
CIK Cells and HDAC Inhibitors in Multiple Myeloma

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
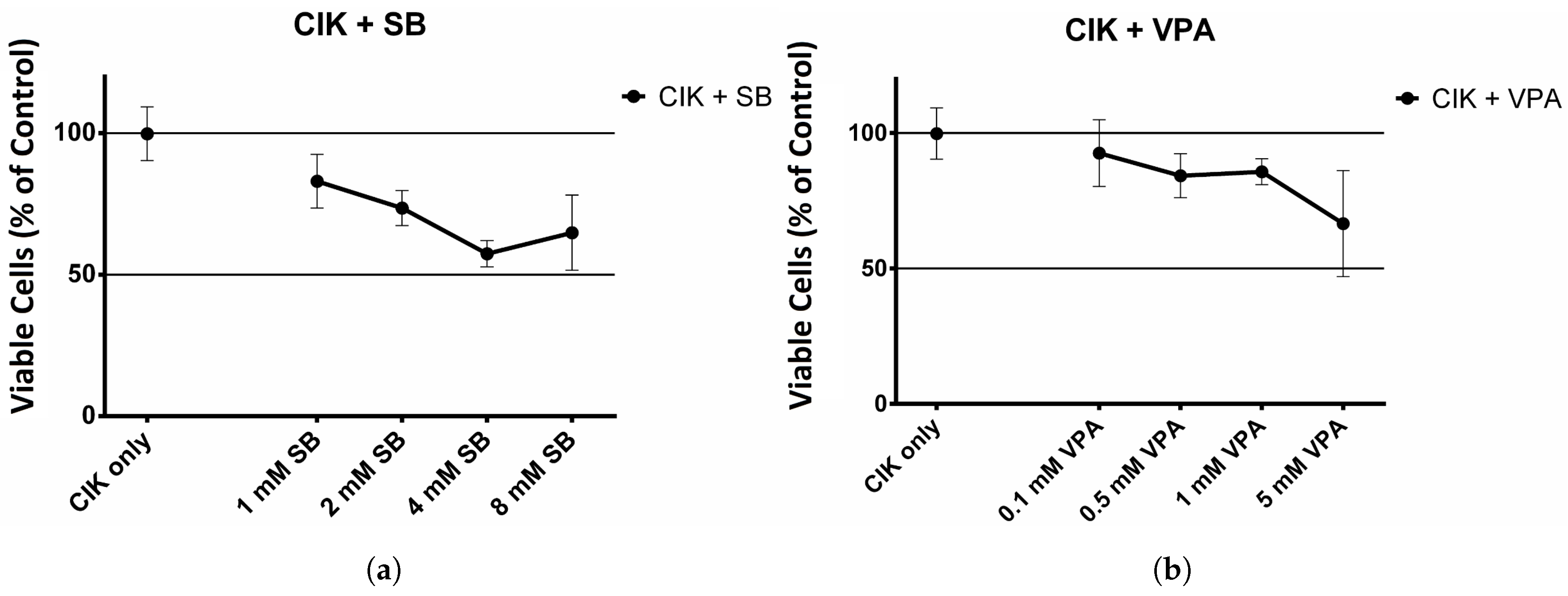
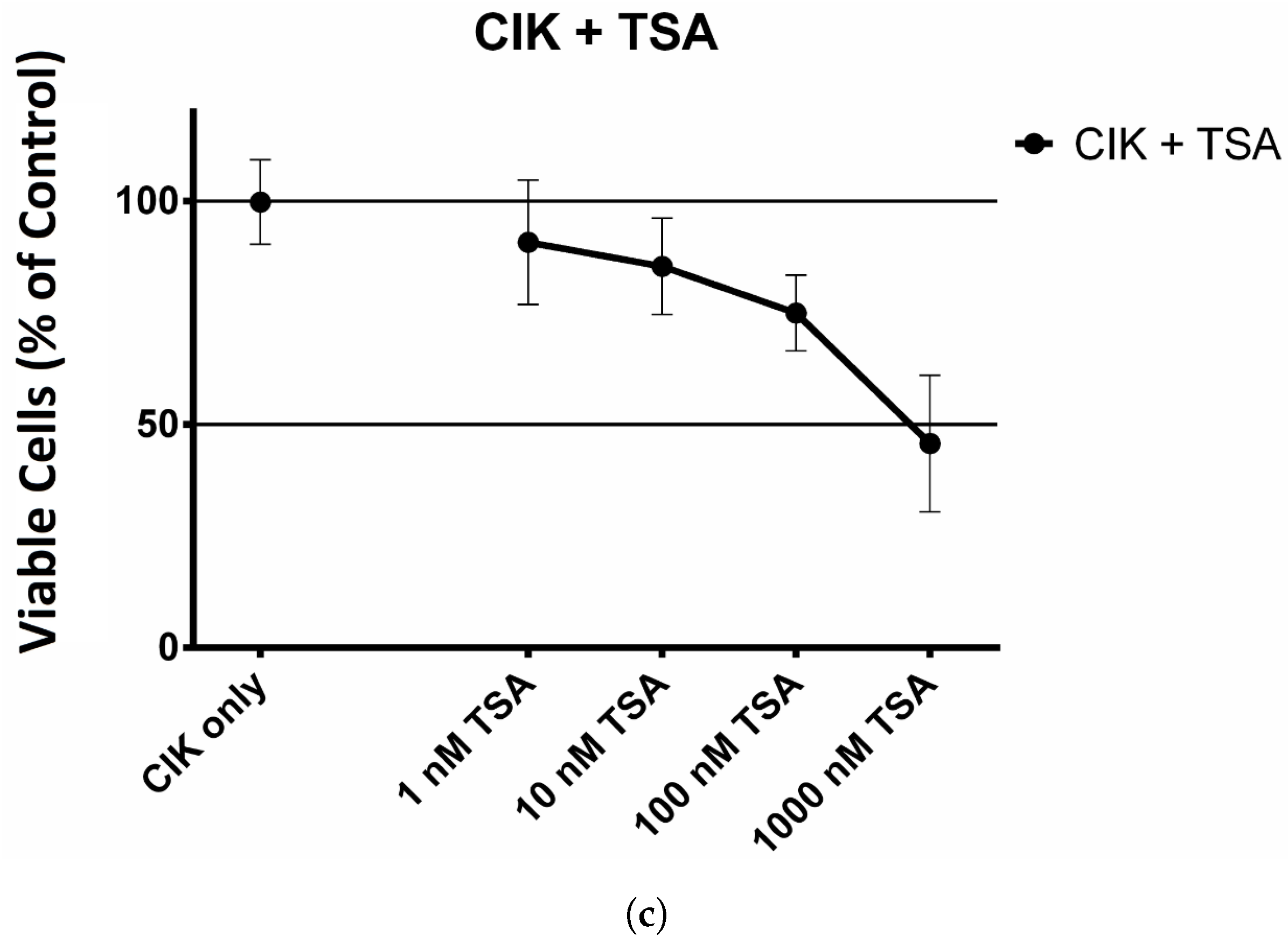
2.1. The Effect of HDAC Inhibitors on Cell Viability of CIK Cells
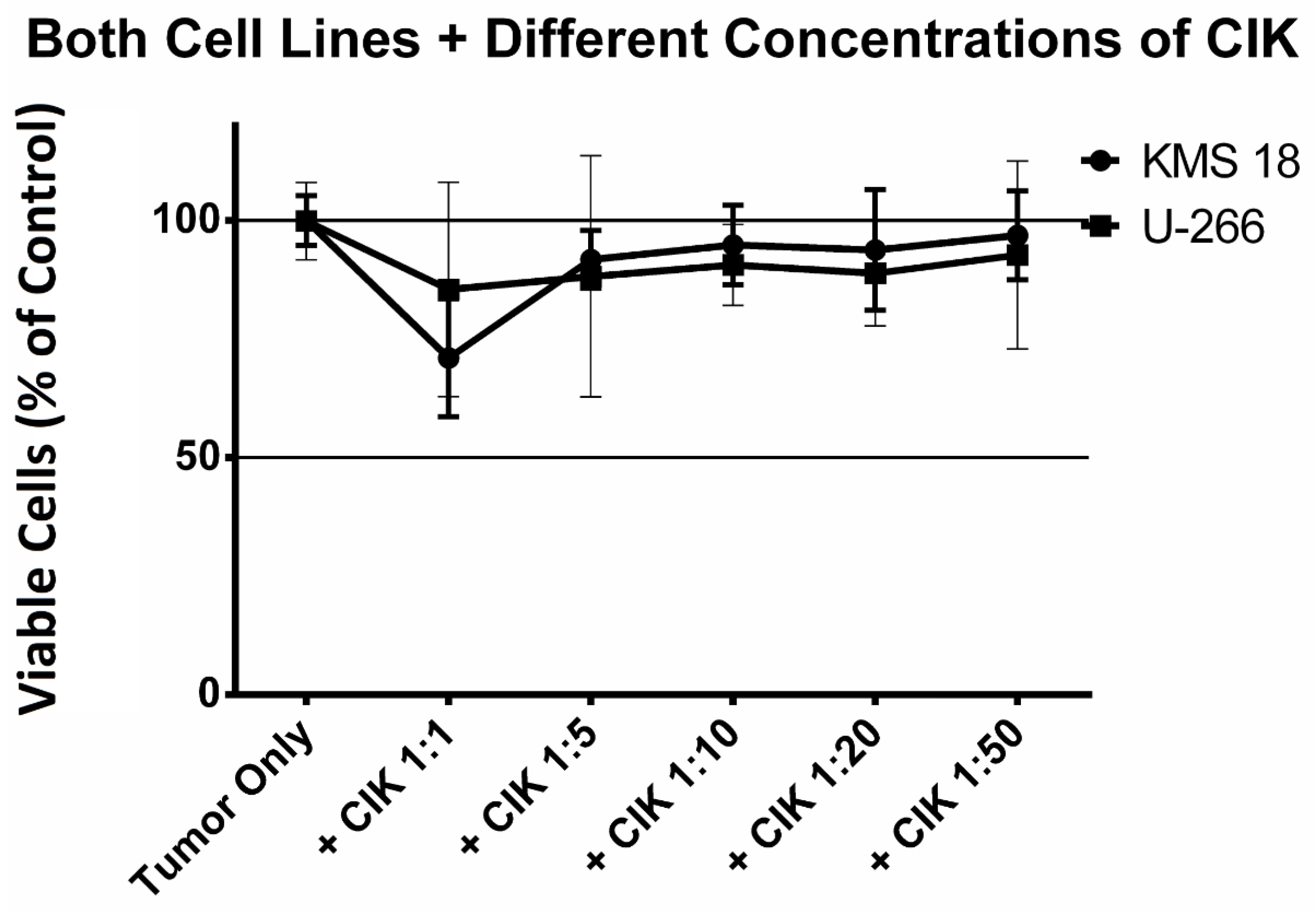
2.2. Finding the Optimum Ratio of CIK Cells to Tumor Cells
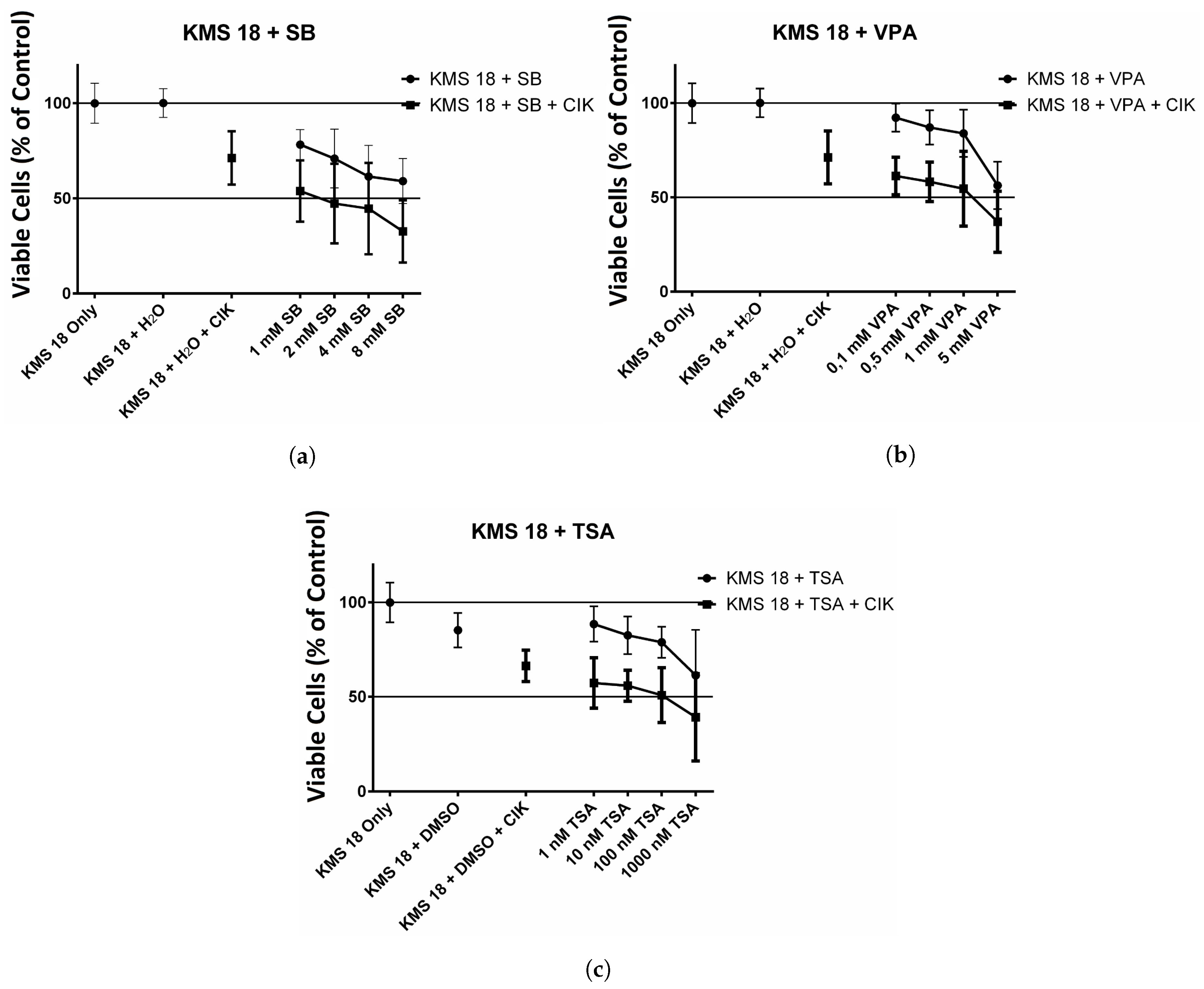
2.3. Effect of HDAC Inhibitors and CIK Cells on KMS 18 cells
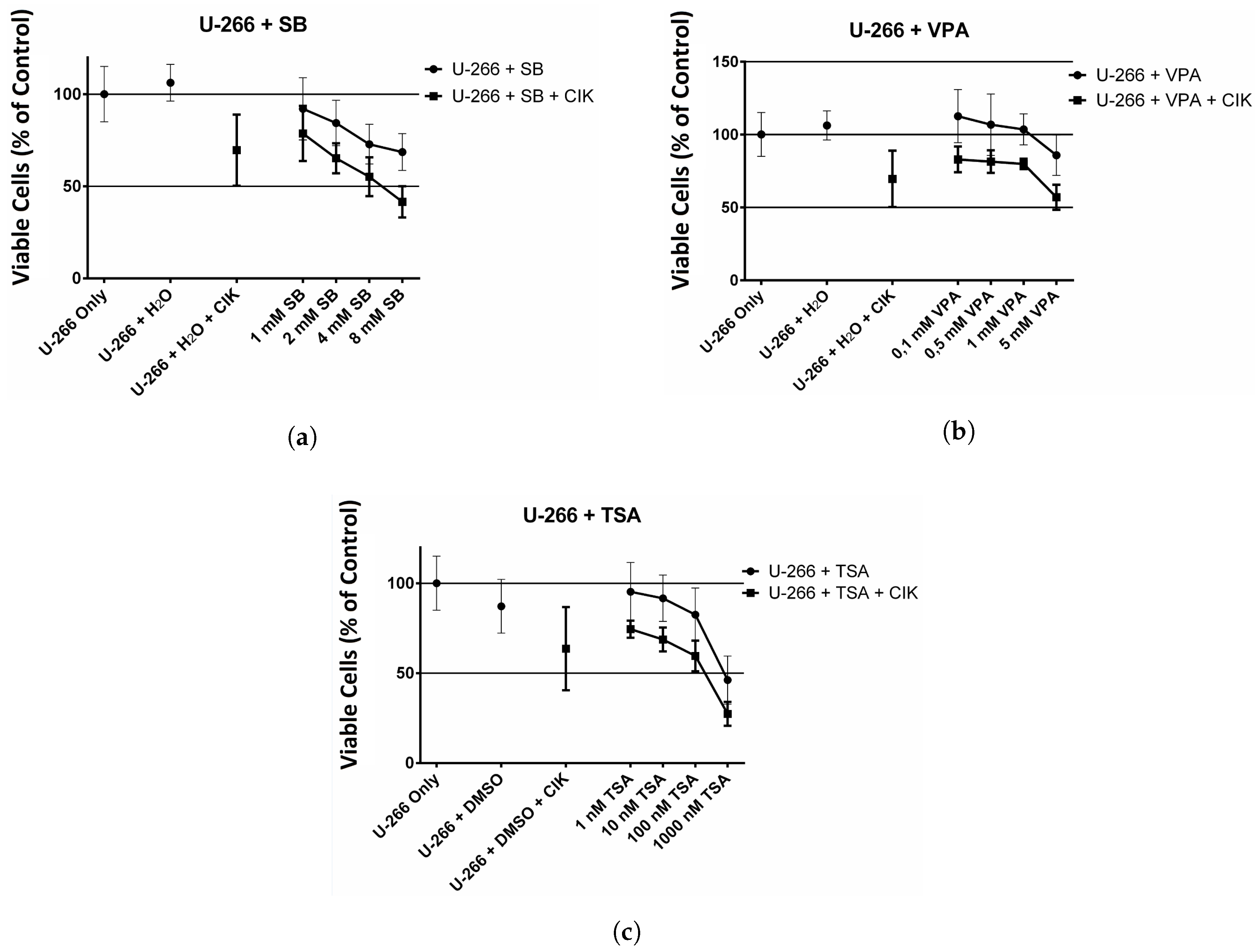
2.4. Effect of HDAC Inhibitors and CIK Cells on U-266 Cells
3. Discussion
4. Materials and Methods
4.1. Cells
4.2. Buffy Coat
- 300 U·mL IL-2
- 50 ng·mL-CD3
- 100 U·mL interleukin1 (IL-1 )
4.3. MTT Assay
Sodium Butyrate:
| Valproic Acid:
| Trichostatin A:
|
4.4. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| MM | Multiple myeloma |
| NK | natural killer |
| CIK | Cytokine-induced killer |
| NKT | natural killer T |
| HDAC | Histone deacetylase |
| SB | Sodium Butyrate |
| VPA | Valproic Acid |
| TSA | Trichostatin A |
| FCS | fetal calf serum |
| P/S | penicillin/streptomycin |
| IL-2 | interleukin-2 |
| PBS | phosphate buffered saline |
| BSA | bovine serum albumin |
| IFN- | interferon gamma |
| IL | interleukin |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
References
- Clark, S.M.; Steinbach, A.; Clemmons, A.B. Pomalidomide for the Treatment of Multiple Myeloma. J. Adv. Pract. Oncol. 2014, 5, 51. [Google Scholar] [PubMed]
- Kröpil, P.; Fenk, R.; Fritz, L.B.; Blondin, D.; Kobbe, G.; Mödder, U.; Cohnen, M. Comparison of whole-body 64-slice multidetector computed tomography and conventional radiography in staging of multiple myeloma. Eur. Radiol. 2008, 18, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Galson, D.L.; Silbermann, R.; Roodman, G.D. Mechanisms of multiple myeloma bone disease. BoneKEy Rep. 2012, 1. [Google Scholar] [CrossRef] [PubMed]
- Romano, A.; Conticello, C.; Cavalli, M.; Vetro, C.; La Fauci, A.; Parrinello, N.L.; Di Raimondo, F. Immunological Dysregulation in Multiple Myeloma Microenvironment. BioMed Res. Int. 2014, 2014, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Soriani, A.; Fionda, C.; Ricci, B.; Iannitto, M.L.; Cippitelli, M.; Santoni, A. Chemotherapy-elicited upregulation of NKG2D and DNAM-1 ligands as a therapeutic target in multiple myeloma. OncoImmunology 2013, 2, e26663. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Wolf, I.; Negrin, R.S.; Kiem, H.P.; Blume, K.G.; Weissman, I.L. Use of a SCID mouse/human lymphoma model to evaluate cytokine-induced killer cells with potent antitumor cell activity. J. Exp. Med. 1991, 174, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Wolf, I.; Lefterova, P.; Mehta, B.; Fernandez, L.; Huhn, D.; Blume, K.; Weissman, I.; Negrin, R. Phenotypic characterization and identification of effector cells involved in tumor cell recognition of cytokine-induced killer cells. Exp. Hematol. 1993, 21, 1673–1679. [Google Scholar] [PubMed]
- Guetgemann, S.; Frank, S.; Strehl, J.; Schmidt-Wolf, I.G. Cytokine-induced killer cells are type II natural killer T cells. GMS German Med.Sci. 2007, 5. [Google Scholar]
- Hontscha, C.; Borck, Y.; Zhou, H.w.; Messmer, D.; Schmidt-Wolf, I. Clinical trials on CIK cells: first report of the international registry on CIK cells (IRCC). J. Cancer Res. Clin. Oncol. 2011, 137, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Mariuzza, R.A. Structural basis for recognition of cellular and viral ligands by NK cell receptors. Front. Immunol. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- McFarland, B.J.; Kortemme, T.; Yu, S.F.; Baker, D.; Strong, R.K. Symmetry recognizing asymmetry: analysis of the interactions between the C-type lectin-like immunoreceptor NKG2D and MHC class I-like ligands. Structure 2003, 11, 411–422. [Google Scholar] [CrossRef]
- Mincheva-Nilsson, L.; Baranov, V. Cancer Exosomes and NKG2D Receptor–Ligand Interactions: Impairing NKG2D-Mediated Cytotoxicity and Anti-Tumour Immune Surveillance; Seminars in Cancer Biology; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Miller, T.A.; Witter, D.J.; Belvedere, S. Histone Deacetylase Inhibitors. J. Med. Chem. 2003, 46, 5097–5116. [Google Scholar] [CrossRef] [PubMed]
- Goetze, S.; Coersmeyer, M.; Mueller, O.; Sievers, S. Histone deacetylase inhibitors induce attenuation of Wnt signaling and TCF7L2 depletion in colorectal carcinoma cells. Int. J. Oncol. 2014, 45, 1715–1723. [Google Scholar]
- Bezecny, P. Histone deacetylase inhibitors in glioblastoma: pre-clinical and clinical experience. Med. Oncol. 2014, 31, 985. [Google Scholar] [CrossRef] [PubMed]
- Laubach, J.P.; Moreau, P.; San-Miguel, J.F.; Richardson, P.G. Panobinostat for the Treatment of Multiple Myeloma. Clin. Cancer Res. 2015, 21, 4767–4773. [Google Scholar] [CrossRef] [PubMed]
- Mai, E.; Goldschmidt, H. Klinik und Therapie des multiplen Myeloms. Radiologe 2014, 54, 538–544. [Google Scholar] [CrossRef] [PubMed]
- West, A.C.; Smyth, M.J.; Johnstone, R.W. The anticancer effects of HDAC inhibitors require the immune system. Oncoimmunology 2014, 3, e27414. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, Z.; Zhou, F.; Sun, Y.; Chen, J.; Li, L.; Jin, H.; Qian, Q. The combination of dendritic cells-cytotoxic T lymphocytes/cytokine-induced killer (DC-CTL/CIK) therapy exerts immune and clinical responses in patients with malignant tumors. Exp. Hematol. Oncol. 2015, 4, 32. [Google Scholar] [CrossRef] [PubMed]
- Mu, Y.; Zhou, C.H.; Chen, S.F.; Ding, J.; Zhang, Y.X.; Yang, Y.P.; Wang, W.H. Effectiveness and safety of chemotherapy combined with cytokine-induced killer cell /dendritic cell–cytokine-induced killer cell therapy for treatment of gastric cancer in China: A systematic review and meta-analysis. Cytotherapy 2016, 18, 1162–1177. [Google Scholar] [CrossRef] [PubMed]
- Nwangwu, C.A.; Weiher, H.; Schmidt-Wolf, I.G.H. Increase of CIK cell efficacy by upregulating cell surface MICA and inhibition of NKG2D ligand shedding in multiple myeloma. Hematol. Oncol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Fu, J.; Zhang, M.; Liu, D. AFM13: a first-in-class tetravalent bispecific anti-CD30/CD16A antibody for NK cell-mediated immunotherapy. J. Hematol. Oncol. 2015, 8, 96. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sanchez, L.; Siegel, D.S.; Wang, M.L. Elotuzumab for the treatment of multiple myeloma. J. Hematol. Oncol. 2016, 9, 55. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, L.; Wang, Y.; Siegel, D.S.; Wang, M.L. Daratumumab: a first-in-class CD38 monoclonal antibody for the treatment of multiple myeloma. J. Hematol. Oncol. 2016, 9, 51. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, A.; Chanan-Khan, A.; Weisel, K.; Nooka, A.K.; Masszi, T.; Beksac, M.; Spicka, I.; Hungria, V.; Munder, M.; Mateos, M.V.; Mark, T.M.; Qi, M.; Schecter, J.; Amin, H.; Qin, X.; Deraedt, W.; Ahmadi, T.; Spencer, A.; Sonneveld, P. Daratumumab, Bortezomib, and Dexamethasone for Multiple Myeloma. N. Engl. J. Med. 2016, 375, 754–766. [Google Scholar] [CrossRef] [PubMed]





© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stephan, D.; Weiher, H.; Schmidt-Wolf, I.G.H. CIK Cells and HDAC Inhibitors in Multiple Myeloma. Int. J. Mol. Sci. 2017, 18, 945. https://doi.org/10.3390/ijms18050945
Stephan D, Weiher H, Schmidt-Wolf IGH. CIK Cells and HDAC Inhibitors in Multiple Myeloma. International Journal of Molecular Sciences. 2017; 18(5):945. https://doi.org/10.3390/ijms18050945
Chicago/Turabian StyleStephan, David, Hans Weiher, and Ingo G.H. Schmidt-Wolf. 2017. "CIK Cells and HDAC Inhibitors in Multiple Myeloma" International Journal of Molecular Sciences 18, no. 5: 945. https://doi.org/10.3390/ijms18050945
APA StyleStephan, D., Weiher, H., & Schmidt-Wolf, I. G. H. (2017). CIK Cells and HDAC Inhibitors in Multiple Myeloma. International Journal of Molecular Sciences, 18(5), 945. https://doi.org/10.3390/ijms18050945