Highly Efficient Targeted Gene Editing in Upland Cotton Using the CRISPR/Cas9 System


Abstract
:1. Introduction
2. Result
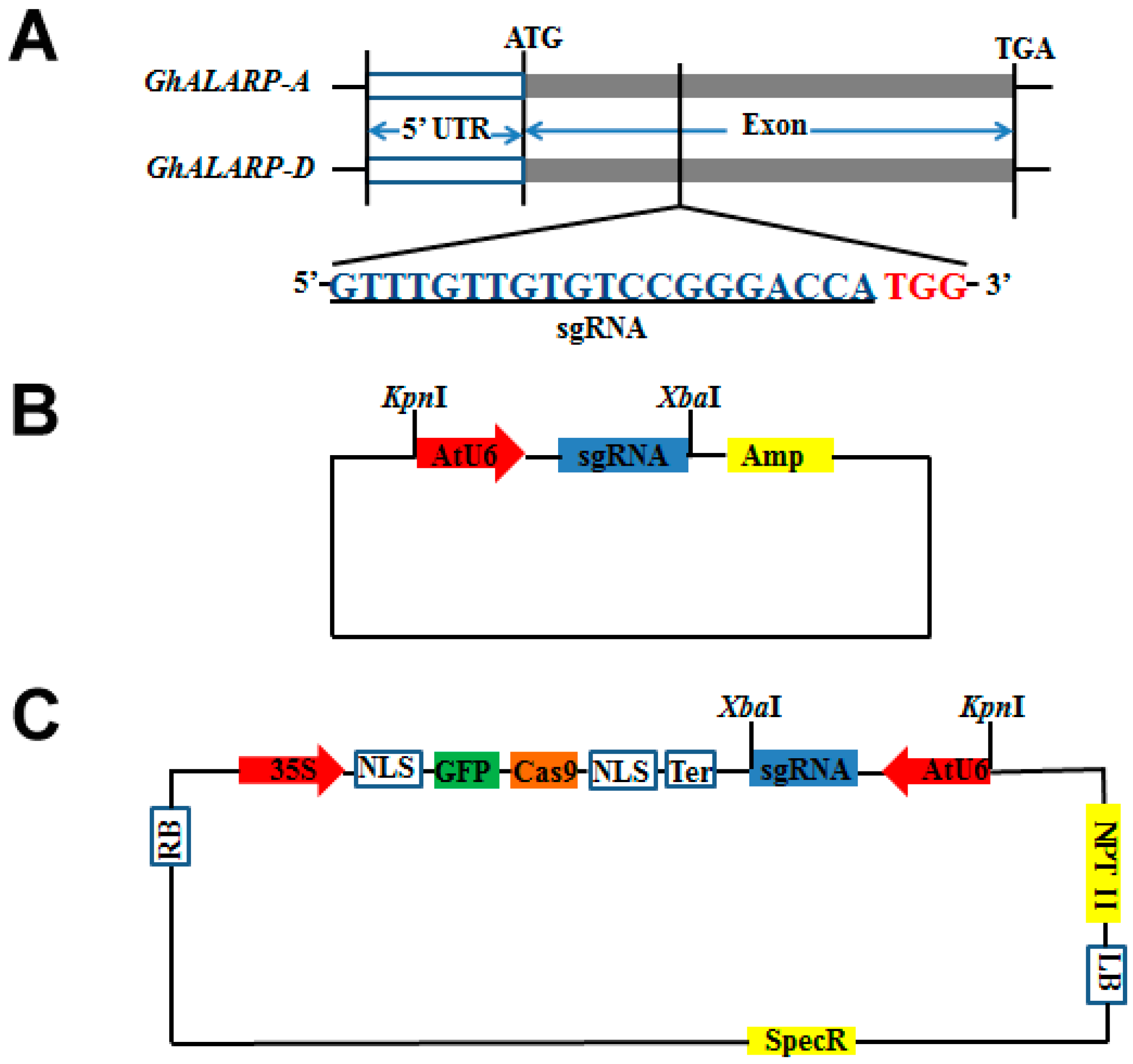
2.1. Construction of the CRISPR/Cas9-GhALARP-sgRNA Vector
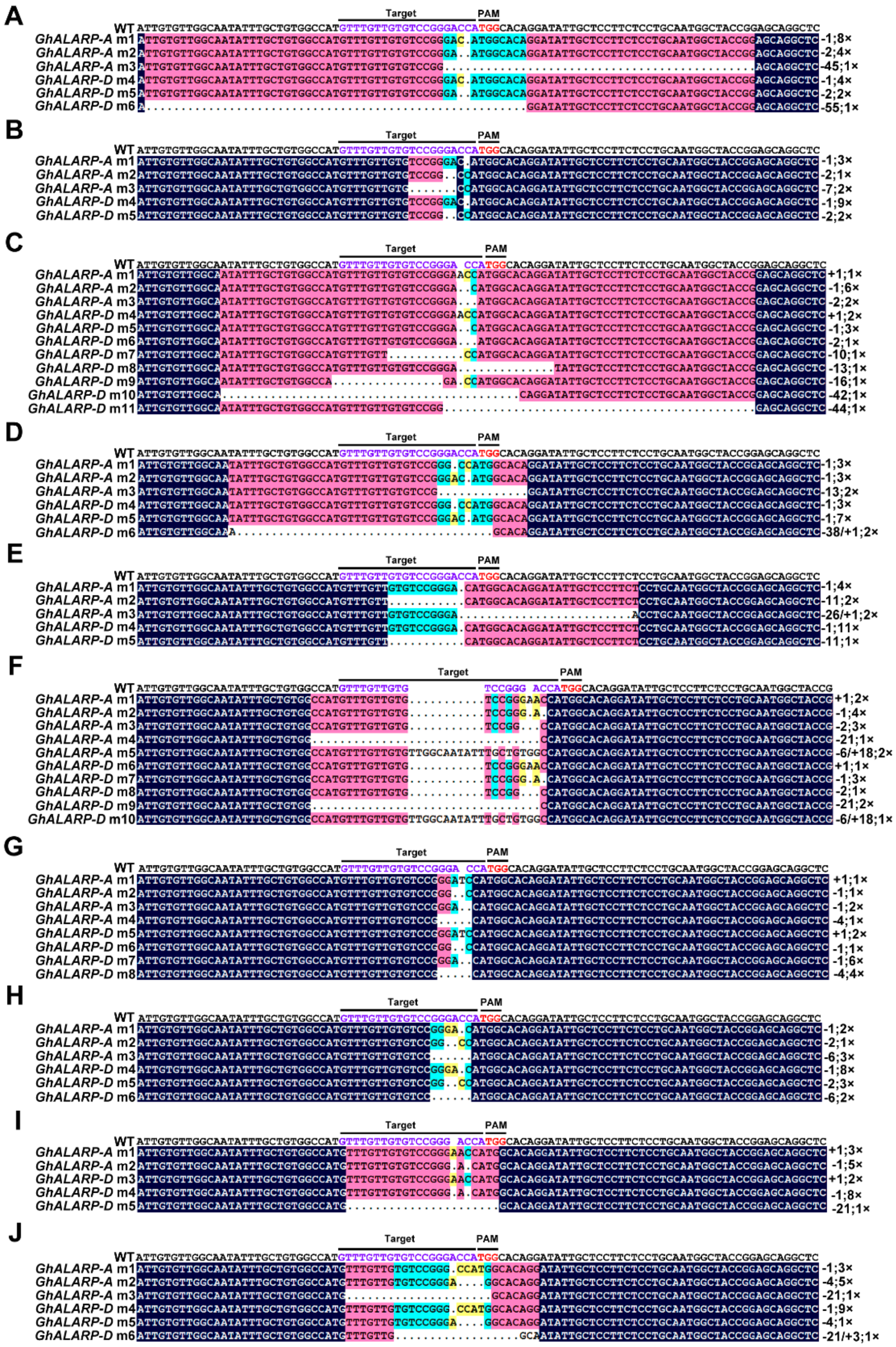
2.2. Analysis of CRISPR/Cas9-sgRNA-Mediated Mutagenesis in GhALARP-A and GhALARP-D
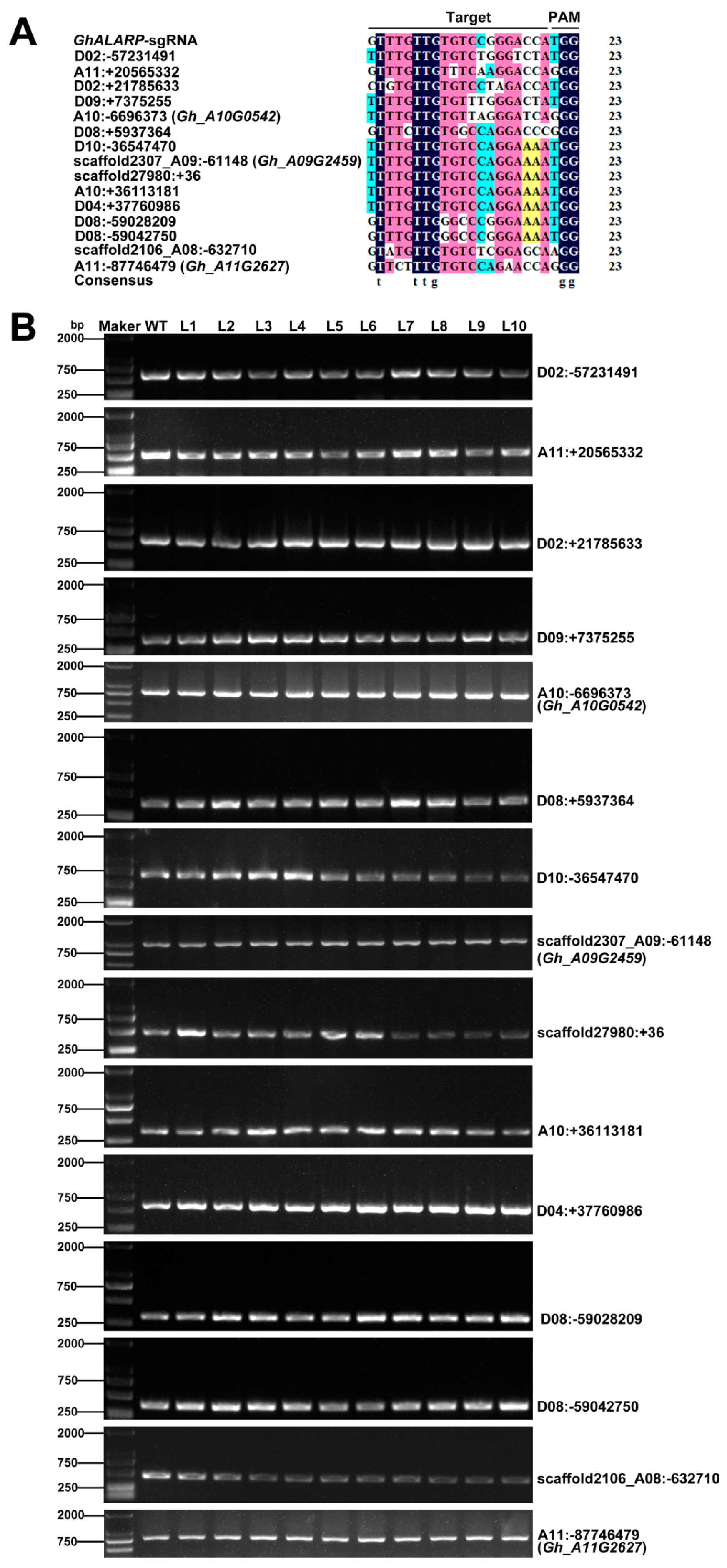
2.3. Off-Target Analysis in the Transgenic Cotton Lines
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth
4.2. Selection of sgRNA Targeting GhALARP
4.3. Construction of 35S-Cas9-AtU6-GhALARP-sgRNA Vector
4.4. Agrobacterium-Mediated Transformation of Cotton
4.5. Mutant Identification and Analysis of Genomic Target Site
4.6. Analysis of Possible Off-Target Sites
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Esvelt, K.M.; Wang, H.H. Genome-scale engineering for systems and synthetic biology. Mol. Syst. Biol. 2013, 9, 641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaj, T.; Gersbach, C.A.; Barbas, C.F. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2013, 31, 397–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bibikova, M.; Golic, M.M.; Golic, K.G.; Carroll, D. Targeted chromosomal cleavage and mutagenesis in Drosophila using zinc-finger nucleases. Genetics 2002, 161, 1169–1175. [Google Scholar] [PubMed]
- Bibikova, M.; Beumer, K.J.; Trautman, J.K.; Carroll, D. Enhancing gene targeting with designed zinc finger nucleases. Science 2003, 300, 764. [Google Scholar] [CrossRef] [PubMed]
- Dreier, B.; Fuller, R.; Segal, D.J.; Lund, C.V.; Blancafort, P.; Huber, A.; Koksch, B.; Barbas, C.F. Development of zinc finger domains for recognition of the 5-CNN-3 family DNA sequences and their use in the construction of artificial transcription factors. J. Biol. Chem. 2005, 280, 35588–35597. [Google Scholar] [CrossRef] [PubMed]
- Shan, Q.W.; Wang, Y.P.; Chen, K.L.; Liang, Z.; Li, J.; Zhang, Y.; Zhang, K.; Liu, J.X.; Voytas, D.F.; Zheng, X.L.; et al. Rapid and efficient gene modification in rice and Brachypodium using TALENs. Mol. Plant 2013, 6, 1365–1368. [Google Scholar] [CrossRef] [PubMed]
- Hockemeyer, D.; Wang, H.Y.; Kiani, S.; Lai, C.S.; Gao, Q.; Cassady, J.P.; Cost, G.J.; Zhang, L.; Santiago, Y.; Miller, J.C.; et al. Genetic engineering of human pluripotent cells using TALE nucleases. Nat. Biotechnol. 2011, 29, 731–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, P.; Xiao, A.; Zhou, M.G.; Zhu, Z.Y.; Lin, S.; Zhang, B. Heritable gene targeting in zebrafish using customized TALENs. Nat. Biotechnol. 2011, 29, 699–700. [Google Scholar] [CrossRef] [PubMed]
- Tesson, L.; Usal, C.; Menoret, S.; Leung, E.; Niles, B.J.; Remy, S.; Santiago, Y.; Vincent, A.I.; Meng, X.D.; Zhang, L.; et al. Knockout rats generated by embryo microinjection of TALENs. Nat. Biotechnol. 2011, 29, 695–696. [Google Scholar] [CrossRef] [PubMed]
- Nekrasov, V.; Staskawicz, B.J.; Weigel, D.; Jones, J.D.; Kamoun, S. Targeted mutagenesis in the model plant Nicotiana benthamiana using Cas9 RNA-guided endonuclease. Nat. Biotechnol. 2013, 31, 691–693. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.Y.; Zhang, B.T.; Ding, W.N.; Liu, X.D.; Yang, D.L.; Wei, P.L.; Cao, F.Q.; Zhu, S.H.; Zhang, F.; Mao, Y.F.; et al. Efficient genome editing in plants using a CRISPR/Cas system. Cell Res. 2013, 23, 1229–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, Q.W.; Wang, Y.P.; Li, J.; Zhang, Y.; Chen, K.L.; Liang, Z.; Zhang, K.; Liu, J.X.; Xi, J.Z.; Qiu, J.L.; et al. Targeted genome modification of crop plants using a CRISPR-Cas system. Nat. Biotechnol. 2013, 31, 686–688. [Google Scholar] [CrossRef] [PubMed]
- Jiao, R.J.; Gao, C.X. The CRISPR/Cas9 genome editing revolution. J. Genet. Genom. 2016, 43, 227–228. [Google Scholar] [CrossRef] [PubMed]
- Demirci, Y.; Zhang, B.H.; Unver, T. CRISPR/Cas9: An RNA-guided highly precise synthetic tool for plant genome editing. J. Cell Physiol. 2018, 233, 1844–1859. [Google Scholar] [CrossRef] [PubMed]
- Mahfouz, M.M.; Li, L.X.; Shamimuzzaman, M.; Wibowo, A.; Fang, X.Y.; Zhu, J.K. De novo-engineered transcription activator-like effector (TALE) hybrid nuclease with novel DNA binding specificity creates double-strand breaks. Proc. Natl. Acad. Sci. USA 2011, 108, 2623–2628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cong, L.; Ran, F.A.; Cox, D.M.; Lin, S.L.; Barretto, R.P.; Habib, N.; Hsu, P.; Wu, X.B.; Jiang, W.Y.; Marraffini, L.A.; et al. Multiplex genome engineering using CRISPR/Cas systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Mali, P.; Yang, L.; Esvelt, K.M.; Aach, J.; Guell, M.; Dicarlo, J.E.; Norville, J.E.; Church, G.M. RNA-guided human genome engineering via Cas9. Science 2013, 339, 823–826. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.Y.; Mao, Y.F.; Xu, N.F.; Zhang, B.T.; Wei, P.L.; Yang, D.L.; Wang, Z.; Zhang, Z.J.; Zheng, R.; Yang, L.; et al. Multigeneration analysis reveals the inheritance, specificity, and patterns of CRISPR/Cas-induced gene modifications in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 4632–4637. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.Z.; Yang, B.; Weeks, D.P. Efficient CRISPR/Cas9-mediated gene editing in Arabidopsis thaliana and inheritance of modified genes in the T2 and T3 generations. PLoS ONE 2014, 9, e99225. [Google Scholar] [CrossRef] [PubMed]
- Xie, K.B.; Yang, Y.N. RNA-guided genome editing in plants using a CRISPR-Cas system. Mol. Plant 2013, 6, 1975–1983. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.B.; Yu, H.; Zhang, Y.; Zhuang, F.F.; Song, X.G.; Gao, S.S.; Gao, C.X.; Li, J.Y. Construction of a genome-wide mutant library in rice using CRISPR/Cas9. Mol. Plant 2017, 10, 1238–1241. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Zhang, K.; Chen, K.L.; Gao, C.X. Targeted mutagenesis in Zea mays using TALENs and the CRISPR/Cas system. J. Genet. Genom. 2014, 41, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Su, H.D.; Bai, H.; Wang, R.; Liu, Y.L.; Guo, X.R.; Liu, C.; Zhang, J.; Yuan, J.; Birchler, J.A.; et al. High efficiency genome editing using a dmc1 promoter controlled CRISPR/Cas9 system in maize. Plant Biotechnol. J. 2018. [Google Scholar] [CrossRef] [PubMed]
- Li, J.F.; Norville, J.E.; Aach, J.; Mccormack, M.; Zhang, D.D.; Bush, J.; Church, G.M.; Sheen, J. Multiplex and homologous recombination-mediated genome editing in Arabidopsis and Nicotiana benthamiana using guide RNA and Cas9. Nat. Biotechnol. 2013, 31, 688–691. [Google Scholar] [CrossRef] [PubMed]
- Ron, M.; Kajala, K.; Pauluzzi, G.; Wang, D.X.; Reynoso, M.; Zumstein, K.; Garcha, J.; Winte, S.; Masson, H.; Inagaki, S.; et al. Hairy root transformation using Agrobacterium rhizogenes as a tool for exploring cell type-specific gene expression and function using tomato as a model. Plant Physiol. 2014, 166, 455–469. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.Z.; Zhou, H.B.; Bi, H.H.; Fromm, M.E.; Yang, B.; Weeks, D.P. Demonstration of CRISPR/Cas9/sgRNA-mediated targeted gene modification in Arabidopsis, tobacco, sorghum and rice. Nucleic Acids Res. 2013, 41, e188. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, T.B.; Lafayette, P.R.; Schmitz, R.J.; Parrott, W.A. Targeted genome modifications in soybean with CRISPR/Cas9. BMC Biotechnol. 2015, 15, 16. [Google Scholar] [CrossRef] [PubMed]
- Michno, J.M.; Wang, X.B.; Liu, J.Q.; Curtin, S.J.; Kono, T.J.; Stupar, R.M. CRISPR/Cas mutagenesis of soybean and Medicago truncatula using a new web-tool and a modified Cas9 enzyme. GM Crops Food 2015, 6, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Fan, D.; Liu, T.T.; Li, C.F.; Jiao, B.; Li, S.; Hou, Y.S.; Luo, K.M. Efficient CRISPR/Cas9-mediated targeted mutagenesis in Populus in the first generation. Sci. Rep. 2015, 5, 12217. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Liu, X.J.; Zhang, Z.; Wang, Y.; Duan, W.; Li, S.H.; Liang, Z.C. CRISPR/Cas9-mediated efficient targeted mutagenesis in Chardonnay (Vitis vinifera L.). Sci. Rep. 2016, 6, 32289. [Google Scholar] [CrossRef] [PubMed]
- Li, F.G.; Fan, G.Y.; Lu, C.R.; Xiao, G.H.; Zou, C.S.; Kohel, R.J.; Ma, Z.Y.; Shang, H.H.; Ma, X.F.; Wu, J.Y.; et al. Genome sequence of cultivated upland cotton (Gossypium hirsutum TM-1) provides insights into genome evolution. Nat. Biotechnol. 2015, 33, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.Z.; Hu, Y.; Jiang, W.K.; Fang, L.; Guan, X.Y.; Chen, J.D.; Zhang, J.B.; Saski, C.A.; Scheffler, B.E.; Stelly, D.M.; et al. Sequencing of allotetraploid cotton (Gossypium hirsutum L. acc. TM-1) provides a resource for fiber improvement. Nat. Biotechnol. 2015, 33, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Janga, M.R.; Campbell, L.M.; Rathore, K.S. CRISPR/Cas9-mediated targeted mutagenesis in upland cotton (Gossypium hirsutum L.). Plant Mol. Biol. 2017, 94, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.C.; Zhang, J.; Sun, L.; Ma, Y.Z.; Xu, J.; Liang, S.J.; Deng, J.W.; Tan, J.F.; Zhang, Q.H.; Tu, L.L.; et al. High efficient multi-sites genome editing in allotetraploid cotton (Gossypium hirsutum) using CRISPR/Cas9 system. Plant Biotechnol. J. 2018, 16, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.G.; Lu, X.K.; Shu, N.; Wang, S.; Wang, J.J.; Wang, D.L.; Guo, L.X.; Ye, W.W. Targeted mutagenesis in cotton (Gossypium hirsutum L.) using the CRISPR/Cas9 system. Sci. Rep. 2017, 7, 44304. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Long, L.; Tian, X.Q.; Xu, F.C.; Liu, J.; Singh, P.K.; Botella, J.R.; Song, C.P. Genome editing in cotton with the CRISPR/Cas9 system. Front. Plant Sci. 2017, 8, 1364. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Unver, T.; Zhang, B.H. A high-efficiency CRISPR/Cas9 system for targeted mutagenesis in cotton (Gossypium hirsutum L.). Sci. Rep. 2017, 7, 43902. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.L.; Meng, Z.G.; Liang, C.Z.; Meng, Z.H.; Wang, Y.; Sun, G.Q.; Zhu, T.; Cai, Y.P.; Guo, S.D.; Zhang, R.; et al. Increased lateral root formation by CRISPR/Cas9-mediated editing of arginase genes in cotton. Sci. China Life Sci. 2017, 60, 524–527. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.S.; Shen, B.; Zhang, C.B.; Huang, X.X.; Zhang, Y.L. sgRNAcas9: A software package for designing CRISPR sgRNA and evaluating potential off-target cleavage sites. PLoS ONE 2014, 9, e100448. [Google Scholar] [CrossRef] [PubMed]
- Bortesi, L.; Fischer, R. The CRISPR/Cas9 system for plant genome editing and beyond. Biotechnol. Adv. 2015, 33, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Lowder, L.G.; Zhang, D.W.; Baltes, N.J.; Paul, J.W.; Tang, X.; Zheng, X.L.; Voytas, D.F.; Hsieh, T.; Zhang, Y.; Qi, Y.P. A CRISPR/Cas9 toolbox for multiplexed plant genome editing and transcriptional regulation. Plant Physiol. 2015, 169, 971–985. [Google Scholar] [CrossRef] [PubMed]
- Seth, K.; Harish. Current status of potential applications of repurposed Cas9 for structural and functional genomics of plants. Biochem. Biophys. Res. Commun. 2016, 480, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Baysal, C.; Bortesi, L.; Zhu, C.F.; Farre, G.; Schillberg, S.; Christou, P. CRISPR/Cas9 activity in the rice OsBEIIb gene does not induce off-target effects in the closely related paralog OsBEIIa. Mol. Breed. 2016, 36, 108. [Google Scholar] [CrossRef]
- Wang, W.; Akhunova, A.R.; Chao, S.M.; Akhunov, E. Optimizing multiplex CRISPR/Cas9-based genome editing for wheat. BioRxiv 2016. [Google Scholar] [CrossRef]
- Zhang, B.H. Transgenic cotton: From biotransformation methods to agricultural application. Methods Mol. Biol. 2013, 985, 3–15. [Google Scholar]
- Shan, Q.W.; Wang, Y.P.; Li, J.; Gao, C.C. Genome editing in rice and wheat using the CRISPR/Cas system. Nat. Protoc. 2014, 9, 2395–2410. [Google Scholar] [CrossRef] [PubMed]
- Xing, H.; Dong, L.; Wang, Z.P.; Zhang, H.Y.; Han, C.Y.; Liu, B.; Wang, X.C.; Chen, Q.J. A CRISPR/Cas9 toolkit for multiplex genome editing in plants. BMC Plant Biol. 2014, 14, 327. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.L.; Zhang, Q.Y.; Zhu, Q.L.; Liu, W.; Chen, Y.; Qiu, R.; Wang, B.; Yang, Z.F.; Li, H.Y.; Lin, Y.; et al. A robust CRISPR/Cas9 system for convenient, high-efficiency multiplex genome editing in monocot and dicot Plants. Mol. Plant 2015, 8, 1274–1284. [Google Scholar] [CrossRef] [PubMed]
- Kusaba, M. RNA interference in crop plants. Curr. Opin. Biotechnol. 2004, 15, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Tuttle, J.R.; Idris, A.M.; Brown, J.K.; Haigler, C.H.; Robertson, D. Geminivirus-mediated gene silencing from cotton leaf crumple virus is enhanced by low temperature in cotton. Plant Physiol. 2008, 148, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.X.; Zhang, X.L.; Nie, Y.C.; Guo, X.P.; Liang, S.G.; Zhu, H. Identification of a novel elite genotype for in vitro culture and genetic transformation of cotton. Biol. Plant. 2006, 50, 519–524. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Gene | Rate of Different Nucleotide Insertions (+) and Deletions (−) (%) | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | +1 | −1 | −2 | −4 | −6 | −7 | −10 | −11 | −13 | −16 | −21 | −42 | −44 | −45 | −55 | −21/+3 | −26/+1 | −38/+1 | −6/+18 | −13/+99 | ||
| L1 | GhALARP-A | 61.5 | 30.8 | 7.7 | ||||||||||||||||||
| GhALARP-D | 57.1 | 28.6 | 14.3 | |||||||||||||||||||
| L2 | GhALARP-A | 50 | 16.7 | 33.3 | ||||||||||||||||||
| GhALARP-D | 64.3 | 14.3 | 21.4 | |||||||||||||||||||
| L3 | GhALARP-A | 11.1 | 66.7 | 22.2 | ||||||||||||||||||
| GhALARP-D | 18.2 | 27.3 | 9.1 | 9.1 | 9.1 | 9.1 | 9.1 | 9.1 | ||||||||||||||
| L4 | GhALARP-A | 75 | 25 | |||||||||||||||||||
| GhALARP-D | 83.3 | 16.7 | ||||||||||||||||||||
| L5 | GhALARP-A | 50 | 25 | 25 | ||||||||||||||||||
| GhALARP-D | 91.7 | 8.3 | ||||||||||||||||||||
| L6 | GhALARP-A | 16.7 | 33.3 | 25 | 8.3 | 16.7 | ||||||||||||||||
| GhALARP-D | 12.5 | 37.5 | 12.5 | 25 | 12.5 | |||||||||||||||||
| L7 | GhALARP-A | 28.6 | 14.3 | 42.9 | 14.3 | |||||||||||||||||
| GhALARP-D | 15.4 | 53.8 | 30.8 | |||||||||||||||||||
| L8 | GhALARP-A | 33.3 | 16.7 | 50 | ||||||||||||||||||
| GhALARP-D | 7.1 | 57.1 | 21.4 | 14.3 | ||||||||||||||||||
| L9 | GhALARP-A | 11.1 | 33.3 | 55.6 | ||||||||||||||||||
| GhALARP-D | 18.2 | 72.7 | 9.1 | |||||||||||||||||||
| L10 | GhALARP-A | 33.3 | 55.6 | 11.1 | ||||||||||||||||||
| GhALARP-D | 81.8 | 9.1 | 9.1 | |||||||||||||||||||
| sgRNA Name | Putative Off-Target | No. of Mismatched Nucleotides | No. of Examined Events | No. of Off-Target Events |
|---|---|---|---|---|
| GhALARP-sgRNA | D02: −57231491 | 4 | 86 | 0 |
| GhALARP-sgRNA | A11: +20565332 | 4 | 61 | 0 |
| GhALARP-sgRNA | D02: +21785633 | 4 | 63 | 0 |
| GhALARP-sgRNA | D09: +7375255 | 4 | 74 | 0 |
| GhALARP-sgRNA | A10: −6696373 (Gh_A10G0542) | 5 | 100 | 0 |
| GhALARP-sgRNA | D08: +5937364 | 5 | 79 | 0 |
| GhALARP-sgRNA | D10: −36547470 | 4 | 82 | 0 |
| GhALARP-sgRNA | scaffold2307_A09: −61148 (Gh_A09G2459) | 4 | 100 | 0 |
| GhALARP-sgRNA | scaffold27980: +36 | 4 | 73 | 0 |
| GhALARP-sgRNA | A10: +36113181 | 4 | 66 | 0 |
| GhALARP-sgRNA | D04: +37760986 | 4 | 92 | 0 |
| GhALARP-sgRNA | D08: −59028209 | 4 | 65 | 0 |
| GhALARP-sgRNA | D08: −59042750 | 4 | 88 | 0 |
| GhALARP-sgRNA | scaffold2106_A08: −632710 | 5 | 83 | 0 |
| GhALARP-sgRNA | A11: −87746479 (Gh_A11G2627) | 5 | 100 | 0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, S.; Yu, X.; Li, Y.; Sun, Y.; Zhu, Q.; Sun, J. Highly Efficient Targeted Gene Editing in Upland Cotton Using the CRISPR/Cas9 System. Int. J. Mol. Sci. 2018, 19, 3000. https://doi.org/10.3390/ijms19103000
Zhu S, Yu X, Li Y, Sun Y, Zhu Q, Sun J. Highly Efficient Targeted Gene Editing in Upland Cotton Using the CRISPR/Cas9 System. International Journal of Molecular Sciences. 2018; 19(10):3000. https://doi.org/10.3390/ijms19103000
Chicago/Turabian StyleZhu, Shouhong, Xiuli Yu, Yanjun Li, Yuqiang Sun, Qianhao Zhu, and Jie Sun. 2018. "Highly Efficient Targeted Gene Editing in Upland Cotton Using the CRISPR/Cas9 System" International Journal of Molecular Sciences 19, no. 10: 3000. https://doi.org/10.3390/ijms19103000
APA StyleZhu, S., Yu, X., Li, Y., Sun, Y., Zhu, Q., & Sun, J. (2018). Highly Efficient Targeted Gene Editing in Upland Cotton Using the CRISPR/Cas9 System. International Journal of Molecular Sciences, 19(10), 3000. https://doi.org/10.3390/ijms19103000