MicroRNA-27b Depletion Enhances Endotrophic and Intravascular Lipid Accumulation and Induces Adipocyte Hyperplasia in Zebrafish


Abstract
:1. Introduction
2. Results
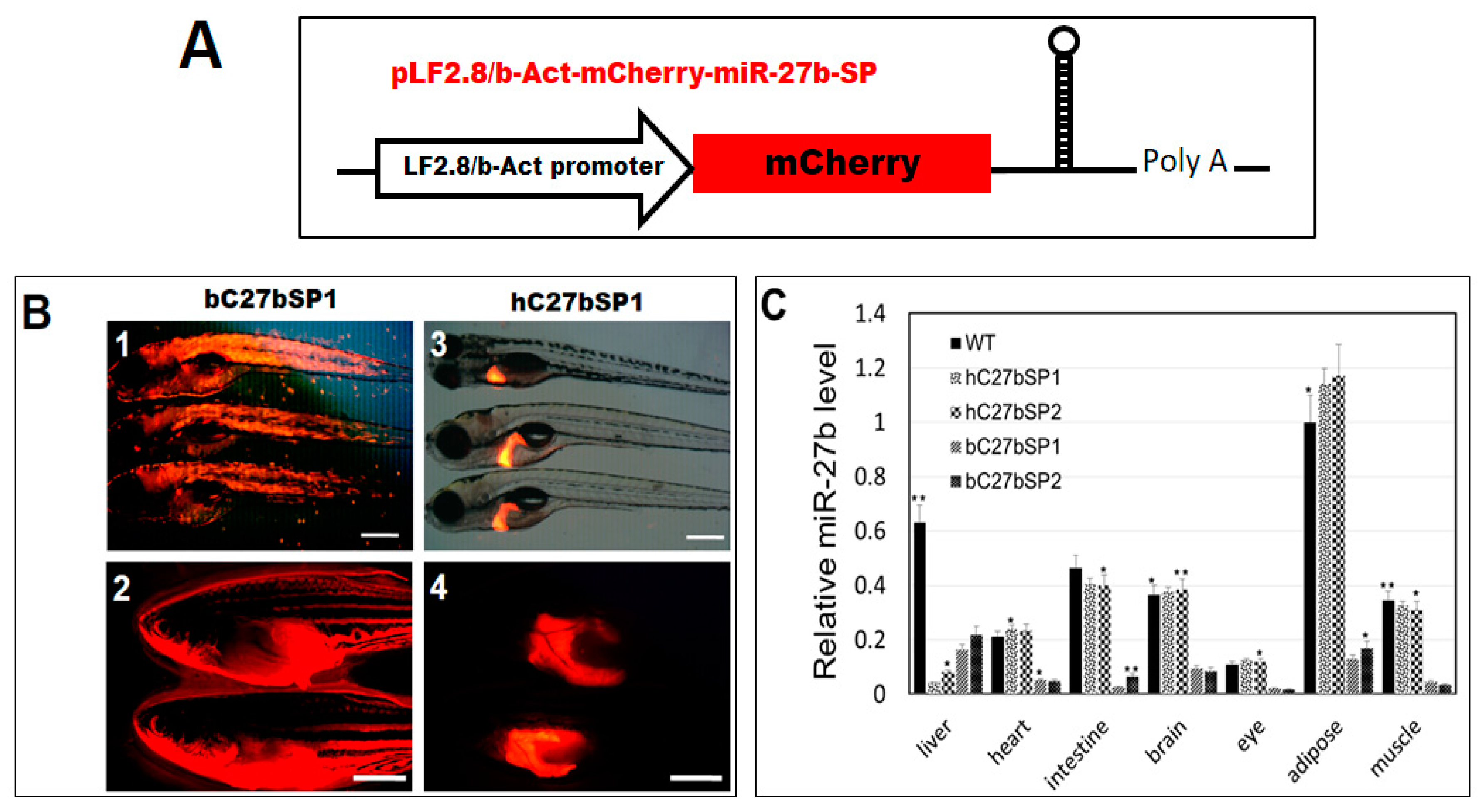
2.1. Designing and Testing the Functionality of the miR-27b Sponge
2.2. Generation of Transgenic C27bSPs (bC27bSP1, 2 and hC27bSP1, 2) Zebrafish Lines
2.3. Inhibition of Endogenous miR-27b Increases Endotrophic and Intravascular Lipid Accumulation
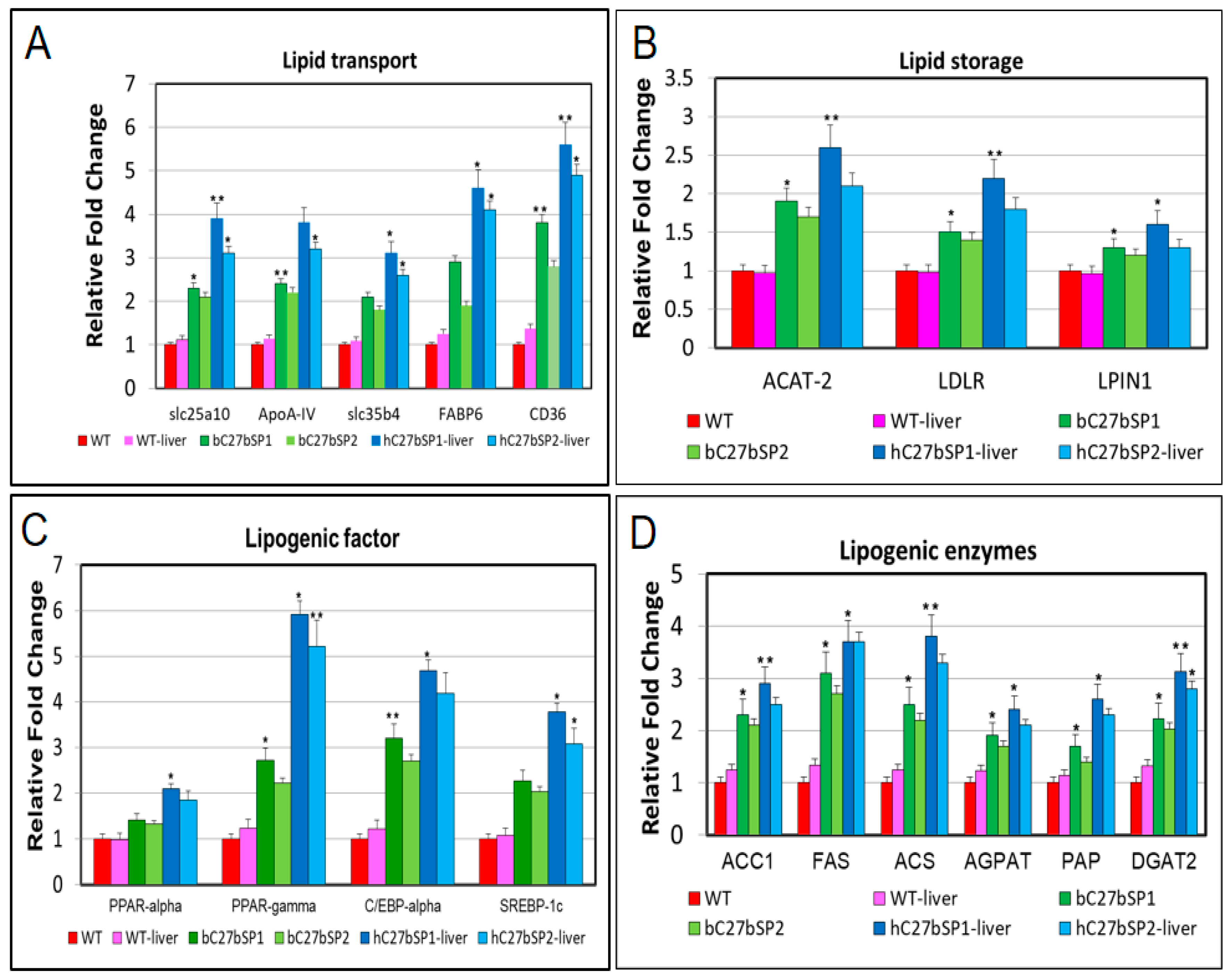
2.4. miR-27b Depletion Increases Expression of Genes Associated with Lipogenesis in C27bSPs
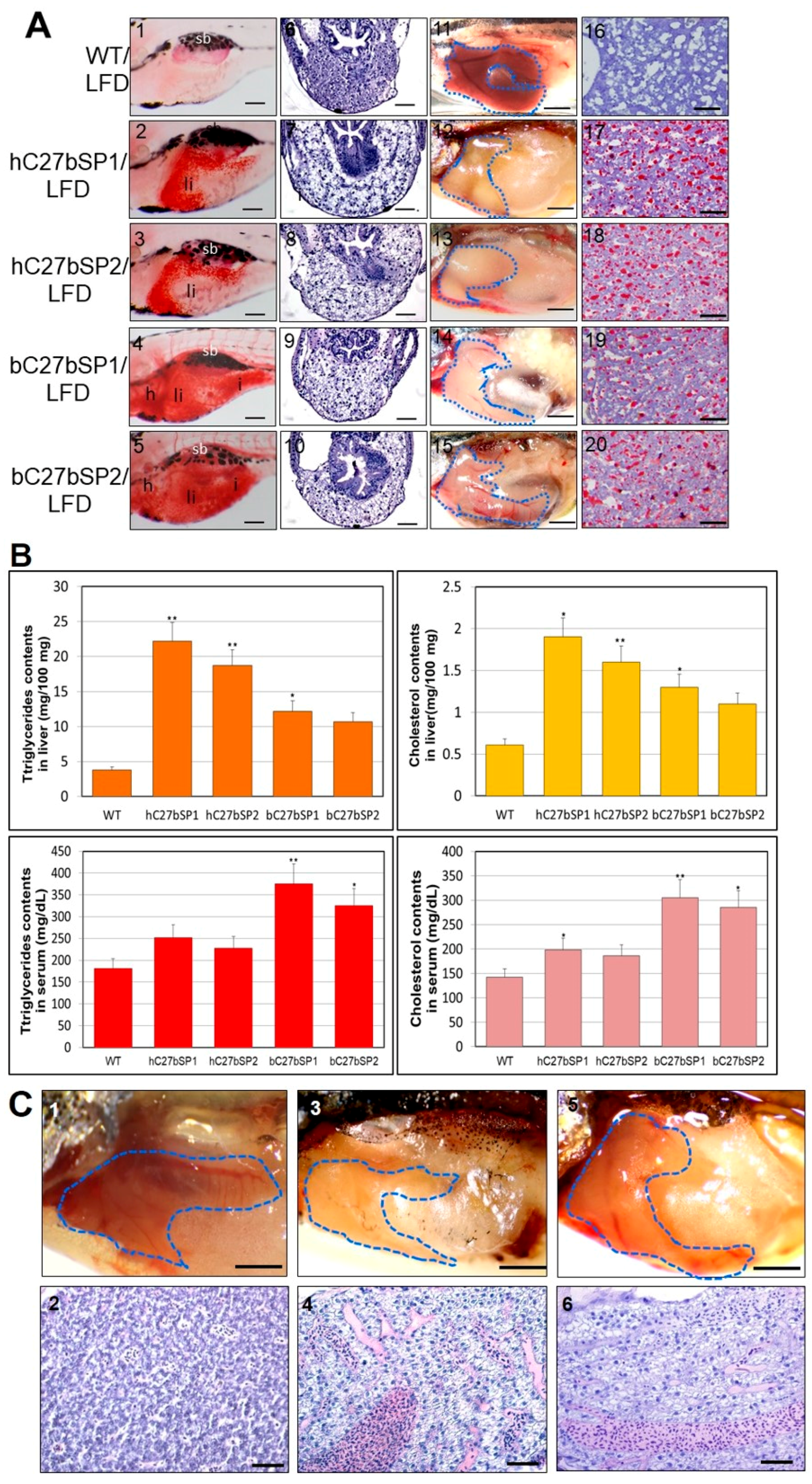
2.5. hC27bSP Adults Developed NAFLD and Eventually NASH
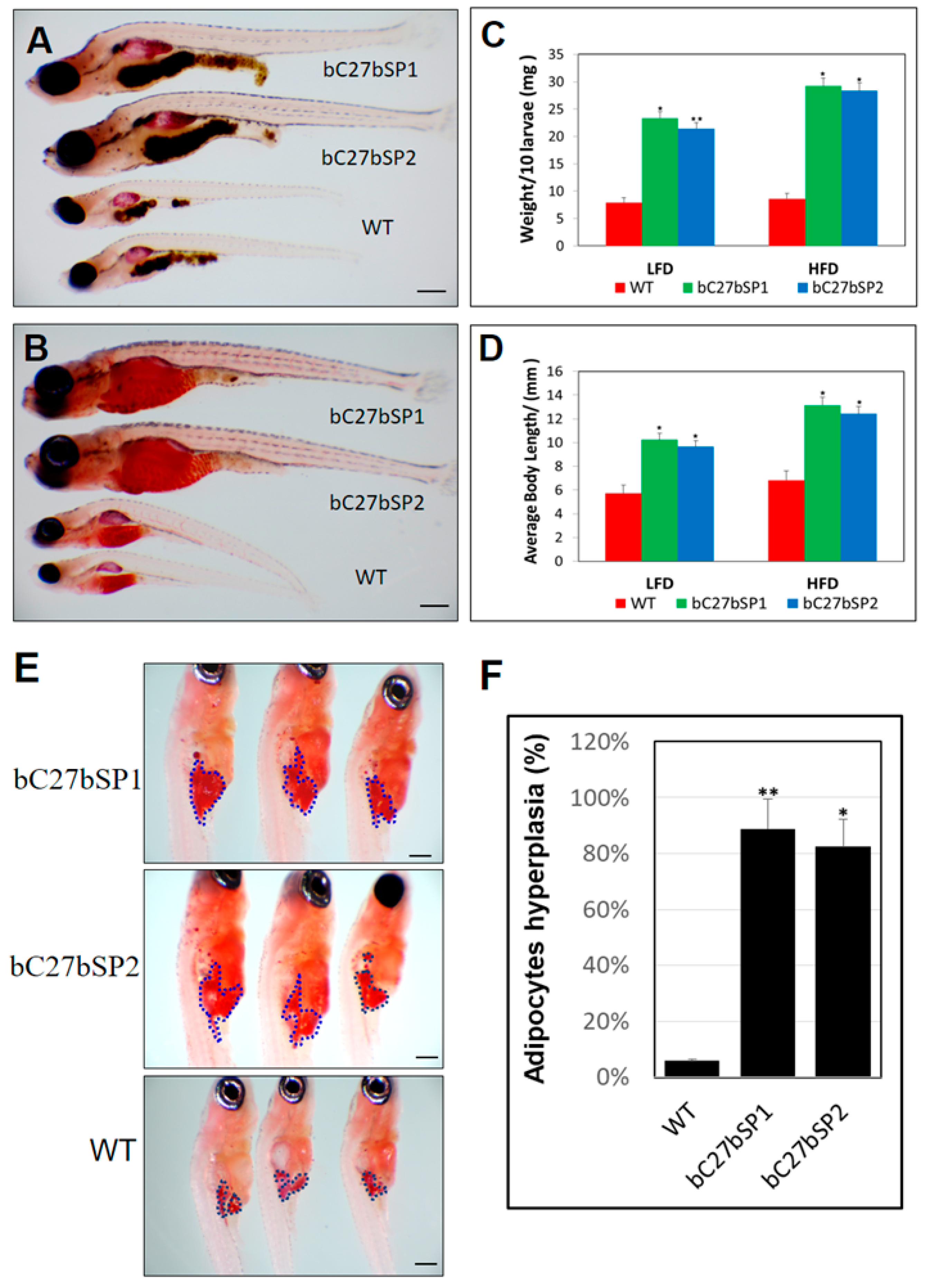
2.6. miR-27b Depletion Induces Early Onset of Adipocyte Hyperplasia in Zebrafish
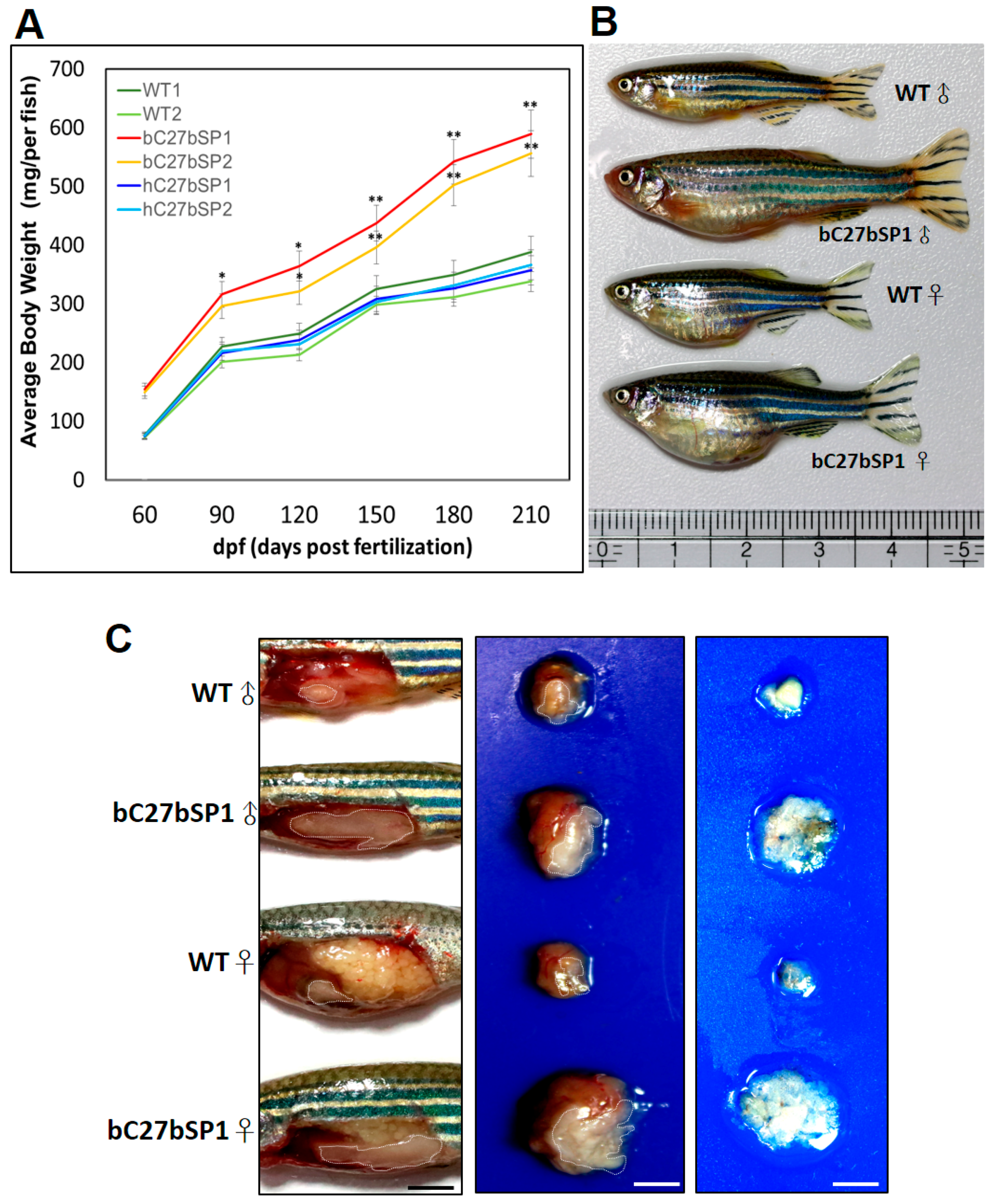
2.7. Adult bC27bSPs are Plump and Have Increased White Adipose Tissue Mass
3. Discussion
4. Materials and Methods
4.1. Plasmid Construction and Oligonucleotides
4.2. Generation and Feeding of hC27bSP and bC27bSP Transgenic Zebrafish
4.3. Cell Culture and Transfection
4.4. Fluorescent Reporter Assay
4.5. Real-Time RT-PCR and Quantification of Mature miR-27b
4.6. Biochemical Analyses of Zebrafish Liver and Blood Lipids
4.7. Western Blot Analysis
4.8. Lipid Staining
4.9. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| NAFLD | nonalcoholic fatty liver disease |
| NASH | nonalcoholic steatohepatitis |
| FLD | fatty liver disease |
| YY1 | Yin Yang 1 |
| CB1R | cannabinoid receptor 1 |
| CHOP-10 | C/EBP homologous protein 10 |
| PPAR-γ | peroxisome proliferator-activated receptor γ |
| C/EBP-α | CCAAT enhancer binding protein-α |
| C/EBP-β | CCAAT enhancer binding protein-β |
| ANGPTL3 | angiopoietin-like 3 |
| GPAM | glycerol-3-phosphate acyltransferase 1 |
| SREBP-1c | sterol regulatory element binding transcription factor 1c |
| miRs | microRNAs |
| miR-SP | microRNA-sponge |
| WAT | white adipose tissue |
References
- Shen, L.; Zhang, Y.; Du, J.; Chen, L.; Luo, J.; Li, X.; Li, M.; Tang, G.; Zhang, S.; Zhu, L. MicroRNA-23a regulates 3T3-L1 adipocyte differentiation. Gene 2016, 575, 761–764. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Xu, G.; Ji, C.; Shi, C.; Shen, Y.; Chen, L.; Zhu, L.; Yang, L.; Zhao, Y.; Guo, X. The role of microRNA-26b in human adipocyte differentiation and proliferation. Gene 2014, 533, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Lie, S.; Morrison, J.L.; Williams-Wyss, O.; Suter, C.M.; Humphreys, D.T.; Ozanne, S.E.; Zhang, S.; MacLaughlin, S.M.; Kleemann, D.O.; Walker, S.K.; et al. Impact of maternal undernutrition around the time of conception on factors regulating hepatic lipid metabolism and microRNAs in singleton and twin fetuses. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E148–E159. [Google Scholar] [CrossRef] [PubMed]
- Massart, J.; Katayama, M.; Krook, A. microManaging glucose and lipid metabolism in skeletal muscle: Role of microRNAs. Biochim. Biophys. Acta 2016, 1861, 2130–2138. [Google Scholar] [CrossRef] [PubMed]
- Baffy, G. MicroRNAs in Nonalcoholic Fatty Liver Disease. J. Clin. Med. 2015, 4, 1977–1988. [Google Scholar] [CrossRef] [PubMed]
- Tessitore, A.; Cicciarelli, G.; del Vecchio, F.; Gaggiano, A.; Verzella, D.; Fischietti, M.; Mastroiaco, V.; Vetuschi, A.; Sferra, R.; Barnabei, R.; et al. MicroRNA expression analysis in high fat diet-induced NAFLD-NASH-HCC progression: Study on C57BL/6J mice. BMC Cancer 2016, 16, 3. [Google Scholar] [CrossRef] [PubMed]
- Parrizas, M.; Novials, A. Circulating microRNAs as biomarkers for metabolic disease. Best Pract. Res. Clin. Endocrinol. Metab. 2016, 30, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Willeit, P.; Skroblin, P.; Kiechl, S.; Fernandez-Hernando, C.; Mayr, M. Liver microRNAs: Potential mediators and biomarkers for metabolic and cardiovascular disease? Eur. Heart J. 2016, 37, 3260–3266. [Google Scholar] [CrossRef] [PubMed]
- Rocic, P. Can microRNAs be biomarkers or targets for therapy of ischemic coronary artery disease in metabolic syndrome? Curr. Drug Targets 2016, 18, 1722–1732. [Google Scholar] [CrossRef] [PubMed]
- Deiuliis, J.A. MicroRNAs as regulators of metabolic disease: Pathophysiologic significance and emerging role as biomarkers and therapeutics. Int. J. Obes. (Lond.) 2016, 40, 88–101. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, D.M.; Simao, A.L.; Rodrigues, C.M.; Castro, R.E. Revisiting the metabolic syndrome and paving the way for microRNAs in non-alcoholic fatty liver disease. FEBS J. 2014, 281, 2503–2524. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Hernando, C.; Ramirez, C.M.; Goedeke, L.; Suarez, Y. MicroRNAs in metabolic disease. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Zampetaki, A.; Mayr, M. MicroRNAs in vascular and metabolic disease. Circ. Res. 2012, 110, 508–522. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, M.; Kishikawa, T.; Yoshikawa, T.; Yamagami, M.; Ohno, M.; Takata, A.; Shibata, C.; Ishibashi, R.; Koike, K. MicroRNAs and liver disease. J. Hum. Genet. 2017, 62, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Sobolewski, C.; Calo, N.; Portius, D.; Foti, M. MicroRNAs in fatty liver disease. Semin. Liver Dis. 2015, 35, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Szabo, G.; Bala, S. MicroRNAs in liver disease. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 542–552. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.W.; Heegaard, N.H.; Orum, H. MicroRNAs in liver disease. Gastroenterology 2012, 142, 1431–1443. [Google Scholar] [CrossRef] [PubMed]
- Karbiener, M.; Fischer, C.; Nowitsch, S.; Opriessnig, P.; Papak, C.; Ailhaud, G.; Dani, C.; Amri, E.-Z.; Scheideler, M. microRNA miR-27b impairs human adipocyte differentiation and targets PPAR-γ. Biochem. Biophys. Res. Commun. 2009, 390, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Zou, B.; Ge, Z.; Zhu, W.; Xu, Z.; Li, C. Persimmon tannin represses 3T3-L1 preadipocyte differentiation via up-regulating expression of miR-27 and down-regulating expression of peroxisome proliferator-activated receptor-γ in the early phase of adipogenesis. Eur. J. Nutr. 2015, 54, 1333–1343. [Google Scholar] [CrossRef] [PubMed]
- Kang, T.; Lu, W.; Xu, W.; Anderson, L.; Bacanamwo, M.; Thompson, W.; Chen, Y.E.; Liu, D. MicroRNA-27 (miR-27) targets prohibitin and impairs adipocyte differentiation and mitochondrial function in human adipose-derived stem cells. J. Biol. Chem. 2013, 288, 34394–34402. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Yu, J.; Bi, J.; Qi, H.; Di, W.; Wu, L.; Wang, L.; Zha, J.; Lv, S.; Zhang, F.; et al. Glucocorticoids transcriptionally regulate miR-27b expression promoting body fat accumulation via suppressing the browning of white adipose tissue. Diabetes 2015, 64, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Vickers, K.C.; Shoucri, B.M.; Levin, M.G.; Wu, H.; Pearson, D.S.; Osei-Hwedieh, D.; Collins, F.S.; Remaley, A.T.; Sethupathy, P. MicroRNA-27b is a regulatory hub in lipid metabolism and is altered in dyslipidemia. Hepatology 2013, 57, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Zeituni, E.M.; Farber, S.A. Studying Lipid Metabolism and Transport During Zebrafish Development. Methods Mol. Biol. 2016, 1451, 237–255. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Dai, Z.; Shi, C.; Zhai, G.; Jin, X.; He, J.; Lou, Q.; Yin, Z. Depletion of myostatin b promotes somatic growth and lipid metabolism in zebrafish. Front. Endocrinol. 2016, 7, 88. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Lv, S.; Liu, J.; Nie, S.; Chen, J.; Dong, Q.; Huang, C.; Yang, D. Chronic perfluorooctanesulfonic acid exposure disrupts lipid metabolism in zebrafish. Hum. Exp. Toxicol. 2016, 36, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.C.; Hsiao, C.D.; Kawakami, K.; Tse, W.K. Triclosan (TCS) exposure impairs lipid metabolism in zebrafish embryos. Aquat. Toxicol. 2016, 173, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.L.; Carten, J.D.; Farber, S.A. Zebrafish lipid metabolism: From mediating early patterning to the metabolism of dietary fat and cholesterol. Methods Cell Biol. 2011, 101, 111–141. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, A. Studying non-alcoholic fatty liver disease with zebrafish: A confluence of optics, genetics, and physiology. Cell. Mol. Life Sci. 2012, 69, 3953–3961. [Google Scholar] [CrossRef] [PubMed]
- Shieh, Y.S.; Chang, Y.S.; Hong, J.R.; Chen, L.J.; Jou, L.K.; Hsu, C.C.; Her, G.M. Increase of hepatic fat accumulation by liver specific expression of Hepatitis B virus X protein in zebrafish. Biochim. Biophys. Acta 2010, 1801, 721–730. [Google Scholar] [CrossRef] [PubMed]
- Her, G.M.; Hsu, C.C.; Hong, J.R.; Lai, C.Y.; Hsu, M.C.; Pang, H.W.; Chan, S.K.; Pai, W.Y. Overexpression of gankyrin induces liver steatosis in zebrafish (Danio rerio). Biochim. Biophys. Acta 2011, 1811, 536–548. [Google Scholar] [CrossRef] [PubMed]
- Her, G.M.; Pai, W.Y.; Lai, C.Y.; Hsieh, Y.W.; Pang, H.W. Ubiquitous transcription factor YY1 promotes zebrafish liver steatosis and lipotoxicity by inhibiting CHOP-10 expression. Biochim. Biophys. Acta 2013, 1831, 1037–1051. [Google Scholar] [CrossRef] [PubMed]
- Pai, W.Y.; Hsu, C.C.; Lai, C.Y.; Chang, T.Z.; Tsai, Y.L.; Her, G.M. Cannabinoid receptor 1 promotes hepatic lipid accumulation and lipotoxicity through the induction of SREBP-1c expression in zebrafish. Transgen. Res. 2013, 22, 823–838. [Google Scholar] [CrossRef] [PubMed]
- Goedeke, L.; Rotllan, N.; Ramirez, C.M.; Aranda, J.F.; Canfran-Duque, A.; Araldi, E.; Fernandez-Hernando, A.; Langhi, C.; de Cabo, R.; Baldan, A.; et al. miR-27b inhibits LDLR and ABCA1 expression but does not influence plasma and hepatic lipid levels in mice. Atherosclerosis 2015, 243, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Li, L.; Zhang, M.; Cheng, H.P.; Gong, D.; Lv, Y.C.; Yao, F.; He, P.P.; Ouyang, X.P.; Lan, G.; et al. MicroRNA-27 prevents atherosclerosis by suppressing lipoprotein lipase-induced lipid accumulation and inflammatory response in apolipoprotein E knockout mice. PLoS ONE 2016, 11, e0157085. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.J.; Yin, K.; Zhao, G.J.; Fu, Y.C.; Tang, C.K. The magic and mystery of microRNA-27 in atherosclerosis. Atherosclerosis 2012, 222, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.M.; Tao, J.; Chen, D.D.; Cai, J.J.; Irani, K.; Wang, Q.; Yuan, H.; Chen, A.F. MicroRNA miR-27b rescues bone marrow-derived angiogenic cell function and accelerates wound healing in type 2 diabetes mellitus. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wu, J.F.; Chen, W.J.; Tang, S.L.; Mo, Z.C.; Tang, Y.Y.; Li, Y.; Wang, J.L.; Liu, X.Y.; Peng, J.; et al. microRNA-27a/b regulates cellular cholesterol efflux, influx and esterification/hydrolysis in THP-1 macrophages. Atherosclerosis 2014, 234, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Linden, D.; William-Olsson, L.; Ahnmark, A.; Ekroos, K.; Hallberg, C.; Sjogren, H.P.; Becker, B.; Svensson, L.; Clapham, J.C.; Oscarsson, J.; et al. Liver-directed overexpression of mitochondrial glycerol-3-phosphate acyltransferase results in hepatic steatosis, increased triacylglycerol secretion and reduced fatty acid oxidation. FASEB J. 2006, 20, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Musunuru, K.; Pirruccello, J.P.; Do, R.; Peloso, G.M.; Guiducci, C.; Sougnez, C.; Garimella, K.V.; Fisher, S.; Abreu, J.; Barry, A.J.; et al. Exome sequencing, ANGPTL3 mutations, and familial combined hypolipidemia. N. Engl. J. Med. 2010, 363, 2220–2227. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Zhang, J.; Huang, G.; Qian, J.; Wang, X.; Mei, S. Over-expressed microRNA-27a and 27b influence fat accumulation and cell proliferation during rat hepatic stellate cell activation. FEBS Lett. 2009, 583, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Vacaru, A.M.; di Narzo, A.F.; Howarth, D.L.; Tsedensodnom, O.; Imrie, D.; Cinaroglu, A.; Amin, S.; Hao, K.; Sadler, K.C. Molecularly defined unfolded protein response subclasses have distinct correlations with fatty liver disease in zebrafish. Dis. Model. Mech. 2014, 7, 823–835. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Lim, B.; Lodish, H.F. microRNAs induced during adipogenesis that accelerate fat cell development are downregulated in obesity. Diabetes 2009, 58, 1050–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Li, Y.C.; Wang, J.; Kong, J.; Qi, Y.; Quigg, R.J.; Li, X. miR-17-92 cluster accelerates adipocyte differentiation by negatively regulating tumor-suppressor Rb2/p130. Proc. Natl. Acad. Sci. USA 2008, 105, 2889–2894. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Gao, Z.; Alarcon, R.M.; Ye, J.; Yun, Z. A role of miR-27 in the regulation of adipogenesis. FEBS J. 2009, 276, 2348–2358. [Google Scholar] [CrossRef] [PubMed]
- Hilton, C.; Neville, M.; Karpe, F. microRNAs in adipose tissue: Their role in adipogenesis and obesity. Int. J. Obes. 2013, 37, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Yu, S.; Li, H.; Xiang, H.; Peng, J.; Jiang, S. microRNAs: Emerging roles in adipogenesis and obesity. Cell Signal. 2014, 26, 1888–1896. [Google Scholar] [CrossRef] [PubMed]
- Flynn, E.J., III; Trent, C.M.; Rawls, J.F. Ontogeny and nutritional control of adipogenesis in zebrafish (Danio rerio). J. Lipid Res. 2009, 50, 1641–1652. [Google Scholar] [CrossRef] [PubMed]
- Den Broeder, M.J.; Kopylova, V.A.; Kamminga, L.M.; Legler, J. Zebrafish as a model to study the role of peroxisome proliferating-activated receptors in adipogenesis and obesity. PPAR Res. 2015, 2015, 358029. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Trajkovski, M. miR-27 orchestrates the transcriptional regulation of brown adipogenesis. Metabolism 2014, 63, 272–282. [Google Scholar] [CrossRef] [PubMed]
- Chan, L.S.; Yue, P.Y.; Kok, T.W.; Keung, M.H.; Mak, N.K.; Wong, R.N. Ginsenoside-Rb1 promotes adipogenesis through regulation of PPAR-γ and microRNA-27b. Horm. Metab. Res. 2012, 44, 819–824. [Google Scholar] [PubMed]
- Seale, P.; Conroe, H.M.; Estall, J.; Kajimura, S.; Frontini, A.; Ishibashi, J.; Cohen, P.; Cinti, S.; Spiegelman, B.M. Prdm16 determines the thermogenic program of subcutaneous white adipose tissue in mice. J. Clin. Investig. 2011, 121, 96–105. [Google Scholar] [CrossRef] [PubMed]
- May, F.J.; Baer, L.A.; Lehnig, A.C.; So, K.; Chen, E.Y.; Gao, F.; Narain, N.R.; Gushchina, L.; Rose, A.; Doseff, A.I.; et al. Lipidomic adaptations in white and brown adipose tissue in response to exercise demonstrate molecular species-specific remodeling. Cell Rep. 2017, 18, 1558–1572. [Google Scholar] [CrossRef] [PubMed]
- Imrie, D.; Sadler, K.C. White adipose tissue development in zebrafish is regulated by both developmental time and fish size. Dev. Dyn. 2010, 239, 3013–3023. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.; Fu, Y.F.; Du, T.T.; Jing, C.B.; Fu, C.T.; Chen, Y.; Jin, Y.; Deng, M.; Liu, T.X. Heritable and lineage-specific gene knockdown in zebrafish embryo. PLoS ONE 2009, 4, e6125. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.J.; Hsu, C.C.; Hong, J.R.; Jou, L.K.; Tseng, H.C.; Wu, J.L.; Liou, Y.C.; Her, G.M. Liver-specific expression of p53-negative regulator mdm2 leads to growth retardation and fragile liver in zebrafish. Dev. Dyn. 2008, 237, 1070–1081. [Google Scholar] [CrossRef] [PubMed]
- Her, G.M.; Chiang, C.C.; Chen, W.Y.; Wu, J.L. In vivo studies of liver-type fatty acid binding protein (L-FABP) gene expression in liver of transgenic zebrafish (Danio rerio). FEBS Lett. 2003, 538, 125–133. [Google Scholar] [CrossRef]
- Pedroso, G.L.; Hammes, T.O.; Escobar, T.D.; Fracasso, L.B.; Forgiarini, L.F.; da Silveira, T.R. Blood collection for biochemical analysis in adult zebrafish. J. Vis. Exp. 2012, 63, e3865. [Google Scholar] [CrossRef] [PubMed]
- Yeh, K.Y.; Lai, C.Y.; Lin, C.Y.; Hsu, C.C.; Lo, C.P.; Her, G.M. ATF4 overexpression induces early onset of hyperlipidaemia and hepatic steatosis and enhances adipogenesis in zebrafish. Sci. Rep. 2017, 7, 16362. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession | Forward Primer | Reverse Primer |
|---|---|---|---|
| PPAR-α | NM_001102567 | CTGCGGGACATCTCTCAGTC | ACCGTAAACACCTGACGACG |
| PPAR-γ | DQ839547 | CCTGTCCGGGAAGACCAGCG | GTGCTCGTGGAGCGGCATGT |
| C/EBP-α | NM_131885 | AACGGAGCGAGCTTGACTT | AAATCATGCCCATTAGCTGC |
| SREBP-1c | NM_001105129 | CAGAGGGTGGGCATGCTGGC | ACCTGGTTCTGGATGAATCG |
| il1b | NM_212844 | TGGCGAACGTCATCCAAG | GGAGCACTGGGCGACGCATA |
| il6 | NM_001261449 | AGACCGCTGCCTGTCTAAAA | TTTGATGTCGTTCACCAGGA |
| tnfa | NM_212859 | GCTTATGAGCCATGCAGTGA | TGCCCAGTCTGTCTCCTTCT |
| ifn-γ | NM_212864.1 | TGGGCGATCAAGGAAAACGA | TTGATGCTTTAGCCTGCCGT |
| nfkb2 | NM_001001840.3 | AAACAAGACGCAAGGAGCCC | GCTGAAGGAAACGTCATAGGC |
| NF-κB | BC162402 | GAAGCATTCAGGCTCGGTGA | CAGGTCTGTCGGTCCCTTTC |
| atf6 | NM_001110519 | CTGTGGTGAAACCTCCACCT | CATGGTGACCACAGGAGATG |
| ern2 | XM_001919315 | TGACGTGGTGGAAGTTGGTA | ACGGATCACATTGGGATGTT |
| ire1 | XM_009306258 | TGACGTGGTGGAAGTTGGTA | ACGGATCACATTGGGATGTT |
| eif2ak3(perk) | XM_005156585 | TGGGCTCTGAAGAGTTCGAT | TGTGAGCCTTCTCCGTCTTT |
| hspa5(bip) | NM_213058 | AAGAGGCCGAAGAGAAGGAC | AGCAGCAGAGCCTCGAAATA |
| ddit3(chop) | NM_001082825 | AAGGAAAGTGCAGGAGCTGA | TCACGCTCTCCACAAGAAGA |
| β-Actin | NM_131031.1 | GCCACCTTAAATGGCCTAGCA | GCCATACAGAGCAGAAGCCA |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, C.-C.; Lai, C.-Y.; Lin, C.-Y.; Yeh, K.-Y.; Her, G.M. MicroRNA-27b Depletion Enhances Endotrophic and Intravascular Lipid Accumulation and Induces Adipocyte Hyperplasia in Zebrafish. Int. J. Mol. Sci. 2018, 19, 93. https://doi.org/10.3390/ijms19010093
Hsu C-C, Lai C-Y, Lin C-Y, Yeh K-Y, Her GM. MicroRNA-27b Depletion Enhances Endotrophic and Intravascular Lipid Accumulation and Induces Adipocyte Hyperplasia in Zebrafish. International Journal of Molecular Sciences. 2018; 19(1):93. https://doi.org/10.3390/ijms19010093
Chicago/Turabian StyleHsu, Chia-Chun, Chi-Yu Lai, Chiu-Ya Lin, Kun-Yun Yeh, and Guor Mour Her. 2018. "MicroRNA-27b Depletion Enhances Endotrophic and Intravascular Lipid Accumulation and Induces Adipocyte Hyperplasia in Zebrafish" International Journal of Molecular Sciences 19, no. 1: 93. https://doi.org/10.3390/ijms19010093
APA StyleHsu, C. -C., Lai, C. -Y., Lin, C. -Y., Yeh, K. -Y., & Her, G. M. (2018). MicroRNA-27b Depletion Enhances Endotrophic and Intravascular Lipid Accumulation and Induces Adipocyte Hyperplasia in Zebrafish. International Journal of Molecular Sciences, 19(1), 93. https://doi.org/10.3390/ijms19010093