
Dectin-1-Mediated Pathway Contributes to Fusarium proliferatum-Induced CXCL-8 Release from Human Respiratory Epithelial Cells

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
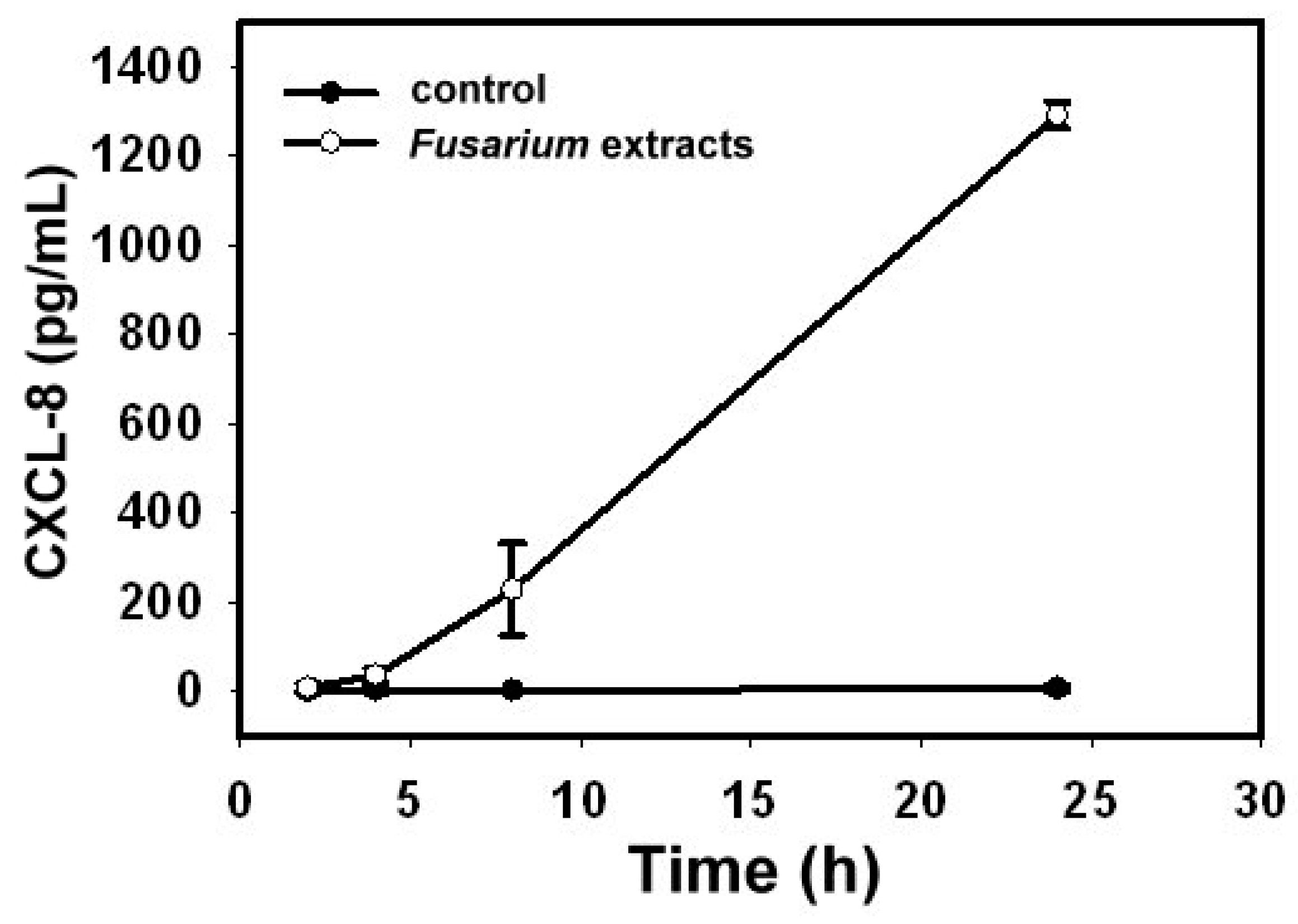
2.1. Fusarium proliferatum-Induced Cytokines Release by BEAS-2B Cells
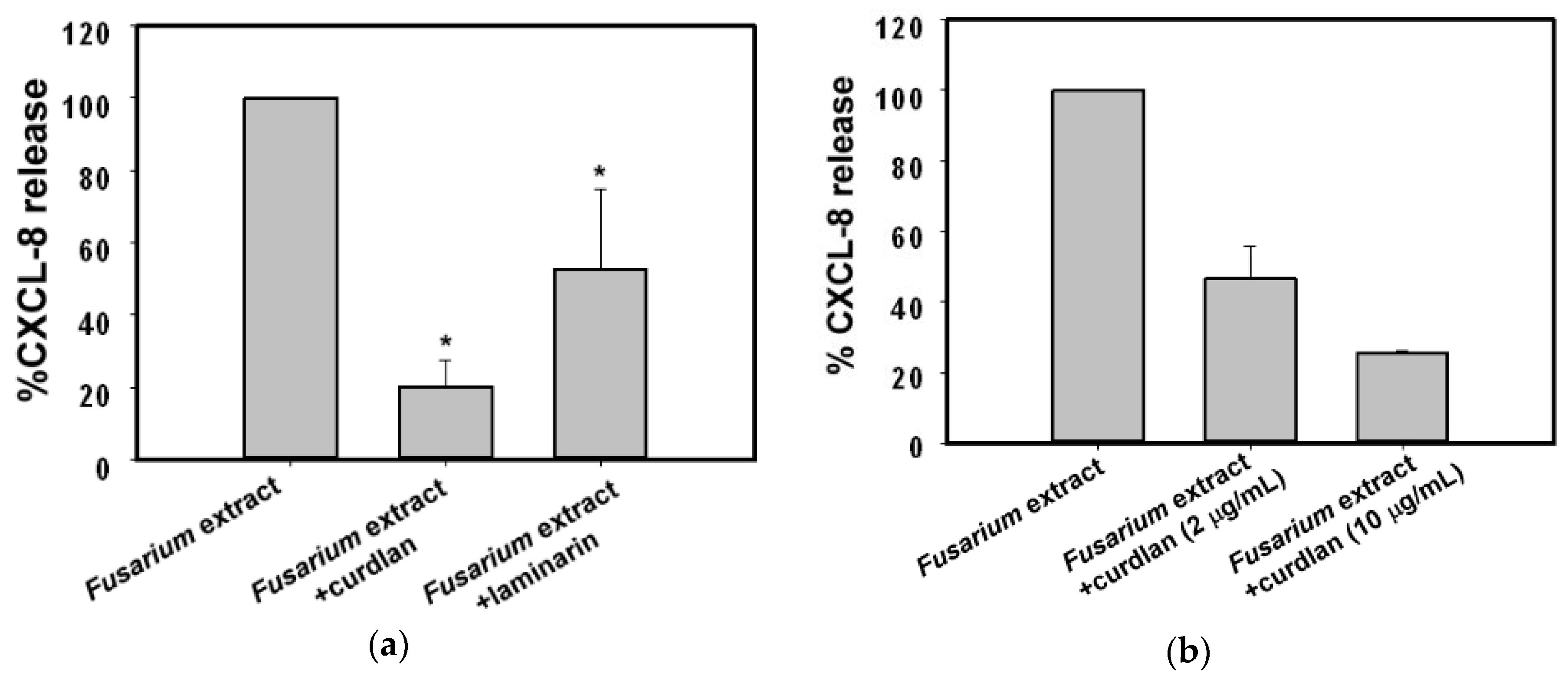
2.2. The Role of β-Glucan Structure in Fusarium proliferatum-Induced CXCL-8 Release by BEAS-2B Cells
2.3. Role of Dectin-1/Spleen Tyrosine Kinase in the Fusarium proliferatum Extract-Induced CXCL-8 Release
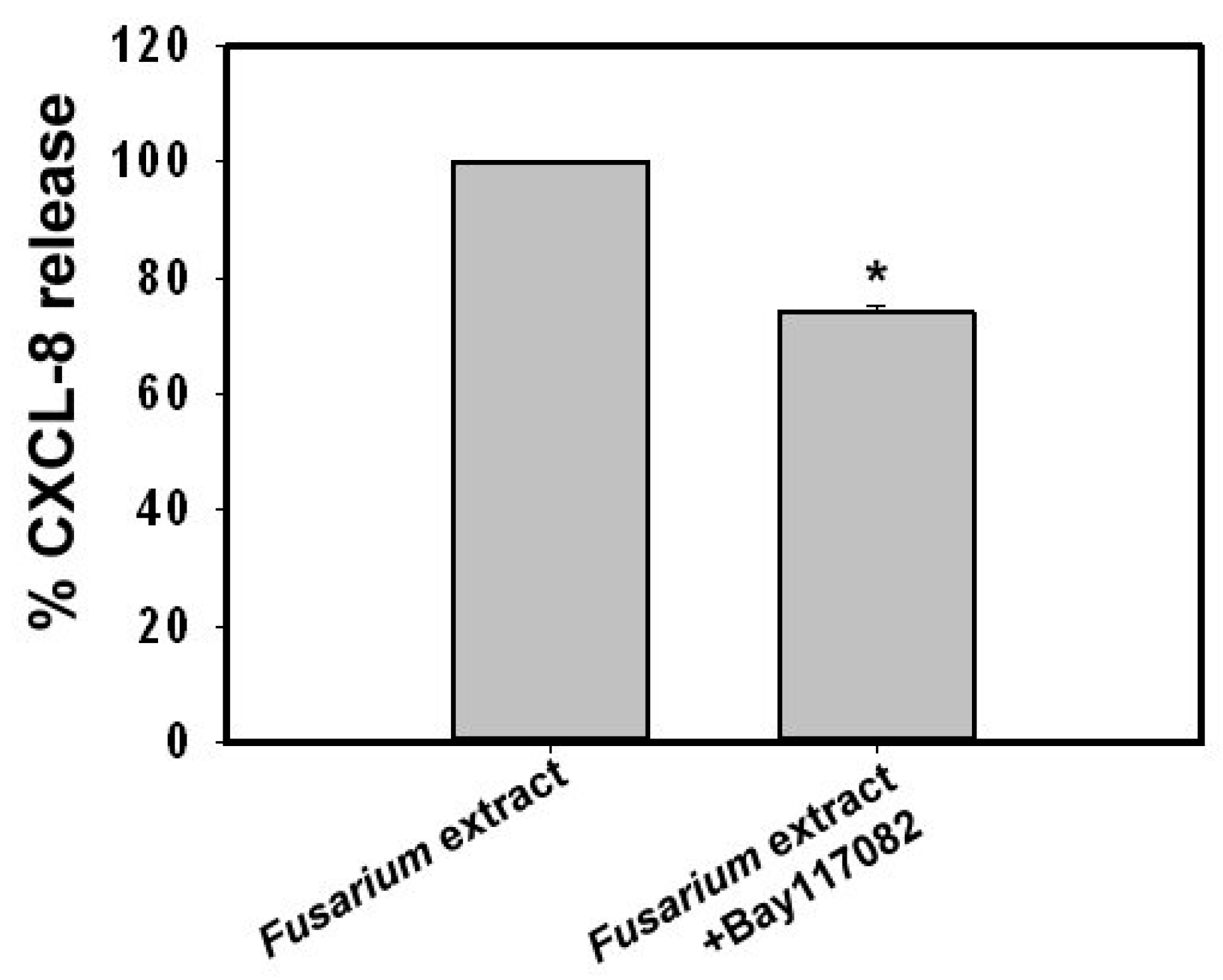
2.4. Involvement of the Mitogen-Activated Protein Kinases, phosphatidylinositol-3-kinase and Nuclear Factor κ-Light-Chain-Enhancer of activated B cells in Fusarium proliferatum-Induced CXCL8 Release
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Crude Extracts of Fusarium proliferatum
4.3. Cell Culture and Treatment
4.4. Cytokine Array and CXCL-8 ELISA Assay
4.5. Statistical Analyses
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| Syk | Spleen tyrosine kinase |
| MAPKs | Mitogen-activated protein kinases |
| PI3K | Phosphatidylinositol-3-kinase |
| HDM | House dust mite |
References
- Sharpe, R.A.; Bearman, N.; Thornton, C.R.; Husk, K.; Osborne, N.J. Indoor fungal diversity and asthma: A meta-analysis and systematic review of risk factors. J. Allergy Clin. Immunol. 2015, 135, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Simon-Nobbe, B.; Denk, U.; Pöll, V.; Rid, R.; Breitenbach, M. The spectrum of fungal allergy. Int. Arch. Allergy Immunol. 2008, 145, 58–86. [Google Scholar] [CrossRef] [PubMed]
- Lehrer, S.B.; Aukrust, L.; Salvaggio, J.E. Respiratory allergy induced by fungi. Clin. Chest 1983, 4, 23–41. [Google Scholar]
- Kauffman, H.F. Interaction of environmental allergens with airway epithelium as a key component of asthma. Curr. Allergy Asthma Rep. 2003, 3, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Tai, H.Y.; Tam, M.F.; Chou, H.; Peng, H.J.; Su, S.N.; Perng, D.W.; Shen, H.D. Pen ch 13 allergen induces secretion of mediators and degradation of occludin protein of human lung epithelial cells. Allergy 2006, 61, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Kauffman, H.F.; Tomee, J.F.; van der Werf, T.S.; de Monchy, J.G.; Koëter, G.K. Review of fungus-induced asthmatic reactions. Am. J. Respir. Crit. Care Med. 1995, 151, 2109–2115. [Google Scholar] [CrossRef] [PubMed]
- Carsin, A.; Mazenq, J.; Ilstad, A.; Dubus, J.C.; Chanez, P.; Gras, D. Bronchial epithelium in children: A key player in asthma. Eur. Respir. Rev. 2016, 25, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Chou, H.; Wu, K.G.; Yeh, C.C.; Tai, H.Y.; Tam, M.F.; Chen, Y.S.; Shen, H.D. The transaldolase, a novel allergen of Fusarium proliferatum, demonstrates IgE cross-reactivity with its human analogue. PLoS ONE 2014, 9, e103488. [Google Scholar] [CrossRef] [PubMed]
- Stroud, R.; Calhoun, K.; Wright, S.; Kennedy, K. Prevalence of hypersensitivity to specific fungal allergens as determined by intradermal dilutional testing. Otolaryngol. Head Neck Surg. 2001, 125, 491–494. [Google Scholar] [CrossRef]
- Yeh, C.C.; Tai, H.Y.; Chou, H.; Wu, K.G.; Shen, H.D. Vacuolar serine protease is a major allergen of Fusarium proliferatum and an IgE-cross reactive pan-fungal allergen. Allergy Asthma Immunol. Res. 2016, 8, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.-C.; Horng, H.-C.; Chou, H.; Tai, H.-Y.; Shen, H.-D.; Hsieh, S.-L.; Wang, P.-H. Department of Obstetrics and Gynecology, Taipei Veterans General Hospital, Taipei 112, Taiwan. BEAS-2B cells and Fusarium proliferatum extracts. 2016. [Google Scholar]
- Mocanu, G.; Mihai, D.; Moscovici, M.; Picton, L.; LeCerf, D. Curdlan microspheres. Synthesis, characterization and interaction with proteins (enzymes, vaccines). Int. J. Biol. Macromol. 2009, 44, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Osorio, F.; Reise Sousa, C. Myeloid C-type lectin receptors in pathogen recognition and host defense. Immunity 2011, 34, 651–664. [Google Scholar] [CrossRef] [PubMed]
- Kerrigan, A.M.; Brown, G.D. Syk-coupled C-type lectins in immunity. Trends Immunol. 2011, 32, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Saijo, S.; Wakura, Y. Dectin-1 and Dectin-2 in innate immunity against fungi. Int. Immunol. 2011, 23, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Koyasu, S. The role of PI3K in immune cells. Nat. Immunol. 2003, 4, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Kingeter, L.M.; Lin, X. C-type lectin receptor-induced NF-κB activation in innate immune and inflammatory responses. Cell Mol. Immunol. 2012, 9, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Amend, A.S.; Seifert, K.A.; Samson, R.; Bruns, T.D. Indoor fungal composition is geographically patterned and more diverse in temperate zones than in the tropics. Proc. Natl. Acad. Sci. USA 2010, 107, 13748–13753. [Google Scholar] [CrossRef] [PubMed]
- Romani, L. Immunity to fungal infections. Nat. Rev. Immunol. 2011, 11, 275–288. [Google Scholar] [CrossRef] [PubMed]
- Douwes, J. (1→3)-β-d-glucans and respiratory health: A review of the scientific evidence. Indoor Air 2005, 15, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Schoffelmeer, E.A.; Klis, F.M.; Sietsma, J.H.; Cornelissen, B.J. The cell wall of Fusarium oxysporum. Fungal Genet. Biol. 1999, 27, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Carmona, E.M.; Lamont, J.D.; Xue, A.; Wylam, M.; Limper, A.H. Pneumocystis cell wall β-glucan stimulates calcium-dependent signaling of IL-8 secretion by human airway epithelial cells. Respir. Res. 2010, 11, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Neveu, W.A.; Bernardo, E.; Allard, J.L.; Nagaleekar, V.; Wargo, M.J.; Davis, R.J.; Iwakura, Y.; Whittaker, L.A.; Rincon, M. Fungal allergen β-glucans trigger p38 mitogen-activated protein kinase-mediated IL-6 translation in lung epithelial cells. Am. J. Respir. Cell Mol. Biol. 2011, 45, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- Nathan, A.T.; Peterson, E.A.; Chakir, J.; Wills-Karp, M. Innate immune responses of airway epithelium to house dust mite are mediated through β-glucan-dependent pathways. J. Allergy Clin. Immunol. 2009, 123, 612–618. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.K.; Lu, X.; Li, X.; Sun, Q.Y.; Su, X.; Song, Y.; Sun, H.M.; Shi, Y. Dectin-1 is inducible and plays a crucial role in Aspergillus-induced innate immune responses in human bronchial epithelial cells. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 2755–2764. [Google Scholar] [CrossRef] [PubMed]
- Heyl, K.A.; Klassert, T.E.; Heinrich, A.; Müller, M.M.; Klaile, E.; Dienemann, H.; Grünewald, C.; Bals, R.; Singer, B.B.; Slevogt, H. Dectin-1 is expressed in human lung and mediates the proinflammatory immune response to nontypeable Haemophilus influenzae. MBio 2014, 5, e01492-14. [Google Scholar] [CrossRef] [PubMed]
- Goodridge, H.S.; Wolf, A.J.; Underhill, D.M. β-Glucan recognition by the innate immune system. Immunol. Rev. 2009, 230, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Goodridge, H.S.; Reyes, C.N.; Becker, C.A.; Katsumoto, T.R.; Ma, J.; Wolf, A.J.; Bose, N.; Chan, A.S.H.; Magee, A.S.; Danielson, M.E.; et al. Activation of the innate immune receptor Dectin-1 upon formation of a “phagocytic synapse”. Nature 2011, 472, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, K.; Muta, T.; Yamazaki, S.; Takeshige, K. Activation of macrophages by linear (1right-arrow3)-β-d-glucan. Impliations for the recognition of fungi by innate immunity. J. Biol. Chem. 2002, 277, 36825–36831. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.C.; Zhao, G.Q.; Lin, J.; Hu, L.T.; Xu, Q.; Peng, X.D.; Wang, X.; Qiu, S. Dectin-1 agonist curdlan modulates innate immunity to Aspergillus fumigatus in human corneal epithelial cells. Int. J. Ophthalmol. 2015, 8, 690–696. [Google Scholar] [PubMed]
- Brown, G.D.; Herre, J.; Williams, D.L.; Willment, J.A.; Marshall, A.S.; Gordon, S. Dectin-1 mediates the biological effects of β-glucans. J. Exp. Med. 2003, 197, 1119–1124. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.D.; Lin, W.L.; Tam, M.F.; Wang, S.R.; Tzean, S.S.; Huang, M.H.; Han, S.H. Characterization of allergens from Penicillium oxalicum and P. notatum by immunoblotting and N-terminal amino acid sequence analysis. Clin. Exp. Allergy 1999, 29, 642–651. [Google Scholar] [CrossRef] [PubMed]






© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yeh, C.-C.; Horng, H.-C.; Chou, H.; Tai, H.-Y.; Shen, H.-D.; Hsieh, S.-L.; Wang, P.-H. Dectin-1-Mediated Pathway Contributes to Fusarium proliferatum-Induced CXCL-8 Release from Human Respiratory Epithelial Cells. Int. J. Mol. Sci. 2017, 18, 624. https://doi.org/10.3390/ijms18030624
Yeh C-C, Horng H-C, Chou H, Tai H-Y, Shen H-D, Hsieh S-L, Wang P-H. Dectin-1-Mediated Pathway Contributes to Fusarium proliferatum-Induced CXCL-8 Release from Human Respiratory Epithelial Cells. International Journal of Molecular Sciences. 2017; 18(3):624. https://doi.org/10.3390/ijms18030624
Chicago/Turabian StyleYeh, Chang-Ching, Huann-Cheng Horng, Hong Chou, Hsiao-Yun Tai, Horng-Der Shen, Shie-Liang Hsieh, and Peng-Hui Wang. 2017. "Dectin-1-Mediated Pathway Contributes to Fusarium proliferatum-Induced CXCL-8 Release from Human Respiratory Epithelial Cells" International Journal of Molecular Sciences 18, no. 3: 624. https://doi.org/10.3390/ijms18030624
APA StyleYeh, C. -C., Horng, H. -C., Chou, H., Tai, H. -Y., Shen, H. -D., Hsieh, S. -L., & Wang, P. -H. (2017). Dectin-1-Mediated Pathway Contributes to Fusarium proliferatum-Induced CXCL-8 Release from Human Respiratory Epithelial Cells. International Journal of Molecular Sciences, 18(3), 624. https://doi.org/10.3390/ijms18030624