
Classical and Novel TSPO Ligands for the Mitochondrial TSPO Can Modulate Nuclear Gene Expression: Implications for Mitochondrial Retrograde Signaling

Abstract
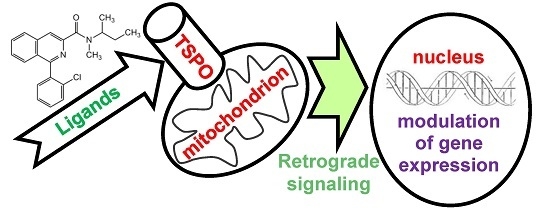
:
1. Introduction
2. Results
2.1. PK 11195 Effects on Gene Expression in General
- Within 15 min, 20 genes significantly changed their expression rate in comparison to vehicle control, and 11 of them code for transcription factors, the other 9 code for proteins, enzymes, and other products.
- After 30 min (indicated in the second column) 14 genes changed significantly their expression rate from vehicle control. At this time point the number of genes coding for transcription factors is 6, the other 8 genes code for proteins, enzymes, and other products.
- After 45 min of PK 11195 exposure (indicated in the third column), 12 genes have their expression changed significantly from vehicle control, 5 of them coding for transcription factors, the other 7 genes code for proteins, enzymes, and other products.
- After 1 h (indicated in the fourth column), 25 genes significantly changed their expression rate, and 14 of them code for transcription factors, the other 11 code for proteins, enzymes, and other products.
- After 3 h (indicated in the fifth column), 14 genes changed significantly their expression rate from vehicle control. The number of genes coding for transcription factors at this time point is 6, the other 8 genes code for proteins, enzymes, and other products.
- After 24 h of PK 11195 exposure (indicated in the sixth column), 29 genes have their expression changed significantly from vehicle control. The number of genes coding for transcription factors is 6 after 24 h of PK 11195 exposure, the remaining majority of the genes (23 genes) codes for proteins, enzymes, and other products at this time point. Indeed, the biggest numbers of gene expression changes for proteins, enzymes, and other products is after 24 h.
2.2. Implied Specific Functional Effects due to Gene Expression Changes Induced by Various PK 11195 Exposure Times
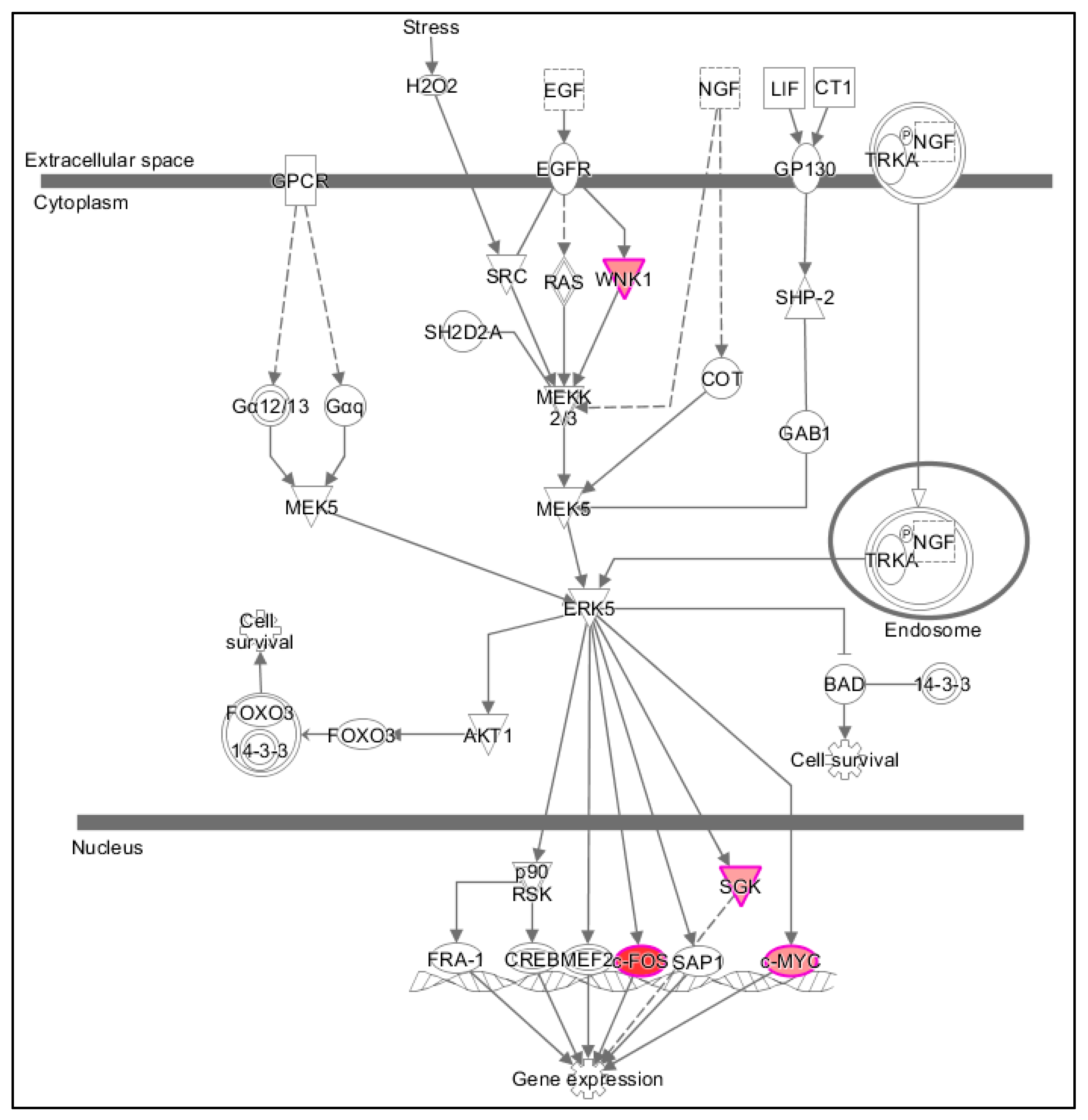
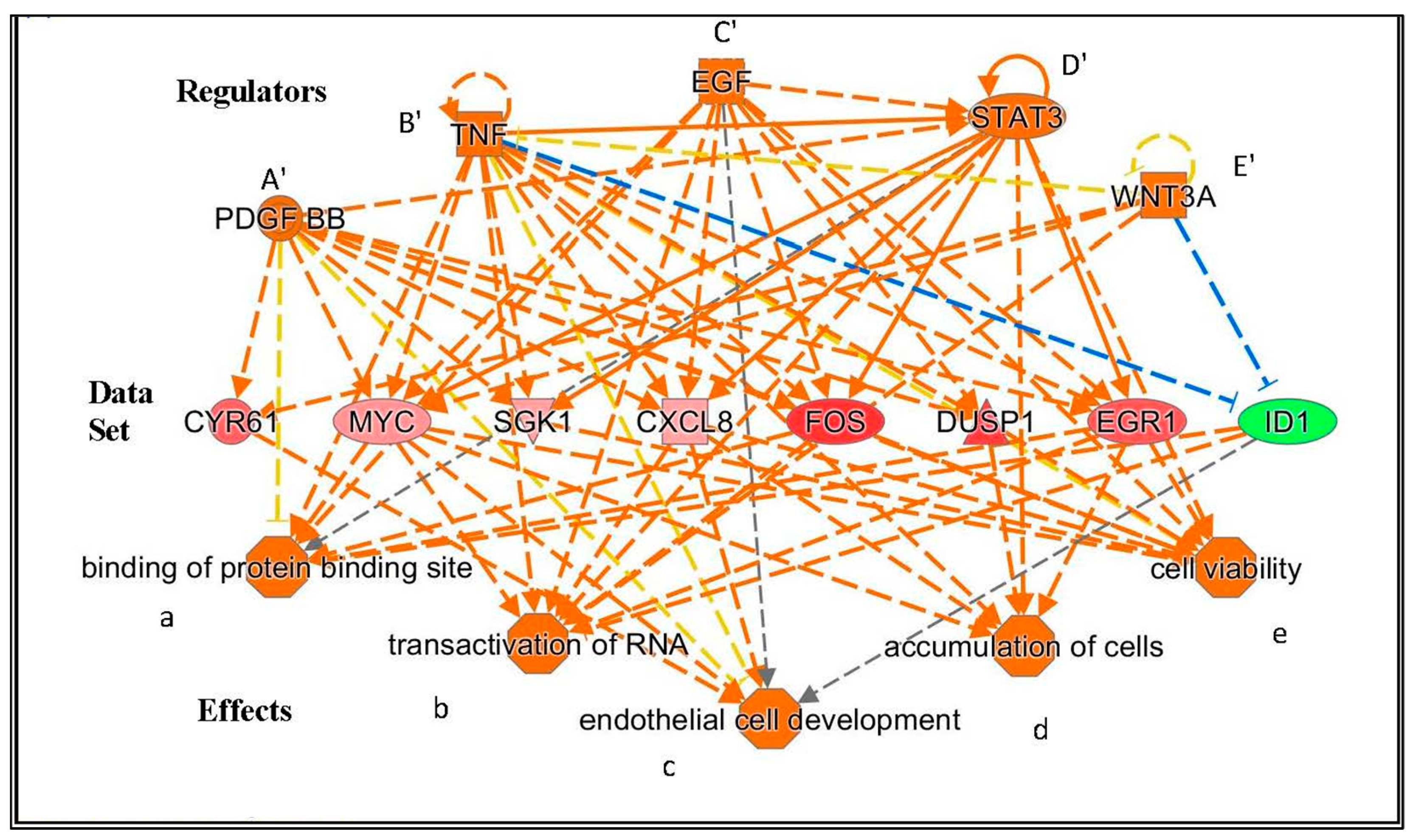
- Regarding functional effects at 15 min, in addition to upregulation of gene expression, in general, several functions appeared to be affected. In particular, binding of protein binding site, transactivation of RNA, cell development, cell viability, and accumulation of cells (Table 4, Figure 3). As seen in Figure 3, 15 min of PK 11195 activates a super-assembly including 5 Regulators, 8 genes, and 5 Effects. In Figure 1 and Figure 2 is shown that PK 11195 application for 15 min affects the canonical pathway for regulation of gene expression.
- Functional effects at 30 min appeared to be more varied than at 15 min of PK 11195 exposure. For simplification, these effects can be classified as: binding of protein binding site, cell division and proliferation, cell viability, metabolism, cell differentiation, cell motility, tumorigenicity, and tissue inflammation. These functions are listed in more detail in Table 4. The super-assembly that can be distinguished at 30 min is elaborate and includes 26 Regulators, 19 genes, and 15 Effects. This super-assembly is provided in the supplementary files.
- After 45 min, cell differentiation effects appeared to be the core functional aspect of the gene expression changes, as well as angiogenesis, proliferation, migration, and cell growth (Table 4). The super-assembly seen at 45 min is relatively small, 9 Regulators, 9 genes, and 3 Effects (provided in the supplementary files). Both the 30 and 45 min of PK 11195 exposures caused gene expression changes associated with the canonical pathway for angiogenesis, also provided in the supplementary files.
- After 1 h, the functional effects in general appeared to include: upregulation of cell cycle, proliferation, cell differentiation, cell viability, and tumorigenesis, but also programmed cell death. These functions are listed in more detail in Table 4. The super-assembly seen at 1 h includes 19 Regulators, 29 genes, and 12 Effects (provided in the supplementary files).
- After 3 h, the general effect due to changes in expression of the various genes after exposure of U118MG cells to 25 µM of PK 11195 appears to imply a less tumorigenic phenotype. The majority of the ‘Effects’ of 3 h of PK 11195 exposure can be classified as down regulation. This down regulation relates to (1) Migration; (2) Inflammatory response; (3) Proliferation, (4) Development, including cell differentiation; (5) Cell viability; and (6) Tumorigenesis. These ‘Effects’ after 3 h are virtually the opposite from those seen after the shorter PK 11195 exposures. These functions are listed in more detail in Table 3. In contrast, programmed cell death is still upregulated, as was also seen after 1 h of PK 11195 exposure. These functions are listed in more detail in Table 4. The super-assembly activated by 3 h of PK 11195 exposure includes 23 Regulators, 30 genes, and 18 Effects, presented in the supplementary files.
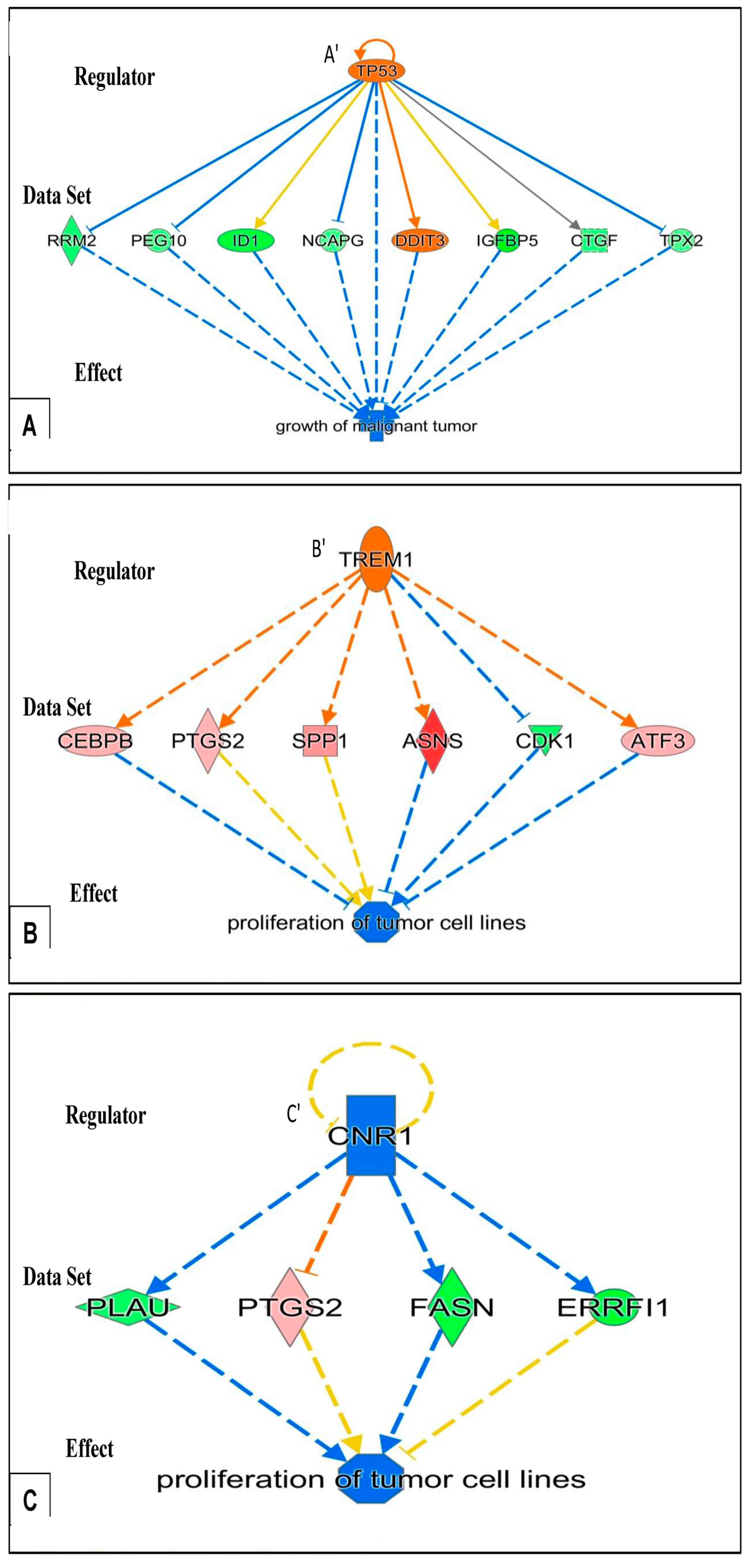
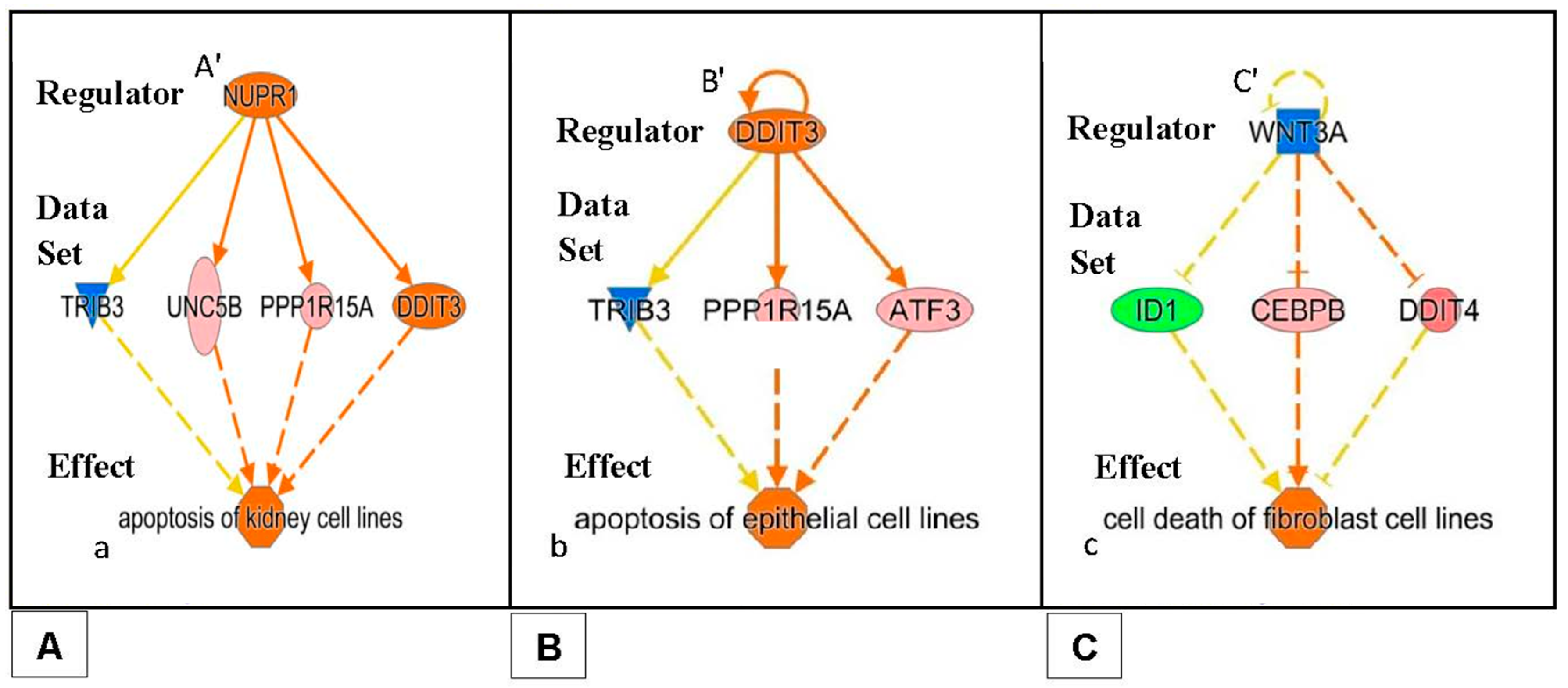
- After 24 h of PK 11195 exposure, pathway analysis with the “Regulator Effects” analytic (IPA®) indicated that due to the significant changes in gene expression only the following general function is down-regulated: tumorigenicity (Figure 4). Several separate pathways were revealed regarding tumorigenicity, each one including just one “Regulator” and a small set of genes forming the “Data Set” (Figure 4A–C). Additional figures in the Supplementary Materials give additional, somewhat more complicated information, i.e., 2 or 3 “Regulators” together modulating ‘Data Sets’ of a dozen to several dozen genes (Supplementary Materials). These figures also impinge on the general theme of reduced tumorigenicity. Thus, after 24 h of PK 11195 exposure no extensive super-assembly was recognized, but several independent assemblies downregulating several aspects of tumorigenicity and upregulation of programmed cell death (Figure 4 and Figure 5).
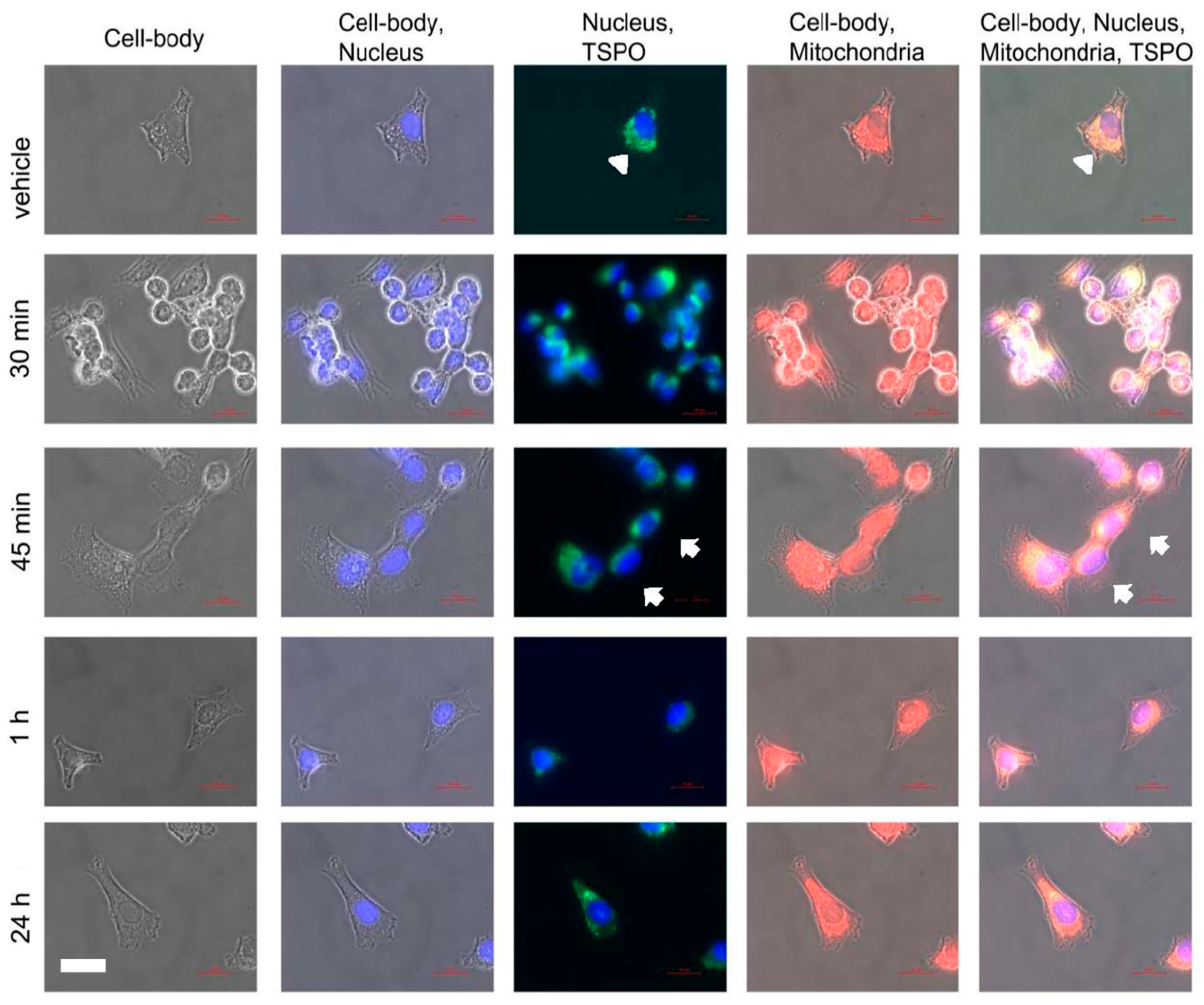
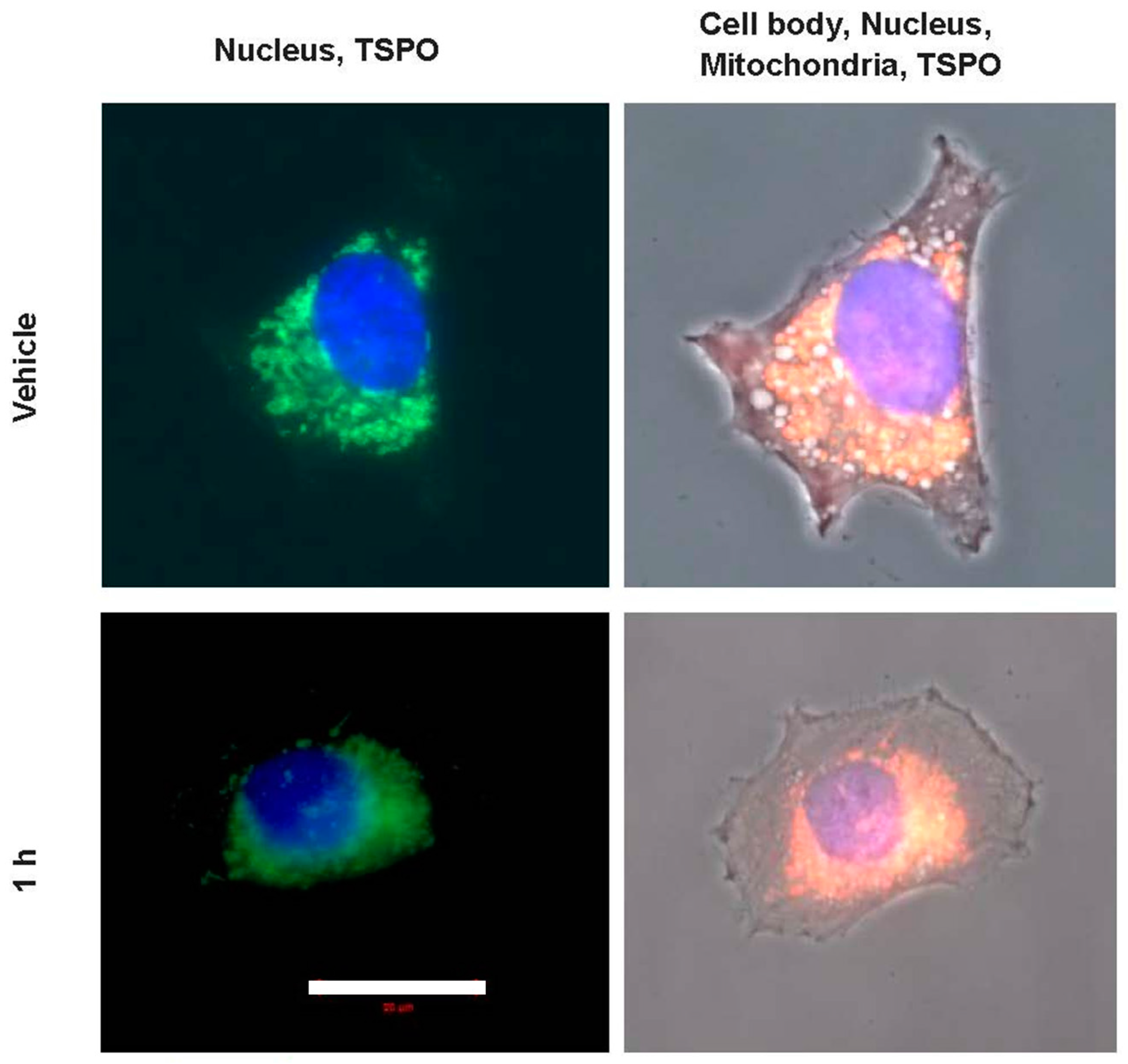
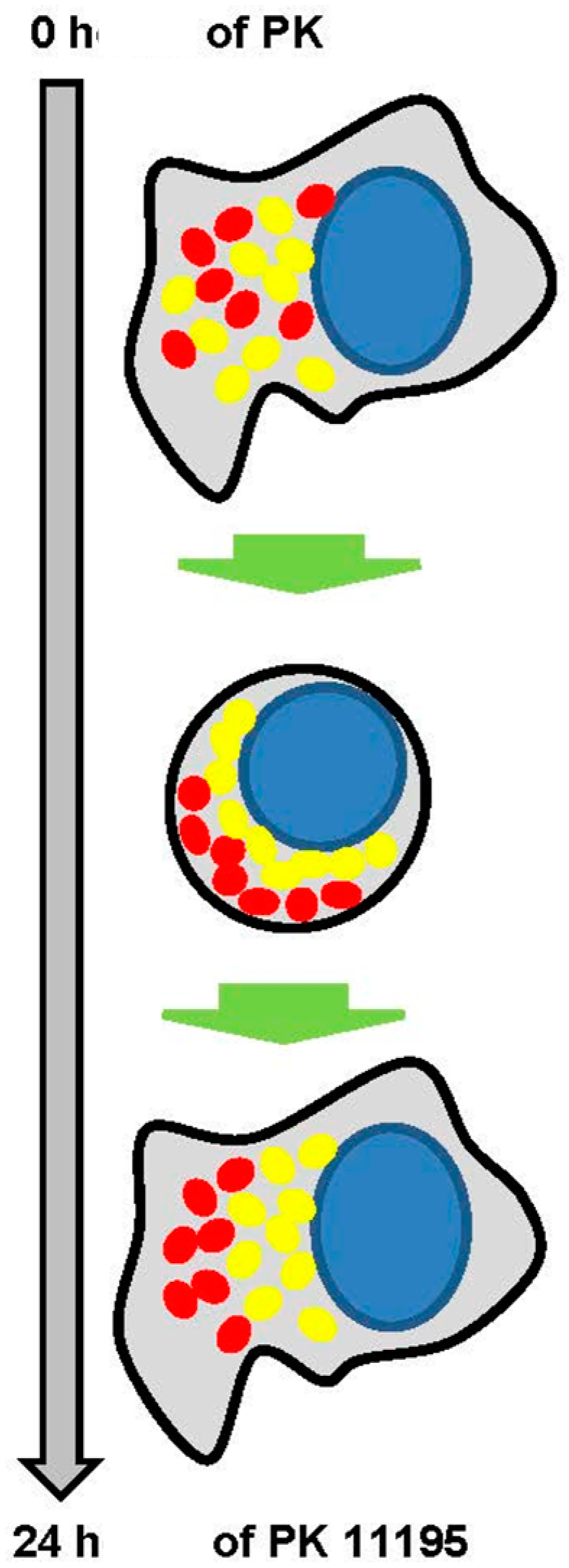
2.3. Microscopic Correlates at Cellular and Intracellular Levels in Association with Changes in Gene Expression due to PK 11195 Exposure
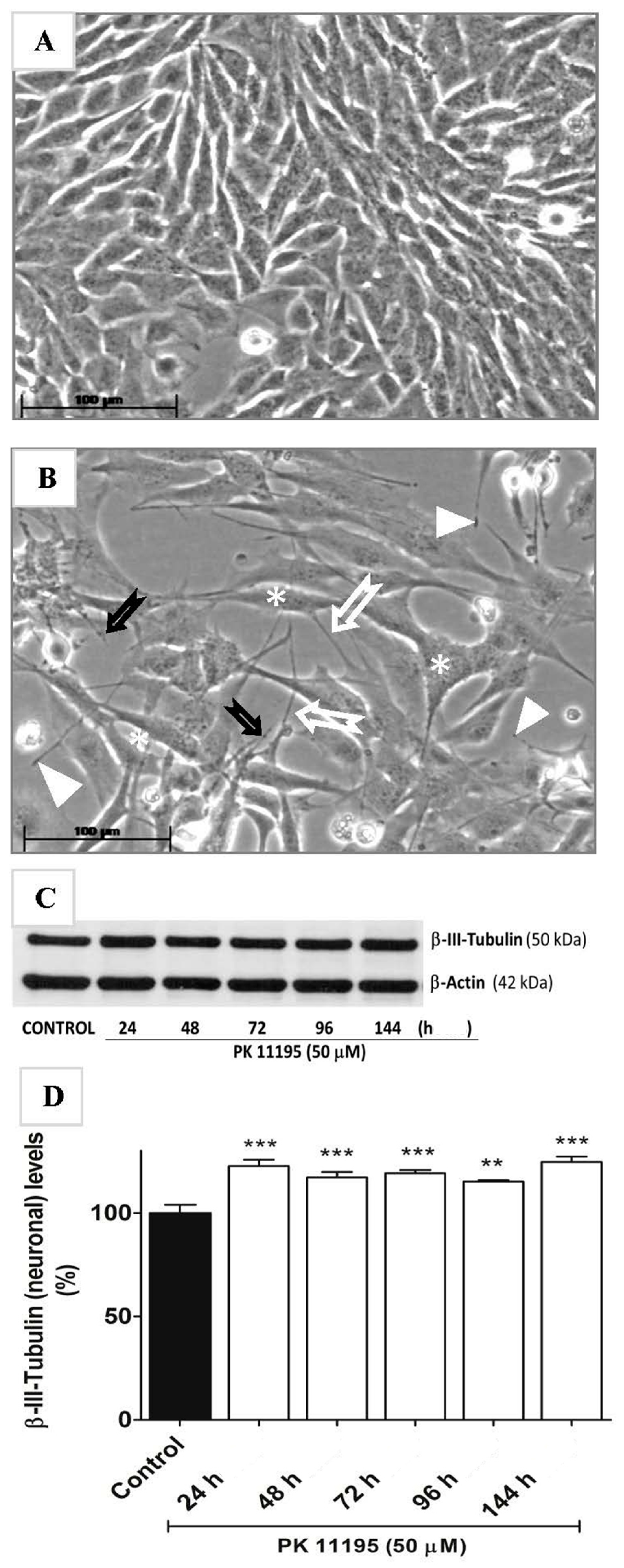
2.4. Actual Phenotypic Effects of PK 11195 Exposure That Were Predicted by Pathway Analysis
2.5. Effects of TSPO Ligands Other Than PK 11195 on Gene Expression
2.6. General Observations
3. Discussion
Caveats and Questions for Future Research
4. Experimental Section
4.1. Materials
Cell Culture
4.2. Exposure to PK 11195 and 2-Cl-MGV-1
4.3. RNA Extraction
4.4. Gene Expression Assay
4.5. Pathway Analysis
4.6. Real-Time RT-PCR
4.7. Microscopic Studies
4.8. Western Blot Analysis of Tubulin Expression in Relation to Neuronal Differentiation
5. Conclusions
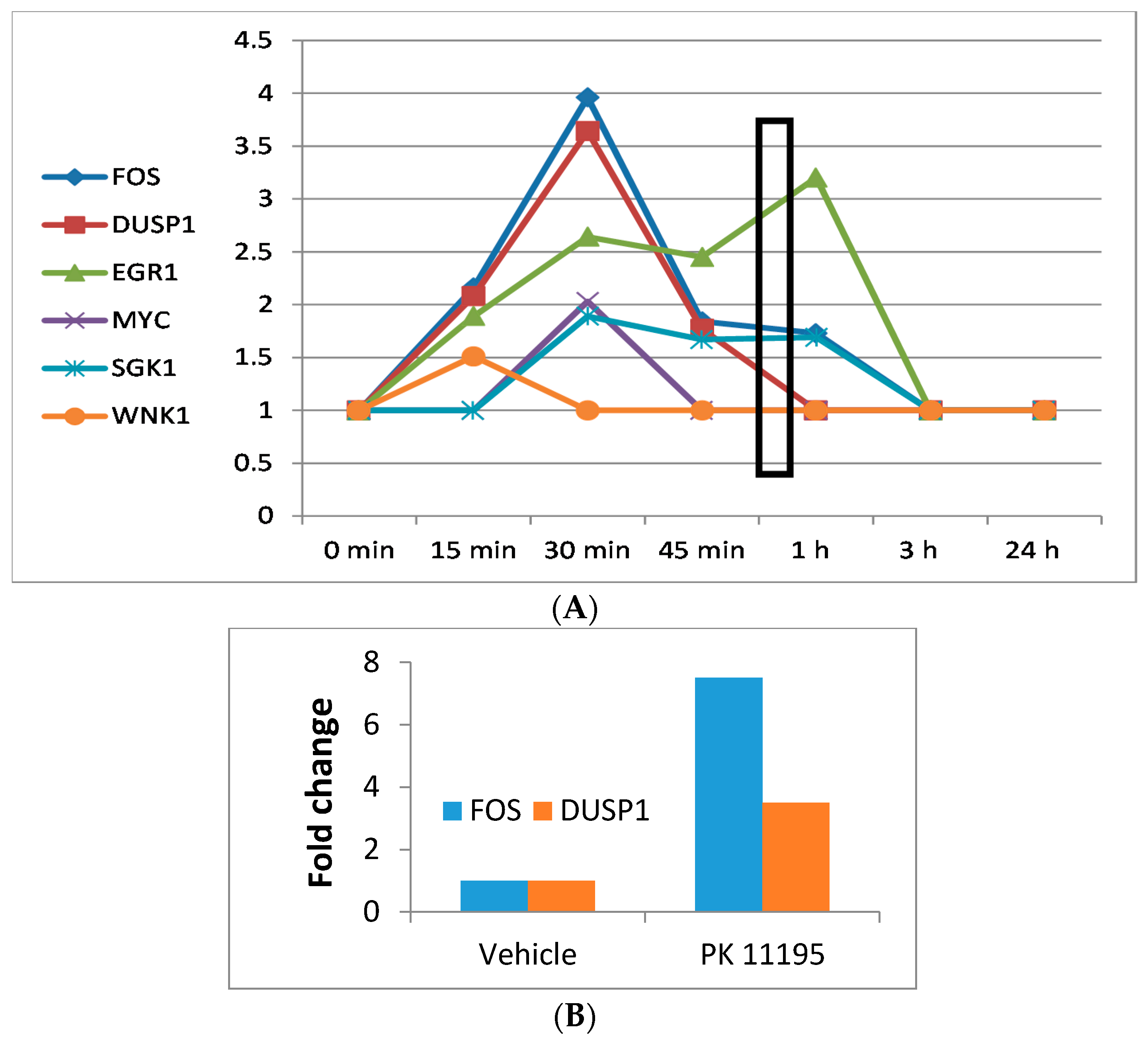
- Our study indicates that the classical TSPO ligand PK 11195 can modulate gene expression in U118MG cells.
- Robust and significant changes in gene expression can already be seen within 15 min and appear to be associated with cell morphological changes within the same time frame.
- At least at 15 min of PK 11195 exposure, expression of several elements of the canonical pathway for regulation of gene expression in U118MG cells is enhanced.
- After 24 h of exposure to PK 11195, changes in gene expression appear to be related to cell viability and tumorigenicity of these U118MG cells.
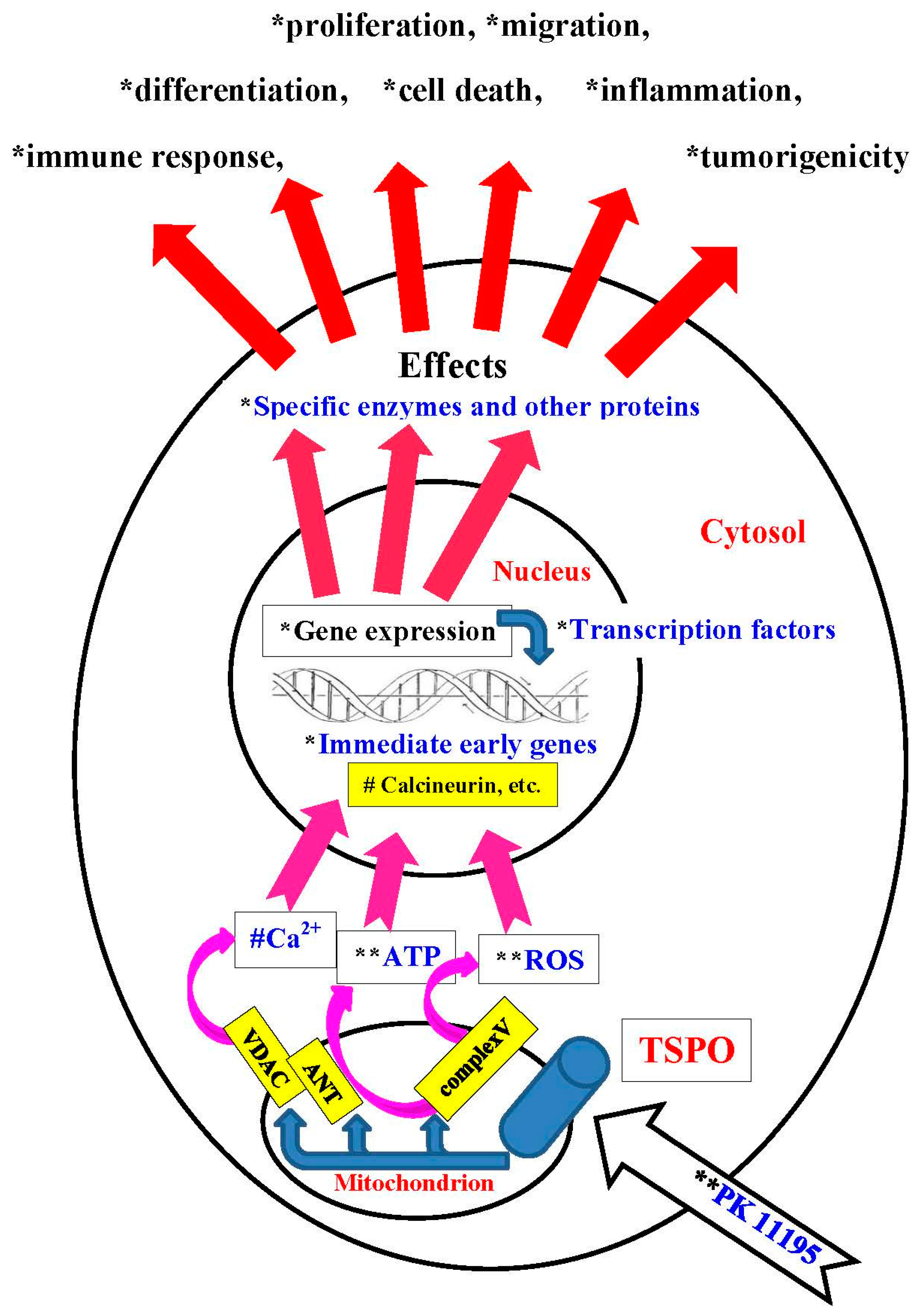
- This modulation in gene expression most likely occurs via mitochondria-to-nucleus signaling, probably via mechanisms including ΔΨm collapse, ROS generation, Ca2+ release, and ATP production (Figure 9). It is well documented by previous studies that ΔΨm collapse, ROS generation, Ca2+ release, and ATP production are under the control of mitochondrial TSPO.
- Thus, TSPO does not just modulate local mitochondrial functions, it also modulates nuclear gene expression.
- Phenotypic changes predicted by the changes in gene expression did actually occur, e.g., cell migration, cell accumulation, cell differentiation, and others.
- The novel TSPO ligand 2-Cl-MGV-1 also specifically modulated gene expression of immediate early genes.
- The modulation of gene expression by the TSPO elucidated in the present study goes a long way in explaining subsequent changes in cellular and organismal functions due to application of TSPO ligands (Figure 9).
- Thus, modulation of nuclear gene expression via the mitochondrial TSPO can induce several vital cell functions, which has major implications for the whole organism in health and disease.
- We believe that our study provides more understanding in the overall biological function of TSPO.
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gavish, M.; Bachman, I.; Shoukrun, R.; Katz, Y.; Veenman, L.; Weisinger, G.; Weizman, A. Enigma of the peripheral benzodiazepine receptor. Pharmacol. Rev. 1999, 51, 629–650. [Google Scholar] [PubMed]
- Veenman, L.; Gavish, M. The peripheral-type benzodiazepine receptor and the cardiovascular system. Implications for drug development. Pharmacol. Ther. 2006, 110, 503–524. [Google Scholar] [CrossRef] [PubMed]
- Veenman, L.; Gavish, M. The role of 18 kDa mitochondrial translocator protein (TSPO) in programmed cell death, and effects of steroids on TSPO expression. Curr. Mol. Med. 2012, 12, 398–412. [Google Scholar] [CrossRef] [PubMed]
- Starosta-Rubinstein, S.; Ciliax, B.J.; Penney, J.B.; McKeever, P.; Young, A.B. Imaging of a glioma using peripheral benzodiazepine receptor ligands. Proc. Natl. Acad. Sci. USA 1987, 84, 891–895. [Google Scholar] [CrossRef] [PubMed]
- Kugler, W.; Veenman, L.; Shandalov, Y.; Leschiner, S.; Spanier, I.; Lakomek, M.; Gavish, M. Ligands of the mitochondrial 18 kDa translocator protein attenuate apoptosis of human glioblastoma cells exposed to erucylphosphohomocholine. Cell. Oncol. 2008, 30, 435–450. [Google Scholar] [PubMed]
- Yeliseev, A.A.; Kaplan, S. TspO of rhodobacter sphaeroides. A structural and functional model for the mammalian peripheral benzodiazepine receptor. J. Biol. Chem. 2000, 275, 5657–5667. [Google Scholar] [PubMed]
- Papadopoulos, V.; Baraldi, M.; Guilarte, T.R.; Knudsen, T.B.; Lacapere, J.J.; Lindemann, P.; Norenberg, M.D.; Nutt, D.; Weizman, A.; Zhang, M.R.; et al. Translocator protein (18 kDa): New nomenclature for the peripheral-type benzodiazepine receptor based on its structure and molecular function. Trends Pharmacol. Sci. 2006, 27, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Veenman, L.; Papadopoulos, V.; Gavish, M. Channel-like functions of the 18-kDa translocator protein (TSPO): Regulation of apoptosis and steroidogenesis as part of the host-defense response. Curr. Pharm. Des. 2007, 13, 2385–2405. [Google Scholar] [CrossRef] [PubMed]
- McEnery, M.W.; Snowman, A.M.; Trifiletti, R.R.; Snyder, S.H. Isolation of the mitochondrial benzodiazepine receptor: Association with the voltage-dependent anion channel and the adenine nucleotide carrier. Proc. Natl. Acad. Sci. USA 1992, 89, 3170–3174. [Google Scholar] [CrossRef] [PubMed]
- Caballero, B.; Veenman, L.; Gavish, M. Role of mitochondrial translocator protein (18 kDa) on mitochondrial-related cell death processes. Recent Pat. Endocr. Metab. Immune Drug Discov. 2013, 7, 86–101. [Google Scholar] [CrossRef] [PubMed]
- Veenman, L.; Shandalov, Y.; Gavish, M. VDAC activation by the 18 kDa translocator protein (TSPO), implications for apoptosis. J. Bioenerg. Biomembr. 2008, 40, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Zeno, S.; Veenman, L.; Katz, Y.; Bode, J.; Gavish, M.; Zaaroor, M. The 18 kDa mitochondrial translocator protein (TSPO) prevents accumulation of protoporphyrin IX. Involvement of reactive oxygen species (ROS). Curr. Mol. Med. 2012, 12, 494–501. [Google Scholar] [CrossRef] [PubMed]
- Veenman, L.; Vainshtein, A.; Yasin, N.; Azrad, M.; Gavish, M. Tetrapyrroles as endogenous TSPO ligands in eukaryotes and prokaryotes: Comparisons with synthetic ligands. Int. J. Mol. Sci. 2016, 17, 880. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Kazumura, K.; Maruyama, W.; Osawa, T.; Naoi, M. Rasagiline and selegiline suppress calcium efflux from mitochondria by PK11195-induced opening of mitochondrial permeability transition pore: A novel anti-apoptotic function for neuroprotection. J. Neural Transm. 2015, 122, 1399–1407. [Google Scholar] [CrossRef] [PubMed]
- Azarashvili, T.; Grachev, D.; Krestinina, O.; Evtodienko, Y.; Yurkov, I.; Papadopoulos, V.; Reiser, G. The peripheral-type benzodiazepine receptor is involved in control of Ca2+-induced permeability transition pore opening in rat brain mitochondria. Cell Calcium 2007, 42, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, N.; Rosenberg, O.; Weizman, A.; Veenman, L.; Gavish, M. In vitro effect of FGIN-1–27, a ligand to 18 kDa mitochondrial translocator protein, in human osteoblast-like cells. J. Bioenerg. Biomembr. 2014, 46, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Balsemão-Pires, E.; Jaillais, Y.; Olson, B.J.; Andrade, L.R.; Umen, J.G.; Chory, J.; Sachetto-Martins, G. The Arabidopsis translocator protein (AtTSPO) is regulated at multiple levels in response to salt stress and perturbations in tetrapyrrole metabolism. BMC Plant Biol. 2011, 11, 108. [Google Scholar] [CrossRef] [PubMed]
- Vanhee, C.; Zapotoczny, G.; Masquelier, D.; Ghislain, M.; Batoko, H. The Arabidopsis multistress regulator TSPO is a heme binding membrane protein and a potential scavenger of porphyrins via an autophagy-dependent degradation mechanism. Plant Cell 2011, 23, 785–805. [Google Scholar] [CrossRef] [PubMed]
- Veenman, L.; Bode, J.; Gaitner, M.; Caballero, B.; Pe’er, Y.; Zeno, S.; Kietz, S.; Kugler, W.; Lakomek, M.; Gavish, M. Effects of 18-kDa translocator protein knockdown on gene expression of glutamate receptors, transporters, and metabolism, and on cell viability affected by glutamate. Pharmacogenet. Genom. 2012, 22, 606–619. [Google Scholar] [CrossRef] [PubMed]
- Vainshtein, A.; Veenman, L.; Shterenberg, A.; Singh, S.; Masarwa, A.; Dutta, B.; Island, B.; Tsoglin, E.; Levin, E.; Leschiner, S.; et al. Quinazoline based tricyclic compounds that regulate programmed cell death, induce neuronal differentiation, and are curative in animal models for excitotoxicity and hereditary brain disease. Cell Death Discov. 2015, 1, 15027. [Google Scholar] [CrossRef] [PubMed]
- Karlstetter, M.; Nothdurfter, C.; Aslanidis, A.; Moeller, K.; Horn, F.; Scholz, R.; Neumann, H.; Weber, B.H.; Rupprecht, R.; Langmann, T. Translocator protein (18 kDa) (TSPO) is expressed in reactive retinal microglia and modulates microglial inflammation and phagocytosis. J. Neuroinflamm. 2014, 11, 3. [Google Scholar] [CrossRef] [PubMed]
- Veenman, L.; Vainsthein, A.; Gavish, M. TSPO as a target for treatments of diseases, including neuropathological disorders. Cell Death Disease 2015, 6, e1911. [Google Scholar] [CrossRef] [PubMed]
- Maaser, K.; Sutter, A.P.; Krahn, A.; Höpfner, M.; Grabowski, P.; Scherübl, H. Cell cycle-related signaling pathways modulated by peripheral benzodiazepine receptor ligands in colorectal cancer cells. Biochem. Biophys. Res. Commun. 2004, 324, 878–886. [Google Scholar] [CrossRef] [PubMed]
- Bode, J.; Veenman, L.; Vainshtein, A.; Kugler, W.; Rosenberg, N.; Gavish, M. Modulation of gene expression associated with the cell cycle and tumorigenicity of glioblastoma cells by the 18 kDa Translocator Protein (TSPO). Austin J. Pharmacol. Ther. 2014, 2, 1053. [Google Scholar]
- Yeliseev, A.A.; Kaplan, S. A sensory transducer homologous to the mammalian peripheral-type benzodiazepine receptor regulates photosynthetic membrane complex formation in Rhodobacter sphaeroides 2.4.1. J. Biol. Chem. 1995, 270, 21167–21175. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.J.; Middleton, R.J.; Kam, W.W.; Chin, D.Y.; Hatty, C.R.; Chan, R.H.; Banati, R.B. Functional gains in energy and cell metabolism after TSPO gene insertion. Cell Cycle 2017, 19. [Google Scholar] [CrossRef] [PubMed]
- Parikh, V.S.; Morgan, M.M.; Scott, R.; Clements, L.S.; Butow, R.A. The mitochondrial genotype can influence nuclear gene expression in yeast. Science 1987, 235, 576–580. [Google Scholar] [CrossRef] [PubMed]
- Ždralević, M.; Guaragnella, N.; Giannattasio, S. Yeast as a tool to study mitochondrial retrograde pathway en route to cell stress response. Methods Mol. Biol. 2015, 1265, 321–331. [Google Scholar] [PubMed]
- Cagin, U.; Enriquez, J.A. The complex crosstalk between mitochondria and the nucleus: What goes in between? Int. J. Biochem. Cell Biol. 2015, 63, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Butow, R.A.; Avadhani, N.G. Mitochondrial signaling: The retrograde response. Mol Cell 2004, 14, 1–15. [Google Scholar] [CrossRef]
- Biswas, G.; Srinivasan, S.; Anandatheerthavarada, H.K.; Avadhani, N.G. Dioxin-mediated tumor progression through activation of mitochondria-to-nucleus stress signaling. Proc. Natl. Acad. Sci. USA 2008, 105, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.P.; Price, N.L.; Ling, A.J.; Moslehi, J.J.; Montgomery, M.K.; Rajman, L.; White, J.P.; Teodoro, J.S.; Wrann, C.D.; Hubbard, B.P.; et al. Declining NAD+ induces a pseudohypoxic state disrupting nuclear-mitochondrial communication during aging. Cell 2013, 155, 1624–1638. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.Y.; Sheng, Z.H. Regulation of mitochondrial transport in neurons. Exp. Cell Res. 2015, 334, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Yi, M.; Weaver, D.; Hajnóczky, G. Control of mitochondrial motility and distribution by the calcium signal: A homeostatic circuit. J. Cell Biol. 2004, 167, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Shargorodsky, L.; Veenman, L.; Caballero, B.; Pe’er, Y.; Leschiner, S.; Bode, J.; Gavish, M. The nitric oxide donor sodium nitroprusside requires the 18 kDa translocator protein to induce cell death. Apoptosis 2012, 17, 647–665. [Google Scholar] [CrossRef] [PubMed]
- Veenman, L.; Alten, J.; Linnemannstöns, K.; Shandalov, Y.; Zeno, S.; Lakomek, M.; Gavish, M.; Kugler, W. Potential involvement of F0F1-ATP(synth)ase and reactive oxygen species in apoptosis induction by the antineoplastic agent erucylphosphohomocholine in glioblastoma cell line: A mechanism for induction of apoptosis via the 18 kDa mitochondrial translocator protein. Apoptosis 2010, 15, 753–768. [Google Scholar] [PubMed]
- Veenman, L.; Gavish, M.; Kugler, W. Apoptosis induction by erucylphosphohomocholine via the 18 kDa mitochondrial translocator protein: Implications for cancer treatment. Anticancer Agents Med. Chem. 2014, 14, 559–577. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.H.; Choi, H.B.; Kim, S.U.; McLarnon, J.G. Mitochondrial ligand inhibits store-operated calcium influx and COX-2 production in human microglia. J. Neurosci. Res. 2006, 83, 1293–1298. [Google Scholar] [CrossRef] [PubMed]
- Azarashvili, T.; Krestinina, O.; Baburina, Y.; Odinokova, I.; Grachev, D.; Papadopoulos, V.; Akatov, V.; Lemasters, J.J.; Reiser, G. Combined effect of G3139 and TSPO ligands on Ca2+-induced permeability transition in rat brain mitochondria. Arch. Biochem. Biophys. 2015, 587, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Costa, B.; da Pozzo, E.; Giacomelli, C.; Taliani, S.; Bendinelli, S.; Barresi, E.; da Settimo, F.; Martini, C. TSPO ligand residence time influences human glioblastoma multiforme cell death/life balance. Apoptosis 2015, 20, 383–398. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Liang, D.; Zhang, H.; Liu, Y.; Li, F.; Chen, Y.H. 4′-Chlorodiazepam, a translocator protein (18 kDa) antagonist, improves cardiac functional recovery during postischemia reperfusion in rats. Exp. Biol. Med. 2010, 235, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Lehtonen, M.T.; Akita, M.; Frank, W.; Reski, R.; Valkonen, J.P. Involvement of a class III peroxidase and the mitochondrial protein TSPO in oxidative burst upon treatment of moss plants with a fungal elicitor. Mol. Plant Microbe Interact. 2012, 25, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Guilarte, T.R.; Loth, M.K.; Guariglia, S.R. TSPO finds NOX2 in microglia for redox homeostasis. Trends Pharmacol. Sci. 2016. [Google Scholar] [CrossRef] [PubMed]
- Zeno, S.; Zaaroor, M.; Leschiner, S.; Veenman, L.; Gavish, M. CoCl2 induces apoptosis via the 18 kDa translocator protein in U118MG human glioblastoma cells. Biochemistry 2009, 48, 4652–4661. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Pae, A.N. Translocator protein (TSPO) ligands for the diagnosis or treatment of neurodegenerative diseases: A patent review (2010–2015; part 1). Expert Opin. Ther. Pat. 2016, 26, 1325–1351. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Das, S.K. Translocator protein (TSPO) in breast cancer. Curr. Mol. Med. 2012, 12, 443–445. [Google Scholar] [CrossRef] [PubMed]
- Ruksha, T.; Aksenenko, M.; Papadopoulos, V. Role of translocator protein in melanoma growth and progression. Arch. Dermatol. Res. 2012, 304, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Bonsack, F., 4th; Alleyne, C.H., Jr.; Sukumari-Ramesh, S. Augmented expression of TSPO after intracerebral hemorrhage: A role in inflammation? J. Neuroinflamm. 2016, 13, 151. [Google Scholar] [CrossRef] [PubMed]
- Randles, M.; Lamb, D.E.; Odat, E.; Taleb-Bendiab, A. Distributed redundancy and robustness in complex systems. J. Comput. Syst. Sci. 2011, 77, 293–304. [Google Scholar] [CrossRef]
- Macia, J.; Solé, R.V. Distributed robustness in cellular networks: Insights from synthetic evolved circuits. J. R. Soc. Interface 2009, 6, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Feala, J.D.; Cortes, J.; Duxbury, P.M.; McCulloch, A.D.; Piermarocchi, C.; Paternostro, G. Statistical properties and robustness of biological controller-target networks. PLoS ONE 2012, 7, e29374. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, N.; Rosenberg, O.; Weizman, A.; Leschiner, S.; Sakoury, Y.; Fares, F.; Soudry, M.; Weisinger, G.; Veenman, L.; Gavish, M. In vitro mitochondrial effects of PK 11195, a synthetic translocator protein 18 kDa (TSPO) ligand, in human osteoblast-like cells. J. Bioenerg. Biomembr. 2011, 43, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Lagutina, I.S.; Mezina, M.N.; Prokof’ev, M.I.; Zakharchenko, V.I.; Galat, V.I. A study of factors affecting the efficiency of electrofusion of enucleated eggs and cell-donors of nuclei. Ontogenez 2002, 33, 100–116. [Google Scholar] [PubMed]
- Tullai, J.W.; Schaffer, M.E.; Mullenbrock, S.; Sholder, G.; Kasif, S.; Cooper, G.M. Immediate-early and delayed primary response genes are distinct in function and genomic architecture. J. Biol. Chem. 2007, 282, 23981–23995. [Google Scholar] [CrossRef] [PubMed]
- Jayakumar, A.R.; Panickar, K.S.; Norenberg, M.D. Effects on free radical generation by ligands of the peripheral benzodiazepine receptor in cultured neural cells. J. Neurochem. 2002, 83, 1226–1234. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, Y.; Guan, M.X. A peep into mitochondrial disorder: Multifaceted from mitochondrial DNA mutations to nuclear gene modulation. Protein Cell 2015, 6, 862–870. [Google Scholar] [CrossRef] [PubMed]
- Arnould, T.; Vankoningsloo, S.; Renard, P.; Houbion, A.; Ninane, N.; Demazy, C.; Remacle, J.; Raes, M. CREB activation induced by mitochondrial dysfunction is a new signaling pathway that impairs cell proliferation. EMBO J. 2002, 21, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Ifuku, M.; Noda, M.; Guilarte, T.R. Translocator protein (18 kDa)/peripheral benzodiazepine receptor specific ligands induce microglia functions consistent with an activated state. Glia 2011, 59, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Levin, E.; Premkumar, A.; Veenman, L.; Kugler, W.; Leschiner, S.; Spanier, I.; Weisinger, G.; Lakomek, M.; Weizman, A.; Snyder, S.H.; et al. The peripheral typebenzodiazepine receptor and tumorigenicity: Isoquinoline binding protein (IBP) antisense knockdown in the C6 glioma cell line. Biochemistry 2005, 44, 9924–9935. [Google Scholar] [CrossRef] [PubMed]
- Bode, J.; Veenman, L.; Caballero, B.; Lakomek, M.; Kugler, W.; Gavish, M. The 18 kDa translocator protein influences angiogenesis, as well as aggressiveness, adhesion, migration, and proliferation of glioblastoma cells. Pharmacogenet. Genom. 2012, 22, 538–550. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Kalyanasundaram, A.; Zhu, J. Structural and biomechanical basis of mitochondrial movement in eukaryotic cells. Int. J. Nanomed. 2013, 8, 4033–4042. [Google Scholar]
- Zhang, Z.W.; Zhang, G.C.; Zhu, F.; Zhang, D.W.; Yuan, S. The roles of tetrapyrroles in plastid retrograde signaling and tolerance to environmental stresses. Planta 2015, 242, 1263–1276. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhai, K.; Xue, Y.; Yang, J.; Yang, Q.; Fu, Y.; Hu, Y.; Liu, F.; Wang, W.; Cui, L.; et al. Global deletion of TSPO does not affect the viability and gene expression profile. PLoS ONE 2016, 11, e0167307. [Google Scholar] [CrossRef] [PubMed]
- Middleton, R.J.; Liu, G.J.; Banati, R.B. Guwiyang Wurra—“Fire Mouse”: A global gene knockout model for TSPO/PBR drug development, loss-of-function and mechanisms of compensation studies. Biochem. Soc. Trans. 2015, 43, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Gut, P.; Zweckstetter, M.; Banati, R.B. Lost in translocation: The functions of the 18-kD translocator protein. Trends Endocrinol. Metab. 2015, 26, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Midzak, A.; Zirkin, B.; Papadopoulos, V. Translocator protein: Pharmacology and steroidogenesis. Biochem. Soc. Trans. 2015, 43, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Liu, J.; Liu, N.; Kuhn, L.A.; Garavito, R.M.; Ferguson-Miller, S. Translocator protein 18 kDa (TSPO): An old protein with new functions? Biochemistry 2016, 55, 2821–2831. [Google Scholar] [CrossRef] [PubMed]
- Gatliff, J.; Campanella, M. TSPO: Kaleidoscopic 18-kDa amid biochemical pharmacology, control and targeting of mitochondria. Biochem. J. 2016, 473, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, V.; Tu, L.N. Current status and future perspectives: TSPO in steroid neuroendocrinology. J. Endocrinol. 2016, 231, R1–R30. [Google Scholar] [CrossRef] [PubMed]
- Batoko, H.; Veljanovski, V.; Jurkiewicz, P. Enigmatic Translocator protein (TSPO) and cellular stress regulation. Trends Biochem. Sci. 2015, 40, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Da Pozzo, E.; Giacomelli, C.; Barresi, E.; Costa, B.; Taliani, S.; Passetti Fda, S.; Martini, C. Targeting the 18-kDa translocator protein: Recent perspectives for neuroprotection. Biochem. Soc. Trans. 2015, 43, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Issop, L.; Ostuni, M.A.; Lee, S.; Laforge, M.; Péranzi, G.; Rustin, P.; Benoist, J.F.; Estaquier, J.; Papadopoulos, V.; Lacapère, J.J. Translocator protein-mediated stabilization of mitochondrial architecture during inflammation stress in colonic cells. PLoS ONE 2016, 11, e0152919. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Pae, A.N. Translocator protein (TSPO) ligands for the diagnosis or treatment of neurodegenerative diseases: A patent review (2010–2015; part 2). Expert Opin. Ther. Pat. 2016, 26, 1353–1366. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar]
- Krämer, A.; Green, J.; Pollard, J.; Tugendreich, S. Causal analysis approaches in ingenuity pathway analysis. Bioinform. 2014, 30, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Caballero, B.; Veenman, L.; Bode, J.; Leschiner, S.; Gavish, M. Concentration-dependent bimodal effect of specific 18 kDa translocator protein (TSPO) ligands on cell death processes induced by ammonium chloride: Potential implications for neuropathological effects due to hyperammonemia. CNS Neurol. Disord. Drug Targets 2014, 13, 574–592. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 15–45 min of PK 11195 Exposure (25 µM) | |||||
| 15 min Up-regulated genes ↑ | 30 min Up-regulated genes ↑ | 45 min Up-regulated genes ↑ | |||
| FOS | ↑ +2.16 | FOS | ↑ +3.96 | EGR1 | ↑ +2.45 |
| DUSP1 | ↑ +2.08 | DUSP1 | ↑ +3.64 | PTGS2 | ↑ +2.00 |
| EGR1 | ↑ +1.89 | EGR1 | ↑ +2.64 | FOS | ↑ +1.84 |
| CYR61 | ↑ +1.82 | CYR61 | ↑ +2.16 | DUSP1 | ↑ +1.77 |
| OSR1 | ↑ +1.71 | MYC | ↑ +2.03 | FOSB | ↑ +1.72 |
| ANP32AP1 | ↑ +1.64 | CXCL8 | ↑ +2.02 | CYR61 | ↑ +1.69 |
| FKBP10 | ↑ +1.54 | PTGS2 | ↑ +2.01 | SGK1 | ↑ +1.67 |
| RNA28S5 | ↑ +1.54 | CTGF | ↑ +1.93 | CXCL8 | ↑ +1.62 |
| DYNC1H1 | ↑ +1.52 | NFKBIZ | ↑ +1.90 | ATF3 | ↑ +1.58 |
| WNK1 | ↑ +1.51 | SGK1 | ↑ +1.89 | ANP32AP1 | ↑ +1.58 |
| 15 min Down-regulated genes ↓ | 30 min Down-regulated genes ↓ | 45 min Down-regulated genes ↓ | |||
| ID3 | ↓ −1.83 | LOC100507412 | ↓ −1.72 | MYLIP | ↓ −1.84 |
| TUFT1 | ↓ −1.83 | TUFT1 | ↓ −1.51 | LOC441087 | ↓ −1.50 |
| ID2 | ↓ −1.68 | MYLIP | ↓ −1.50 | ||
| PTMA | ↓ −1.60 | KDM3A | ↓ −1.50 | ||
| ID1 | ↓ −1.56 | ||||
| KDM3A | ↓ −1.55 | ||||
| NABP1 | ↓ −1.52 | ||||
| RPL21P28 | ↓ −1.50 | ||||
| CLK1 | ↓ −1.50 | ||||
| 1–24 h of PK 11195 Exposure (25 µM) | |||||
| 1 h Up-regulated genes ↑ | 3 h Up-regulated genes ↑ | 24 h Up-regulated genes ↑ | |||
| CYR61 | ↑ +9.3 | ID3 | ↑ +3.38 | ASN | ↑ +2.47 |
| FOSB | ↑ +4.29 | ID1 | ↑ +2.08 | SLC7A5 | ↑ +2.39 |
| EGR2 | ↑ +3.32 | GBP1 | ↑ +2.04 | TRIB3 | ↑ +2.21 |
| EGR1 | ↑ +3.2 | SMAD6 | ↑ +1.89 | PCK2 | ↑ +2.18 |
| CTGF | ↑ +3.04 | ID2 | ↑ +1.86 | LOC729779 | ↑ +2.12 |
| ID3 | ↑ +2.62 | ATOH8 | ↑ +1.85 | PSAT1 | ↑ +1.97 |
| TUFT1 | ↑ +2.49 | TXNIP | ↑ +1.84 | NUPR1 | ↑ +1.94 |
| ID1 | ↑ +2.33 | NEXN | ↑ +1.77 | P8 | ↑ +1.88 |
| SRF | ↑ +2.15 | SLC3A2 | ↑ +1.74 | DDIT4 | ↑ +1.84 |
| GBP1 | ↑ +2.12 | ACTG2 | ↑ +1.71 | FAM102A | ↑ +1.82 |
| PTGS2 | ↑ +2.11 | FHL2 | ↑ +1.65 | SLC1A5 | ↑ +1.8 |
| TRIB1 | ↑ +2.09 | DDIT3 | ↑ +1.8 | ||
| ERRFI1 | ↑ +2.04 | ATF4 | ↑ +1.77 | ||
| ATF3 | ↑ +1.95 | TGIF1 | ↑ +1.75 | ||
| KLF6 | ↑ +1.74 | SPRR2D | ↑ +1.72 | ||
| FOS | ↑ +1.73 | PHGDH | ↑ +1.71 | ||
| DUSP5 | ↑ +1.71 | SLC3A2 | ↑ +1.69 | ||
| PTGER4 | ↑ +1.69 | PLEKHF1 | ↑ +1.67 | ||
| SGK | ↑ +1.69 | FOLR3 | ↑ +1.65 | ||
| GADD45A | ↑ +1.69 | BEX2 | ↑ +1.63 | ||
| SGK1 | ↑ +1.69 | SLC6A15 | ↑ +1.63 | ||
| ID2 | ↑ +1.68 | IGFBP1 | ↑ +1.56 | ||
| FILIP1L | ↑ +1.67 | ||||
| 1 h Down-regulated genes ↓ | 3 h Down-regulated genes ↓ | 24 h Down-regulated genes ↓ | |||
| BCL6 | ↓ −2.14 | IL8 | ↓ −2.47 | MYLIP | ↓ −2.24 |
| DDIT4 | ↓ −1.72 | MYLIP | ↓ −2.36 | UHRF1 | ↓ −1.80 |
| SOX4 | ↓ −1.89 | IGFBP5 | ↓ −1.79 | ||
| RGS4 | ↓ −1.72 | ||||
| PDE5A | ↓ −1.69 | ||||
| TYMS | ↓ −1.65 | ||||
| ERRFI1 | ↓ −1.63 | ||||
| Gene Expression Changes in U118MG Glioblastoma Cells after PK 11195 Exposure for different Time Periods | |||||
|---|---|---|---|---|---|
| 15 min | 30 min | 45 min | 60 min | 3 h | 24 h |
| All Types of Genes Combined | |||||
| 20 Genes | 14 Genes | 12 Genes | 25 Genes | 14 Genes | 29 Genes |
| Transcription Factors | |||||
| ATF3 * | |||||
| BCL6 * | |||||
| DUSP5 | |||||
| CLK1 | EGR1 * | ||||
| DUSP1 * | EGR2 * | ||||
| EGR1 * | FOS * | ||||
| FOS * | FOSB * | ||||
| OSR1 * | GADD45A * | ||||
| ID1 * | DUSP1 * | ID1 * | ATOH8 | ATF4 * | |
| ID2 * | EGR1 * | ATF3 * | ID2 * | ID1 * | DDIT3 * |
| ID3 * | FOS * | DUSP1 * | ID3 * | ID2 * | NUPR1 * |
| KDM3A * | KDM3A | EGR1 * | KLF6 * | ID3 * | TGIF1 |
| NABP1 | MYC * | FOS * | SRF | SOX4 | TRIB3 * |
| PTMA * | NFKBIZ * | FOSB * | TRIB1 * | SMAD6 * | UHRF1 |
| (11 genes) | (6 genes) | (5 genes) | (14 genes) | (6 genes) | (6 genes) |
| Proteins, Enzymes, and other Products | |||||
| ASNS * | |||||
| BEX2 * | |||||
| DDIT4 * | |||||
| ERRFI1 | |||||
| FAM102A | |||||
| FOLR3 | |||||
| IGFBP1 * | |||||
| IGFBP5 * | |||||
| LOC729779 | |||||
| MYLIP | |||||
| PCK2 | |||||
| PDE5A | |||||
| CTGF * | PHGDH | ||||
| CYR61 * | PLEKHF1 | ||||
| ANP32AP1 | DDIT4 * | P8 | |||
| CYR61 * | CTGF * | ERRFI1 | ACTG | PSAT1 | |
| DYNC1H1 * | CXCL8 | ANP32AP1 | GBP1 | FHL2 * | RGS4 |
| FKBP10 | CYR61 * | CXCL8 | FILIP1L | GBP1 | SLC1A5 |
| MIR22HG | LOC100507412 | CYR61 * | PTGER4 | IL8 | SLC3A2 |
| RNA28S5 | MYLIP | LOC441087 | PTGS2 * | MYLIP | SLC6A15 |
| RPL21P28 | PTGS2 * | MYLIP | SGK | NEXN | SLC7A5 |
| TUFT1 | SGK1 * | PTGS2 * | SGK1 * | SLC3A2 | SPRR2D |
| WNK1 | TUFT1 | SGK1 * | TUFT1 | TXNIP * | TYMS |
| (9 genes) | (8 genes) | (7 genes) | (11 genes) | (8 genes) | (23 genes) |
| Ct | Vehicle | PK 11195 |
|---|---|---|
| B2M | 22.10 ± 0.85 | 21.85 ± 0.21 n.s. |
| FOS | 29.35 ± 0.07 | 26.20 ± 0.28 *** |
| DUSP1 | 24.90 ± 0.42 | 22.85 ± 0.21 ** |
| 15 min | 30 min | 45 min | 1 h | 3 h | 24 h |
|---|---|---|---|---|---|
| Gene expression modulation
Binding of protein binding site Transactivation of RNA Endothelial cell development Cell viability Accumulation of cells | Binding of protein binding site Synthesis of DNA Differentiation of connective tissue cells Development of neurons Formation of cells Microtubule dynamics Chemotaxis of cells Cell movement of fibroblast cell lines Metastasis of tumor cell lines Abdominal neoplasm Proliferation of lymphocytes Growth of tumor Cell viability Metabolism of carbohydrate Inflammation of body region | Synthesis of DNA Development of neurons Formation of cellular protrusions Angiogenesis Proliferation Migration Cell growth | Apoptosis of fibroblast cell lines Malignant solid tumor S phase Cell cycle progression of fibroblast cell lines Development of cardiovascular system Cell viability Formation of cellular protrusions Growth of malignant tumor Proliferation of tumor cells Formation of cells Development of reproductive system | Cell death of central nervous system cells Apoptosis of fibroblasts Apoptosis of myeloid cells Apoptosis of muscle cell lines Necrosis of epithelial tissue Migration of colon cancer cell lines Cell movement of leukocyte cell lines Migration of smooth muscle cells Migration of phagocytes Chemotaxis Cell viability Development of epithelial tissue Proliferation of leukocyte cell lines Activation of leukocytes Inflammatory response Accumulation of leukocytes Proliferation of leukemia cell lines Activation of tumor cell lines | Cell death of fibroblast cell lines Apoptosis of kidney cell lines Apoptosis of epithelial cell lines Abdominal cancer Digestive system cancer Growth of digestive organ tumor Growth of malignant tumor Epithelial cancer Proliferation of tumor cells |
| 50 µM | 100 µM | ||
|---|---|---|---|
| Up-regulated genes ↑ | Up-regulated genes ↑ | ||
| FOS | ↑ 2.5 | FOS | ↑ 3.46 |
| ZFP36 | ↑ 1.68 | ZFP36 | ↑ 1.72 |
| DUSP1 | ↑ 1.62 | DUSP1 | ↑ 1.71 |
| Down-regulated genes ↓ | Down-regulated genes ↓ | ||
| TUFT1 | ↓ 1.64 | ID2 | ↓ 1.68 |
| Observations | Caveats or Questions | Answer or Future Studies |
|---|---|---|
| PK 11195 applications affect gene expression and the related functions. | Is this context dependent? | The application of PK 11195 can be further refined (dose, time window, co-factors, etc.). |
| TSPO ligands other than PK 11195 affect gene expression and related functions. | Is this context dependent? | If desired, the application of ligands other than PK 11195 can be further refined (dose, time window, co-factors, etc.). |
| TSPO knockdown affects gene expression and related functions. | Is this context dependent? | If desired, the application of TSPO knockdown and knockout can be further refined (transient, stable, time course of effects, co-factors, etc.). |
| Interaction between PK 11195 and TSPO. PK 11195 can interact with TSPO to finally affect gene expression. | PK 11195 can also bind to other receptors. This can also occur via other receptors. | Knockout all the other receptors binding to PK 11195 (Difficult in practice). TSPO can be knocked down to address this question (This has been done). It can be checked whether other TSPO ligands can affect gene expression (This has been done). |
| Interactions between TSPO and PK 11195 affect mitochondrial to nuclear signaling, for example via the initiating steps of ROS regeneration, and Ca2+ and ATP release. | Will different concentrations of ATP and Ca2+ and levels of ROS relate to different changes in gene expression? | ATP and Ca2+ levels and ROS generation can be measured after TSPO manipulations. Gene expression can be measured. Protein expression of calcium sensitive proteins can be measured. |
| Mitochondrial to nuclear signaling apparently induced by TSPO and its ligands implicates various calcium sensitive proteins. | Which calcium sensitive proteins can be activated due to TSPO modulations? | One can measure calcium binding proteins after TSPO manipulations. |
| Immediate early genes and other transcription factors are activated as a consequence of TSPO knockdown and TSPO ligand applications. | TSPO ligands can act via other receptors than TSPO. TSPO knockdown can possibly be compensated by various cellular mechanisms. | One should not rely on one method to induce effects of TSPO manipulation on gene expression. The time interval between measurements after TSPO manipulation, such as knockdown or knockout, or application of TSPO ligands, or other agents affecting TSPO activity should be as short as possible. In this way, compensatory events are precluded. |
| Immediate early genes induce changes in expression of other genes. | How is it ‘decided’ which genes will be modulated? One molecule such as PK 11195 by itself cannot determine which complex of genes will change expression. | TSPO modulation has to be combined with co-factors or different contexts. The assumption is that these additional variations, together with TSPO modulation determine the patterns in changes of gene expression. For example switching from minimal to maximal cell culture medium results in major changes in numbers of gene expression changes. |
| The final changes in gene expression correlate with functional changes. | Does this always occur? | Modulation of TSPO activity, modulation of gene expression, and modulation of function always have to be considered in association with each other and subjected to combined studies (an approach taken in the present study). |
| Modulation of gene expression typically has functional effects. | How important is control of TSPO and its ligands of gene expression? | We think it is very important, at least in cell culture and also in animal models we have seen major phenotypic changes. |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yasin, N.; Veenman, L.; Singh, S.; Azrad, M.; Bode, J.; Vainshtein, A.; Caballero, B.; Marek, I.; Gavish, M. Classical and Novel TSPO Ligands for the Mitochondrial TSPO Can Modulate Nuclear Gene Expression: Implications for Mitochondrial Retrograde Signaling. Int. J. Mol. Sci. 2017, 18, 786. https://doi.org/10.3390/ijms18040786
Yasin N, Veenman L, Singh S, Azrad M, Bode J, Vainshtein A, Caballero B, Marek I, Gavish M. Classical and Novel TSPO Ligands for the Mitochondrial TSPO Can Modulate Nuclear Gene Expression: Implications for Mitochondrial Retrograde Signaling. International Journal of Molecular Sciences. 2017; 18(4):786. https://doi.org/10.3390/ijms18040786
Chicago/Turabian StyleYasin, Nasra, Leo Veenman, Sukhdev Singh, Maya Azrad, Julia Bode, Alex Vainshtein, Beatriz Caballero, Ilan Marek, and Moshe Gavish. 2017. "Classical and Novel TSPO Ligands for the Mitochondrial TSPO Can Modulate Nuclear Gene Expression: Implications for Mitochondrial Retrograde Signaling" International Journal of Molecular Sciences 18, no. 4: 786. https://doi.org/10.3390/ijms18040786
APA StyleYasin, N., Veenman, L., Singh, S., Azrad, M., Bode, J., Vainshtein, A., Caballero, B., Marek, I., & Gavish, M. (2017). Classical and Novel TSPO Ligands for the Mitochondrial TSPO Can Modulate Nuclear Gene Expression: Implications for Mitochondrial Retrograde Signaling. International Journal of Molecular Sciences, 18(4), 786. https://doi.org/10.3390/ijms18040786