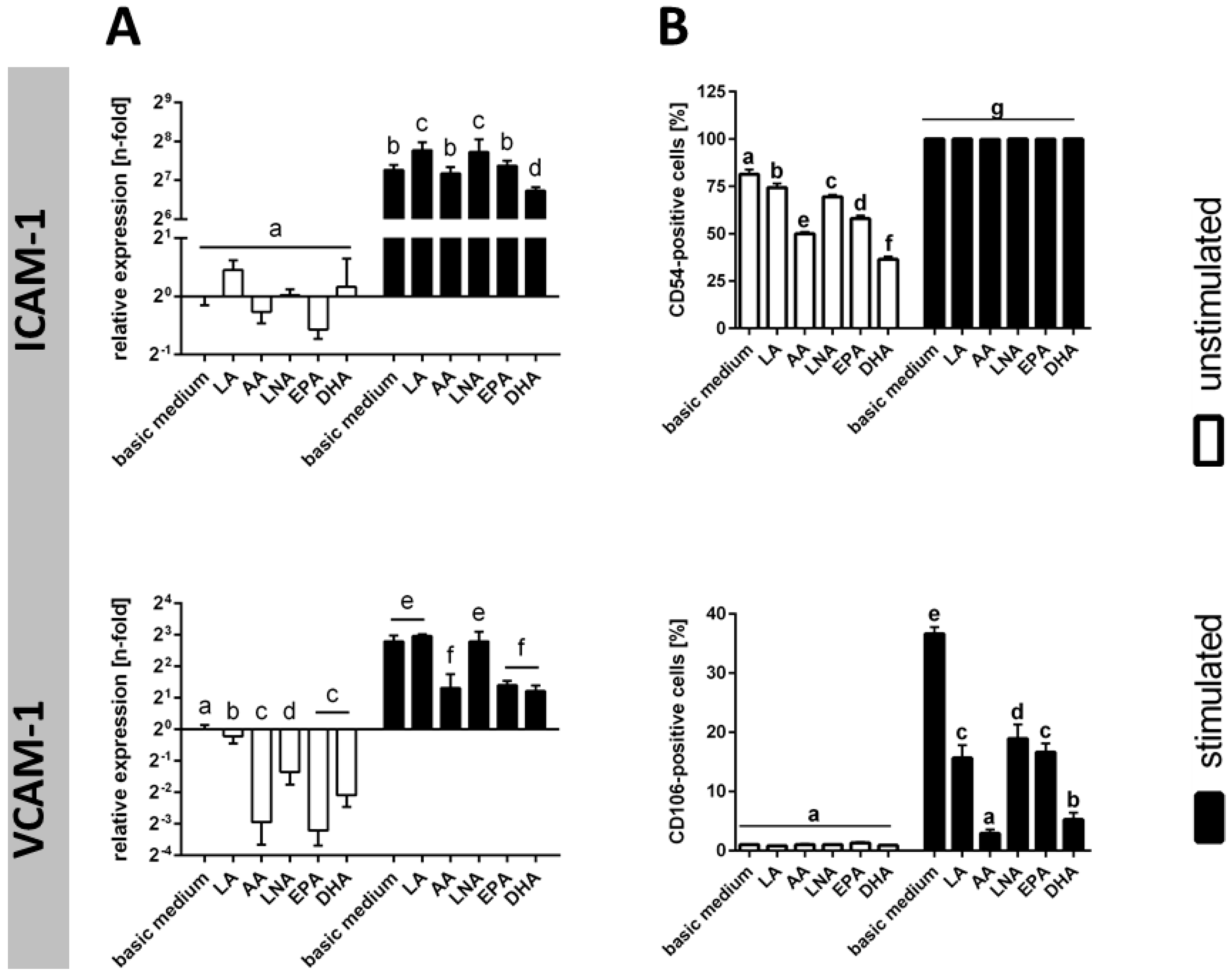
2.4. Adhesion Molecules
2.5. Coagulation Factors
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture, Fatty Acid Supplementation, and Stimulation
4.3. Gas Chromatography
4.4. Viability Assay
4.5. Quantitative Real-Time PCR
4.6. ELISA
4.7. Flow Cytometry
4.8. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Krishnaswamy, G.; Kelley, J.; Yerra, L.; Smith, J.K.; Chi, D.S. Human endothelium as a source of multifunctional cytokines: Molecular regulation and possible role in human disease. J. Interferon Cytokine Res. 1999, 19, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Aird, W.C. The role of the endothelium in severe sepsis and multiple organ dysfunction syndrome. Blood 2003, 101, 3765–3777. [Google Scholar] [CrossRef] [PubMed]
- Cepinskas, G.; Wilson, J.X. Inflammatory response in microvascular endothelium in sepsis: Role of oxidants. J. Clin. Biochem. Nutr. 2008, 42, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Opitz, B.; Eitel, J.; Meixenberger, K.; Suttorp, N. Role of Toll-like receptors, NOD-like receptors and RIG-I-like receptors in endothelial cells and systemic infections. Thromb. Haemostasis 2009, 102, 1103–1109. [Google Scholar] [CrossRef] [PubMed]
- Sprague, A.H.; Khalil, R.A. Inflammatory cytokines in vascular dysfunction and vascular disease. Biochem. Pharmacol. 2009, 78, 539–552. [Google Scholar] [CrossRef] [PubMed]
- Legrand, M.; Klijn, E.; Payen, D.; Ince, C. The response of the host microcirculation to bacterial sepsis: Does the pathogen matter? J. Mol. Med. 2010, 88, 127–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, K.; Unger, R.E.; Brunner, J.; Kirkpatrick, C.J. Molecular basis of endothelial dysfunction in sepsis. Cardiovasc. Res. 2003, 60, 49–57. [Google Scholar] [CrossRef]
- Lee, W.L.; Liles, W.C. Endothelial activation, dysfunction and permeability during severe infections. Curr. Opin. Hematol. 2011, 18, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Noll, G. Pathogenesis of atherosclerosis: A possible relation to infection. Atherosclerosis 1998, 140, S3–S9. [Google Scholar] [CrossRef]
- Kaperonis, E.A.; Liapis, C.D.; Kakisis, J.D.; Dimitroulis, D.; Papavassiliou, V.G. Inflammation and atherosclerosis. Eur. J. Vasc. Endovasc. Surg. 2006, 31, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Botham, K.M.; Wheeler-Jones, C.P.D. Postprandial lipoproteins and the molecular regulation of vascular homeostasis. Prog. Lipid Res. 2013, 52, 446–464. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. The relationship between the fatty acid composition of immune cells and their function. Prostaglandins Leukotrienes Essent. Fatty Acids 2008, 79, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Chiang, N.; van Dyke, T.E. Resolving inflammation: Dual anti-inflammatory and pro-resolution lipid mediators. Nature Rev. Immunol. 2008, 8, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.Y.; Talukdar, S.; Bae, E.J.; Imamura, T.; Morinaga, H.; Fan, W.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. GPR120 Is an Omega-3 Fatty Acid Receptor Mediating Potent Anti-inflammatory and Insulin-Sensitizing Effects. Cell 2010, 142, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Schumann, J.; Fuhrmann, H. Impairment of NFκB activity by unsaturated fatty acids. Int. Immunopharmacol. 2010, 10, 978–984. [Google Scholar] [CrossRef] [PubMed]
- Wanten, G.J.A.; Calder, P.C. Immune modulation by parenteral lipid emulsions. Am. J. Clin. Nutr. 2007, 85, 1171–1184. [Google Scholar] [PubMed]
- Cao, S.; Ren, J.; Sun, L.; Gu, G.; Yuan, Y.; Li, J. Fish oil-supplemented parenteral nutrition prolongs survival while beneficially altering phospholipids’ Fatty Acid composition and modulating immune function in rat sepsis. Shock 2011, 36, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.; Sata, M.; Fukuda, D.; Tanaka, K.; Soma, M.; Hirata, Y.; Nagai, R. Orally administered eicosapentaenoic acid reduces and stabilizes atherosclerotic lesions in ApoE-deficient mice. Atherosclerosis 2008, 197, 524–533. [Google Scholar] [CrossRef] [PubMed]
- Casós, K.; Sáiz, M.P.; Ruiz-Sanz, J.I.; Mitjavila, M.T. Atherosclerosis prevention by a fish oil-rich diet in apoE−/− mice is associated with a reduction of endothelial adhesion molecules. Atherosclerosis 2008, 201, 306–317. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lim, E.-J.; Toborek, M.; Hennig, B. The role of fatty acids and caveolin-1 in tumor necrosis factor alpha-induced endothelial cell activation. Metab. Clin. Exp. 2008, 57, 1328–1339. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, R.A.; Harvey, K.A.; Ruzmetov, N.; Miller, S.J.; Zaloga, G.P. n-3 Fatty acids prevent whereas trans-fatty acids induce vascular inflammation and sudden cardiac death. Br. J. Nutr. 2009, 102, 1811–1819. [Google Scholar] [CrossRef] [PubMed]
- Majkova, Z.; Toborek, M.; Hennig, B. The role of caveolae in endothelial cell dysfunction with a focus on nutrition and environmental toxicants. J. Cell. Mol. Med. 2010, 14, 2359–2370. [Google Scholar] [CrossRef] [PubMed]
- Layne, J.; Majkova, Z.; Smart, E.J.; Toborek, M.; Hennig, B. Caveolae: A regulatory platform for nutritional modulation of inflammatory diseases. J. Nutr. Biochem. 2011, 22, 807–811. [Google Scholar] [CrossRef] [PubMed]
- Chong, D.L.W.; Sriskandan, S. Pro-inflammatory mechanisms in sepsis. Contrib. Microbiol. 2011, 17, 86–107. [Google Scholar] [PubMed]
- Mayer, K.; Schmidt, R.; Muhly-Reinholz, M.; Bogeholz, T.; Gokorsch, S.; Grimminger, F.; Seeger, W. In vitro mimicry of essential fatty acid deficiency in human endothelial cells by TNFα impact of ω-3 versus ω-6 fatty acids. J. Lipid Res. 2002, 43, 944–951. [Google Scholar] [PubMed]
- Kelley, E.E.; Buettner, G.R.; Burns, C.P. Relative alpha-tocopherol deficiency in cultured cells: Free radical-mediated lipid peroxidation, lipid oxidizability, and cellular polyunsaturated fatty acid content. Arch. Biochem. Biophys. 1995, 319, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Schefe, J.H.; Lehmann, K.E.; Buschmann, I.R.; Unger, T.; Funke-Kaiser, H. Quantitative real-time RT-PCR data analysis: Current concepts and the novel “gene expression’s CT difference” formula. J. Mol. Med. 2006, 84, 901–910. [Google Scholar] [CrossRef] [PubMed]
- Ruijter, J.M.; Ramakers, C.; Hoogaars, W.M.H.; Karlen, Y.; Bakker, O.; van den Hoff, M.J.B.; Moorman, A.F.M. Amplification efficiency: Linking baseline and bias in the analysis of quantitative PCR data. Nucleic Acids Res. 2009, 37, e45. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid | Basic Medium | LNA | EPA | DHA | LA | AA |
|---|---|---|---|---|---|---|
| C18:3n3 | n.d. | 108.2 ± 9.7 b | 0.9 ± 0.1 a | 0.4 ± 0.1 a | n.d. | 0.9 ± 0.2 a |
| C20:3n3 | 0.2 ± 0.2 a | 37.2 ± 4.9 b | 0.6 ± 0.1 a | 0.5 ± 0.1 a | 0.3 ± 0.1 a | 0.8 ± 0.2 a |
| C20:4n3 | 1.2 ± 0.2 a | 49.6 ± 4.2 c | 15.1 ± 0.7 b | 2.5 ± 0.2 a | 0.4 ± 0.1 a | 1.7 ± 0.5 a |
| C20:5n3 | 8.9 ± 0.5 a | 80.9 ± 8.2 c | 116.0 ± 3.8 d | 33.8 ± 3.4 b | 1.5 ± 0.1 a | 3.2 ± 0.6 a |
| C22:4n3 | n.d. | 7.5 ± 0.8 c | 1.8 ± 0.1 b | 1.0 ± 0.2 a | n.d. | n.d. |
| C22:5n3 | 37.1 ± 2.6 a | 134.5 ± 14.5 b | 242.3 ± 10.0 c | 40.7 ± 5.9 a | 28.3 ± 3.3 a | 35.5 ± 3.6 a |
| C22:6n3 | 42.4 ± 2.9 a | 42.2 ± 4.6 a | 62.5 ± 2.5 b | 129.7 ± 15.4 c | 30.2 ± 3.6 a | 23.4 ± 5.2 a |
| C18:2n6 | 10.6 ± 0.8 a | 16.3 ± 1.5 a | 16.1 ± 0.6 a | 15.2 ± 1.8 a | 139.4 ± 17.3 b | 20.9 ± 4.0 a |
| C18:3n6 | 1.2 ± 0.2 a | 1.7 ± 0.2 a | 2.0 ± 0.2 a | 1.4 ± 0.1 a | 5.3 ± 0.7 b | 14.3 ± 1.6 c |
| C20:3n6 | 13.7 ± 1.3 a | 10.2 ± 1.1 a | 10.8 ± 0.3 a | 16.7 ± 1.8 a | 40.0 ± 4.9 b | 64.4 ± 6.3 b |
| C20:4n6 | 51.2 ± 4.0 a | 31.6 ± 3.0 a | 31.4 ± 1.9 a | 44.6 ± 5.5 a | 97.9 ± 12.3 a | 368.8 ± 85.3 b |
| C22:3n6 | n.d. | n.d. | n.d. | n.d. | 6.1 ± 1.0 a | 6.8 ± 2.4 a |
| C22:4n6 | 9.0 ± 0.6 a | 5.3 ± 0.5 a | 6.3 ± 0.3 a | 5.1 ± 0.9 a | 58.8 ± 7.0 b | 303.0 ± 60.4 c |
| C22:5n6 | 3.0 ± 0.2 a | 1.4 ± 0.3 a | 1.9 ± 0.2 a | 2.3 ± 0.3 a | 25.4 ± 3.1 b | 35.5 ± 11.3 c |
| Total fatty acids | 967 ± 75 a | 1146 ± 105 a | 1234 ± 45 a | 1105 ± 138 a | 1118 ± 137 a | 1681 ± 214 b |
| Total saturated | 380 ± 28 a | 423 ± 37 a | 507 ± 19 b | 470 ± 59 b | 466 ± 58 b | 552 ± 56 b |
| Total n3 | 90 ± 6 a | 467 ± 45 c | 442 ± 16 c | 210 ± 25 b | 61 ± 7 a | 67 ± 9 a |
| Total n6 | 87 ± 6 a | 66 ± 6 a | 69 ± 2 a | 85 ± 10 a | 365 ± 41 b | 779 ± 164 c |
| Total n7 | 91 ± 8 a | 50 ± 5 b | 56 ± 2 b | 82 ± 10 a | 52 ± 6 b | 64 ± 7 b |
| Total n9 | 293 ± 26 a | 124 ± 12 c | 144 ± 5 c | 237 ± 30 b | 134 ± 16 c | 163 ± 16 c |
| MBI | 76 ± 2 a | 152 ± 2 d | 163 ± 1 e | 111 ± 2 c | 102 ± 1 b | 160 ± 6 e |
| Target | Primer Sequence (5′→3′) | Product Size (bp) | Annealing Temperature (°C) | Extension Time (s) |
|---|---|---|---|---|
| IL-6 | AAGCCAGAGCTGTGCAGATG CTGGCATTTGTGGTTGGGTC | 106 | 60 | 10 |
| IL-8 | CCTGATTTCTGCAGCTCTGTG CCAGACAGAGCTCTCTTCCAT | 197 | 56 | 20 |
| GM-CSF | CCATGATGGCCAGCCACTAC CTGGCTCCCAGCAGTCAAAG | 141 | 60 | 20 |
| MCP-1 | GTCTCTGCCGCCCTTCTGTGC AACAGCAGGTGACTGGGGCA | 100 | 60 | 10 |
| PAI-1 | CAGACCAAGAGCCTCTCC ATCACTTGGCCCATGAAAAG | 202 | 54 | 20 |
| vWF | GGCAATTCCTTCCTCCACAAAC CAGTTGACCCGATGACTCTTCA | 167 | 61 | 20 |
| TF | CACAGAGTGTGACCTCACCG ATTGTTGGCTGTCCGAGGTT | 177 | 60 | 20 |
| ICAM-1 | CTGATGGGCAGTCAACAGCTA GCAGCGTAGGGTAAGGTTCT | 116 | 60 | 10 |
| VCAM-1 | AGTCCCTGGAAACCAAGAGT TGCAGCTTTGTGGATGGATT | 199 | 58 | 20 |
| β-actin | GCACAGAGCCTCGCCTT CCTTGCACATGCCGGAG | 112 | 61 | 20 |
| GAPDH | CTCAACACGGGAAACCTCAC CGGACATCTAAGGGCATCAC | 268 | 56 | 20 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trommer, S.; Leimert, A.; Bucher, M.; Schumann, J. Impact of Unsaturated Fatty Acids on Cytokine-Driven Endothelial Cell Dysfunction. Int. J. Mol. Sci. 2017, 18, 2739. https://doi.org/10.3390/ijms18122739
Trommer S, Leimert A, Bucher M, Schumann J. Impact of Unsaturated Fatty Acids on Cytokine-Driven Endothelial Cell Dysfunction. International Journal of Molecular Sciences. 2017; 18(12):2739. https://doi.org/10.3390/ijms18122739
Chicago/Turabian StyleTrommer, Simon, Anja Leimert, Michael Bucher, and Julia Schumann. 2017. "Impact of Unsaturated Fatty Acids on Cytokine-Driven Endothelial Cell Dysfunction" International Journal of Molecular Sciences 18, no. 12: 2739. https://doi.org/10.3390/ijms18122739
APA StyleTrommer, S., Leimert, A., Bucher, M., & Schumann, J. (2017). Impact of Unsaturated Fatty Acids on Cytokine-Driven Endothelial Cell Dysfunction. International Journal of Molecular Sciences, 18(12), 2739. https://doi.org/10.3390/ijms18122739