Key Immunological Functions Involved in the Progression of Epithelial Ovarian Serous Carcinoma Discovered by the Gene Ontology-Based Immunofunctionome Analysis

 ,
,  ,
, 
 , ,
, ,
Abstract
:1. Introduction
2. Results
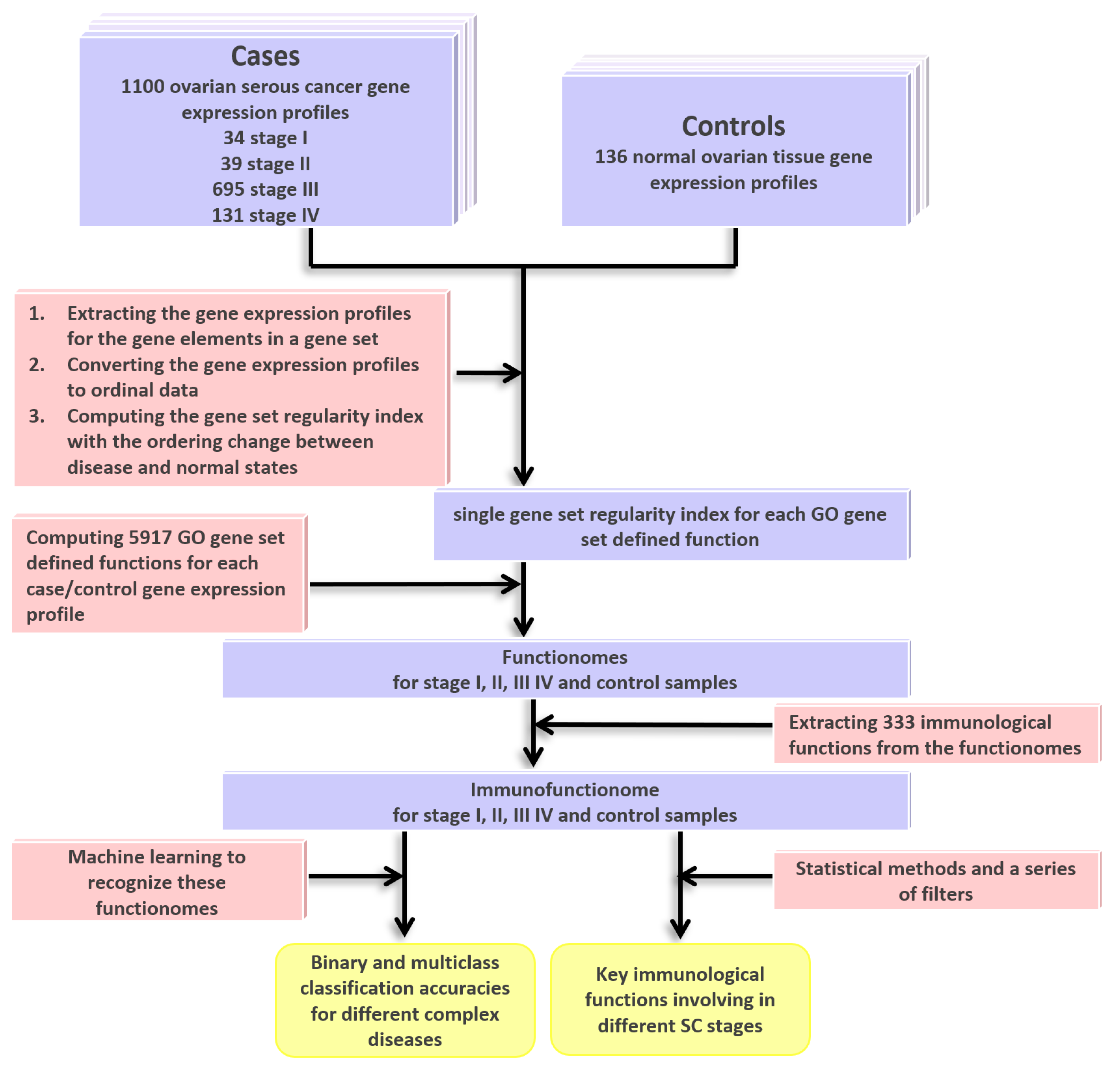
2.1. DNA Microarray Gene Expression Datasets for SC and Gene Ontology (GO) Gene Set Definition
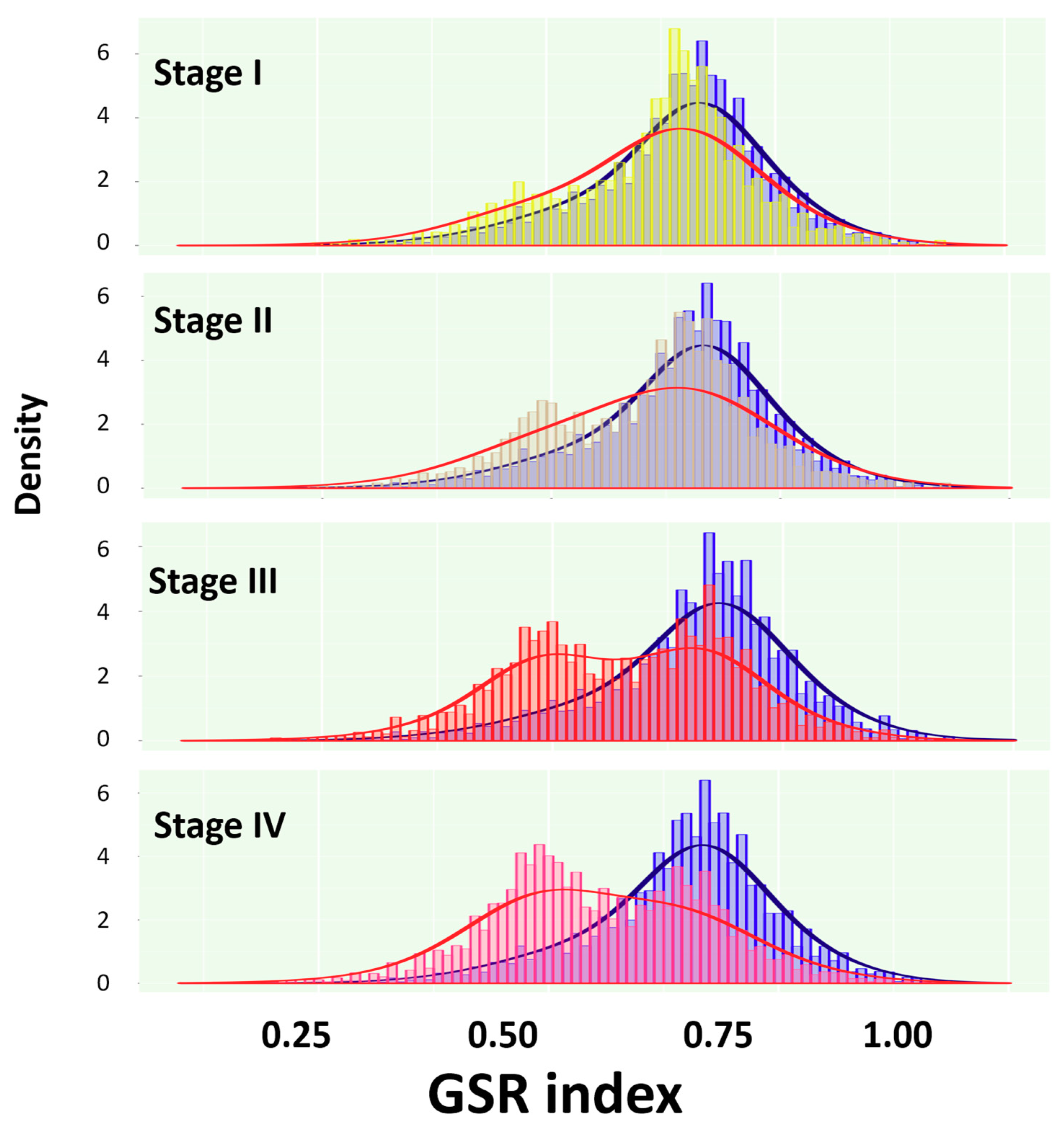
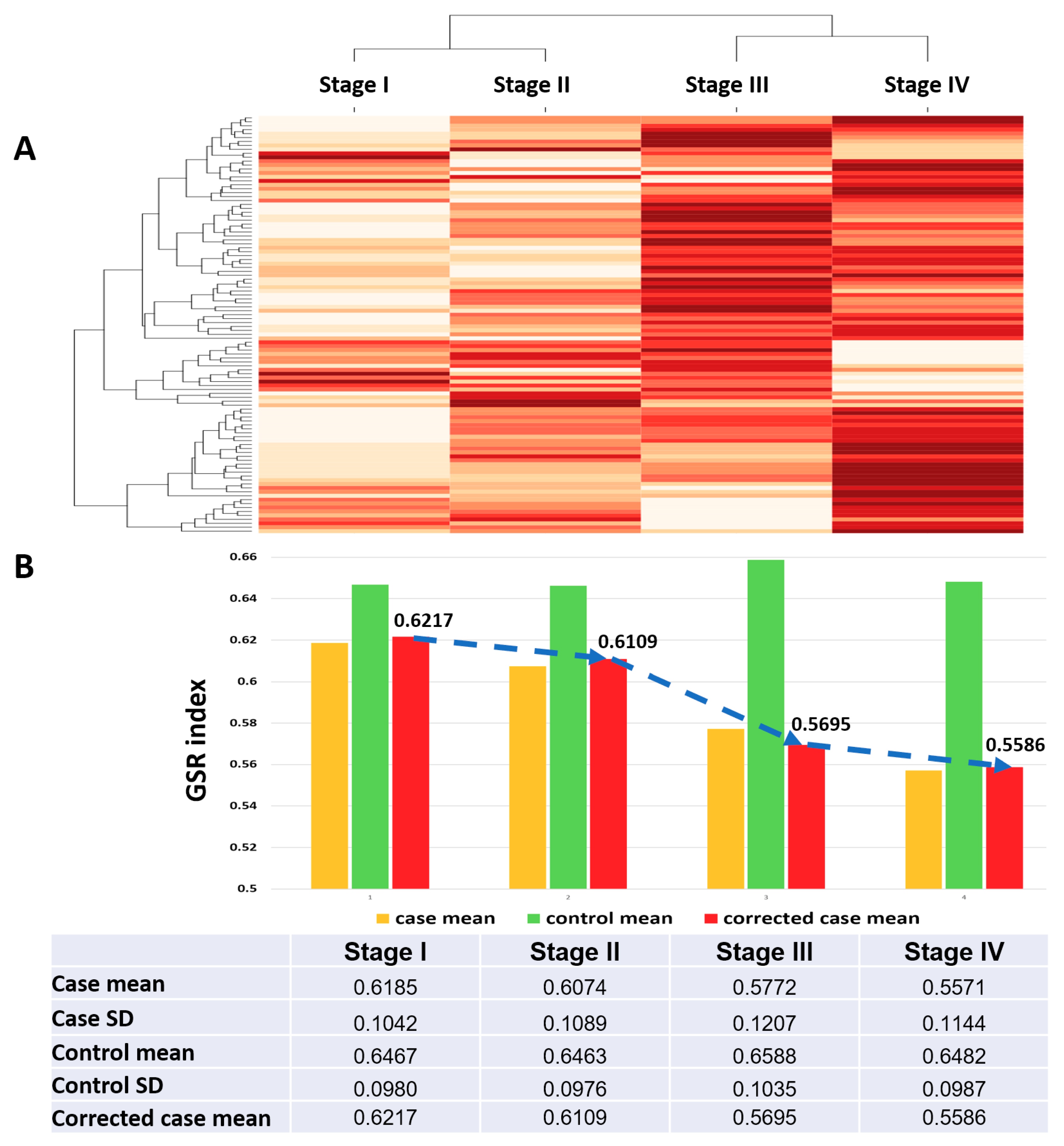
2.2. Reconstruction and Comparison of Functionomes Between the SC Groups and Normal Controls
2.3. Comparison of the Immunofunctionomes among the Four SC Staging Groups
2.4. The Global Function Regulation from Stage I to IV Shows Distinct Pattern That Can Be Correctly Classified and Predicted by Machine Learning
2.5. The Most Significantly Deregulated Immunological Fucntions for the Four SC Staging Groups
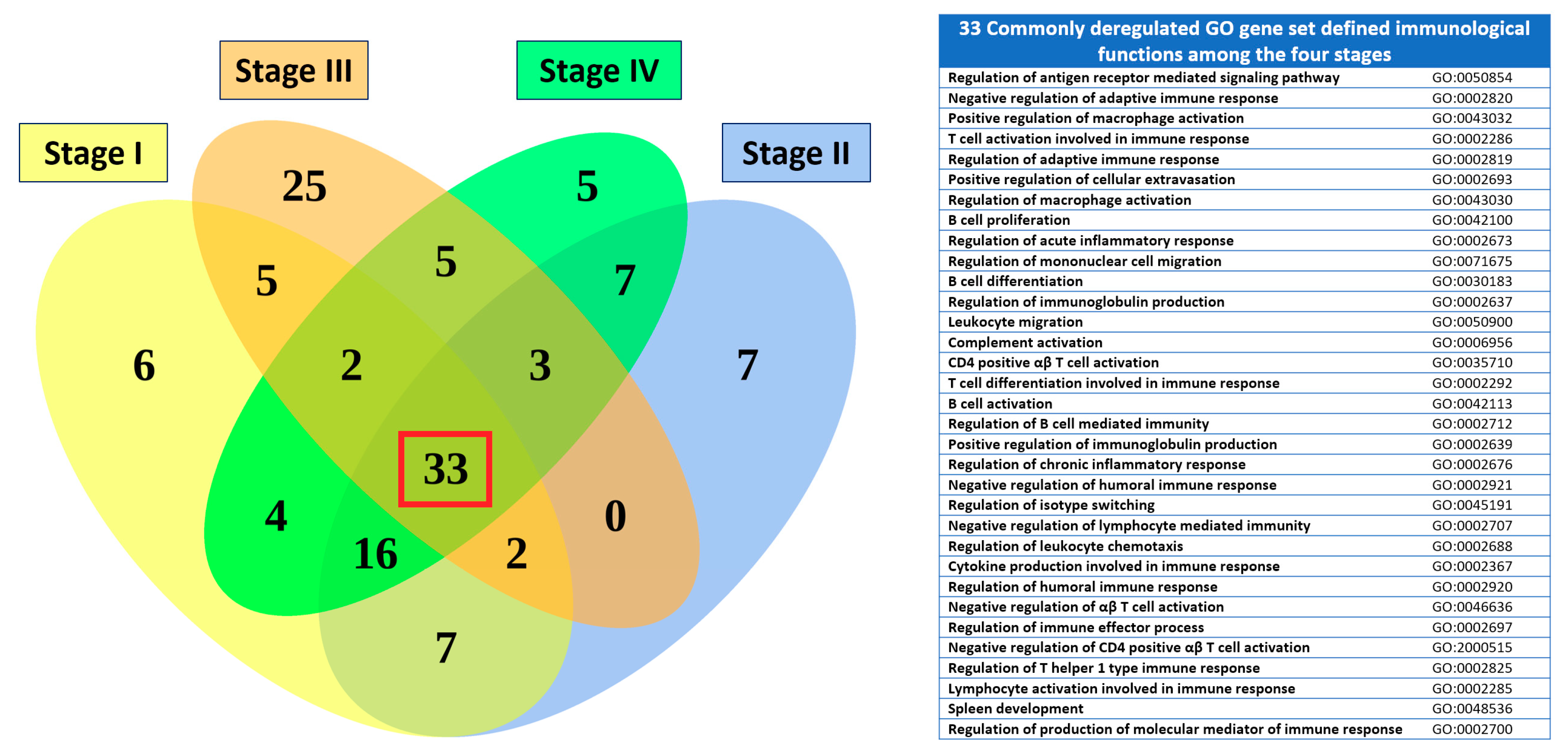
2.6. The Commonly Deregulated Immunological Functions among the Four Staging Groups
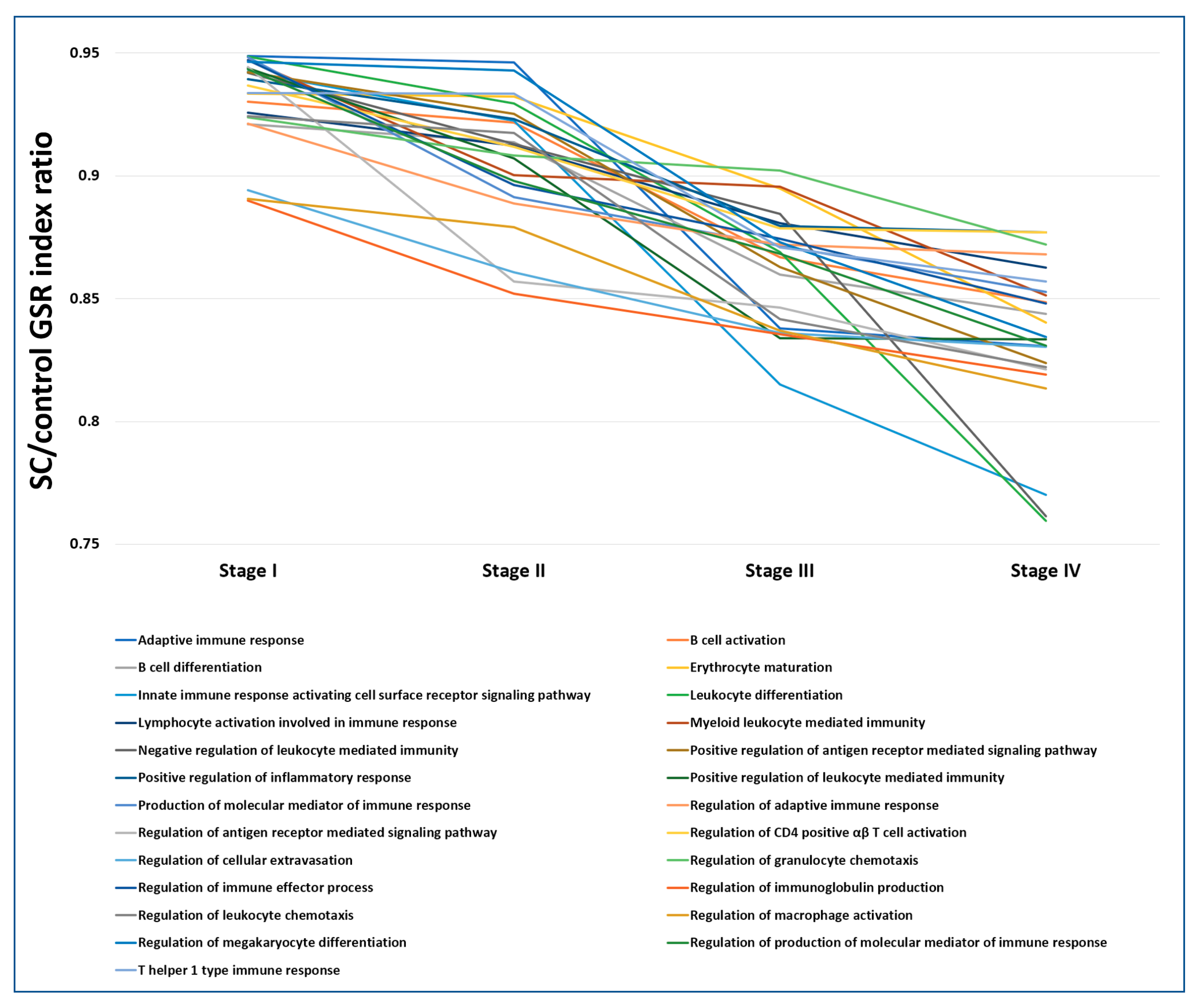
2.7. The Progressively Deregulated Immunological Functions in the Pathogenesis of SC from Stage I to IV
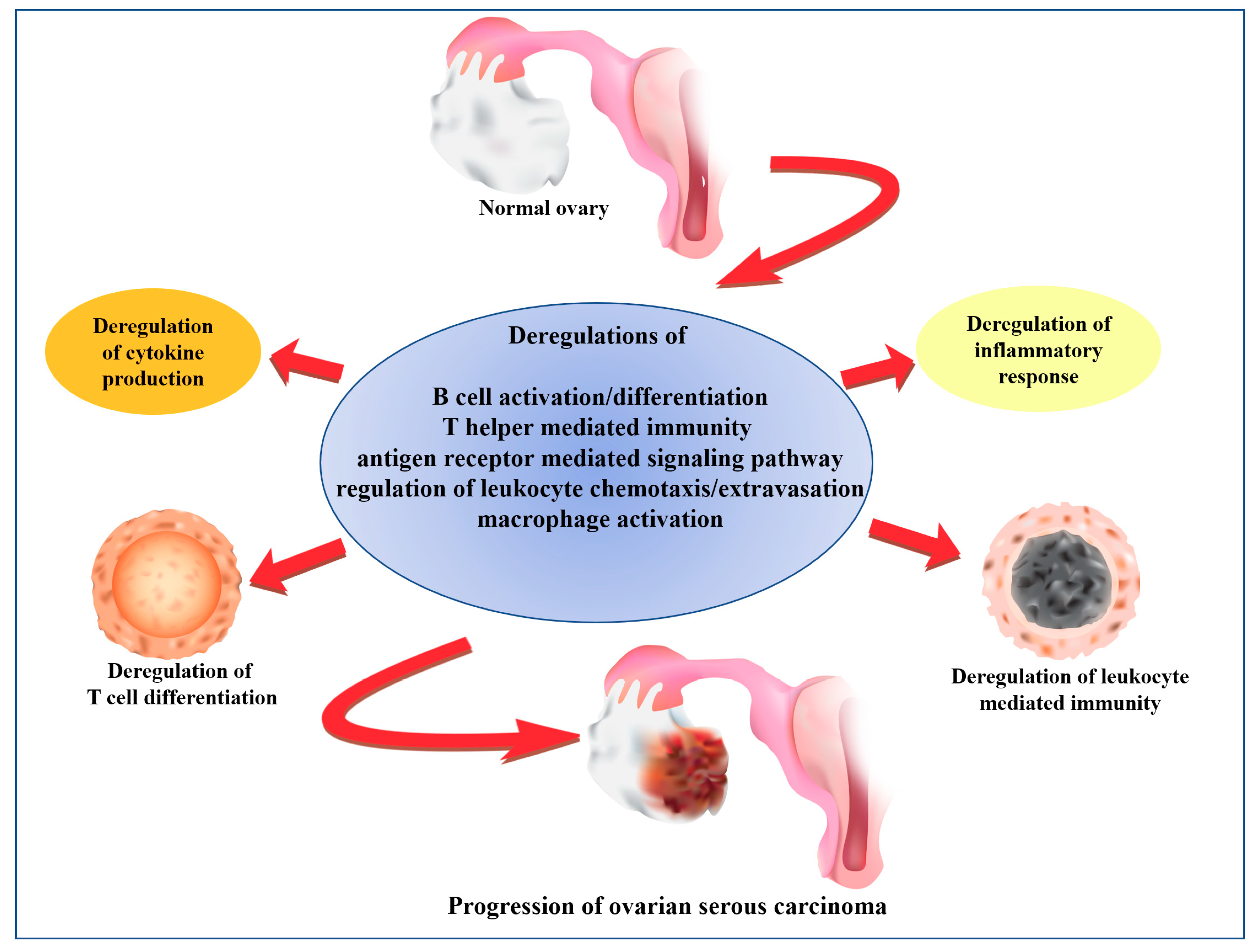
2.8. The Core and Auxiliary Elements of Deregulated Immunological Functions Involved in the Progression of SC
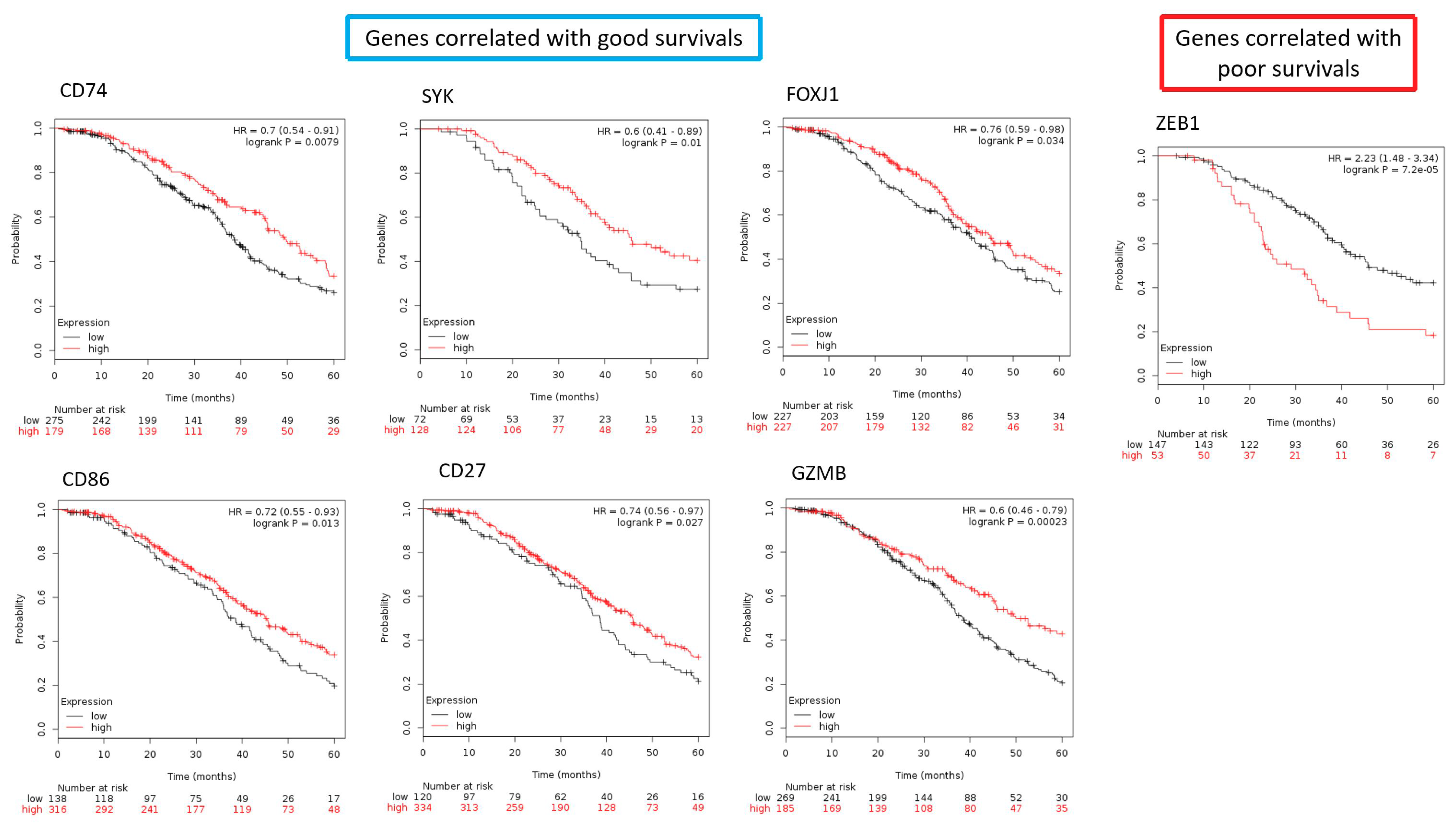
2.9. The Differentially Expressed Genes in the Core Elements of Deregulated Immunological Functions Involved in the Progression of SC
3. Discussion
4. Materials and Methods
4.1. Computing the GSR Indices and Reconstruction of Functionome and Immunofunctionome
4.2. Microarray Datasets Collection
4.3. Statistical Analysis
4.4. Classification and Prediction by Machine Learning
4.5. Set Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| SC | Serous carcinoma |
| GO | Gene ontology |
| EOC | Epithelial ovarian cancer |
| FIGO | Federation of gynecologists and obstetrics |
| ERBB | Erb-B2 Receptor Tyrosine Kinase |
| PI3K | Phosphoinositide 3-kinase |
| EAOC | endometriosis-associated ovarian carcinoma |
| GEO | Gene expression omnibus |
| MSigDB | Molecular signatures database |
| GSR | Gene set regularity |
| SD | Standard deviation |
| SVM | Support vector machine |
| AUC | Area under curve |
| CD74 | CD74 Molecule |
| NK | Natural killer |
| WNT5A | Wnt family member 5A |
| NR1H3 | Nuclear receptor subfamily 1 group H member 3 |
| STAP1 | Signal transducing adaptor family member 1 |
| RORA | RAR related orphan receptor A |
| ZC3H12A | Zinc finger CCCH-type containing 12A |
| PLA2G10 | Phospholipase A2 group X |
| IL33 | Interleukin 33 |
| CDKN2A | Cyclin dependent kinase inhibitor 2A |
| SYK | Spleen associated tyrosine kinase |
| MYB | MYB proto-oncogene, transcription factor |
| FOXJ1 | Forkhead box J1 |
| ZEB1 | Zinc finger E-box binding homeobox 1 |
| CD74 | CD74 molecule |
| LGALS9 | Galectin 9 |
| ADAM8 | ADAM metallopeptidase domain 8 |
| GLI2 | GLI family zinc finger 2 |
| CD86 | CD86 molecule |
| PRKCD | Protein kinase C delta |
| CORO1A | Coronin 1A |
| PTPN6 | Protein tyrosine phosphatase, non-receptor type 6 |
| MSH2 | MutS homolog 2 |
| EXO1 | Exonuclease 1 |
| SLC11A1 | Solute carrier family 11 member 1 |
| CD27 | CD27 molecule |
| GATA3 | GATA binding protein 3 |
| GZMB | Granzyme B |
| IL2 | Interleukin 2 |
| TAA | Tumor-associated antigens |
| MHC | histocompatibility complex |
| TIL | tumor infiltrating lymphocytes |
| CD3 | cluster of differentiation 3 |
| CD8 | cluster of differentiation 8 |
| IL4 | interleukin 4 |
| IL13 | interleukin 13 |
| IL10 | interleukin 10 |
| M-CSF | macrophage colony-stimulating factor |
| GM-CSF | Granulocyte-macrophage colony-stimulating factor |
| IFN-γ | Interferon γ |
| TLR | Toll-like receptor |
| IL8 | Interleukin 8 |
| Th1 | T helper 1 |
| Th2 | T helper 2 |
| IL12 | interleukin 12 |
| TNF-α | Tumor Necrosis Factor |
| Treg | regulatory T cell |
| DIRAC | Differential rank conservation |
| TCGA | The cancer genome atlas |
References
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Benedet, J.L.; Bender, H.; Jones, H.; Ngan, H.Y.; Pecorelli, S. Figo staging classifications and clinical practice guidelines in the management of gynecologic cancers. Figo committee on gynecologic oncology. Int. J. Gynaecol. Obstet. 2000, 70, 209–262. [Google Scholar] [PubMed]
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Conejo-Garcia, J.R.; Katsaros, D.; Gimotty, P.A.; Massobrio, M.; Regnani, G.; Makrigiannakis, A.; Gray, H.; Schlienger, K.; Liebman, M.N.; et al. Intratumoral t cells, recurrence, and survival in epithelial ovarian cancer. N. Engl. J. Med. 2003, 348, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.M.; Chuang, C.M.; Wang, M.L.; Yang, M.J.; Chang, C.C.; Yen, M.S.; Chiou, S.H. Gene set-based functionome analysis of pathogenesis in epithelial ovarian serous carcinoma and the molecular features in different figo stages. Int. J. Mol. Sci. 2016, 17, 886. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.M.; Chuang, C.M.; Wang, M.L.; Yang, Y.P.; Chuang, J.H.; Yang, M.J.; Yen, M.S.; Chiou, S.H.; Chang, C.C. Gene set-based integrative analysis revealing two distinct functional regulation patterns in four common subtypes of epithelial ovarian cancer. Int. J. Mol. Sci. 2016, 17, 1272. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.M.; Yang, Y.P.; Chuang, J.H.; Chuang, C.M.; Lin, T.W.; Wang, P.H.; Yu, M.H.; Chang, C.C. Discovering the deregulated molecular functions involved in malignant transformation of endometriosis to endometriosis-associated ovarian carcinoma using a data-driven, function-based analysis. Int. J. Mol. Sci. 2017, 18, E2345. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.M.; Wang, M.L.; Lu, K.H.; Yang, Y.P.; Juang, C.M.; Wang, P.H.; Hsu, R.J.; Yu, M.H.; Chang, C.C. Integrating the dysregulated inflammasome-based molecular functionome in the malignant transformation of endometriosis-associated ovarian carcinoma. Oncotarget 2018, 9, 3704–3726. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y. Cancer immunotherapy: Harnessing the immune system to battle cancer. J. Clin. Investig. 2015, 125, 3335–3337. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breuer, K.; Foroushani, A.K.; Laird, M.R.; Chen, C.; Sribnaia, A.; Lo, R.; Winsor, G.L.; Hancock, R.E.; Brinkman, F.S.; Lynn, D.J. Innatedb: Systems biology of innate immunity and beyond—Recent updates and continuing curation. Nucleic Acids Res. 2013, 41, D1228–D1233. [Google Scholar] [CrossRef] [PubMed]
- Gyorffy, B.; Lanczky, A.; Szallasi, Z. Implementing an online tool for genome-wide validation of survival-associated biomarkers in ovarian-cancer using microarray data from 1287 patients. Endocr. Relat. Cancer 2012, 19, 197–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gavalas, N.G.; Karadimou, A.; Dimopoulos, M.A.; Bamias, A. Immune response in ovarian cancer: How is the immune system involved in prognosis and therapy: Potential for treatment utilization. Clin. Dev. Immunol. 2010, 2010, 791603. [Google Scholar] [CrossRef] [PubMed]
- Spurrell, E.L.; Lockley, M. Adaptive immunity in cancer immunology and therapeutics. Ecancermedicalscience 2014, 8, 441. [Google Scholar] [PubMed]
- Tomsova, M.; Melichar, B.; Sedlakova, I.; Steiner, I. Prognostic significance of CD3+ tumor-infiltrating lymphocytes in ovarian carcinoma. Gynecol. Oncol. 2008, 108, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Sozzani, S.; Locati, M.; Allavena, P.; Sica, A. Macrophage polarization: Tumor-associated macrophages as a paradigm for polarized m2 mononuclear phagocytes. Trends Immunol. 2002, 23, 549–555. [Google Scholar] [CrossRef]
- Guo, C.; Buranych, A.; Sarkar, D.; Fisher, P.B.; Wang, X.Y. The role of tumor-associated macrophages in tumor vascularization. Vasc. Cell 2013, 5, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knutson, K.L.; Disis, M.L. Tumor antigen-specific t helper cells in cancer immunity and immunotherapy. Cancer Immunol. Immunother. 2005, 54, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Whiteside, T.L. Immune responses to malignancies. J. Allergy Clin. Immunol. 2010, 125, S272–S283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gajewski, T.F.; Schreiber, H.; Fu, Y.X. Innate and adaptive immune cells in the tumor microenvironment. Nat. Immunol. 2013, 14, 1014–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssen, L.M.E.; Ramsay, E.E.; Logsdon, C.D.; Overwijk, W.W. The immune system in cancer metastasis: Friend or foe? J. Immunother. Cancer 2017, 5, 79. [Google Scholar] [CrossRef] [PubMed]
- Gavalas, N.; Dimopoulos, M.A.; Bamias, A. Anti-tumor immune response in ovarian cancer: Clinical implications, prognostic significance and potential for novel treatment strategies. Oncol. Rev. 2011, 3, 173–186. [Google Scholar] [CrossRef]
- Sarvaria, A.; Madrigal, J.A.; Saudemont, A. B cell regulation in cancer and anti-tumor immunity. Cell Mol. Immunol. 2017, 14, 662–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Zhang, C.; Zhang, J.; Tian, Z. Macrophages help Nk cells to attack tumor cells by stimulatory nkg2d ligand but protect themselves from nk killing by inhibitory ligand qa-1. PLoS ONE 2012, 7, e36928. [Google Scholar] [CrossRef] [PubMed]
- Sakata, J.; Kajiyama, H.; Suzuki, S.; Utsumi, F.; Niimi, K.; Sekiya, R.; Shibata, K.; Senga, T.; Kikkawa, F. Impact of positive zeb1 expression in patients with epithelial ovarian carcinoma as an oncologic outcome-predicting indicator. Oncol. Lett. 2017, 14, 4287–4293. [Google Scholar] [CrossRef] [PubMed]
- Koury, J.; Lucero, M.; Cato, C.; Chang, L.; Geiger, J.; Henry, D.; Hernandez, J.; Hung, F.; Kaur, P.; Teskey, G.; et al. Immunotherapies: Exploiting the immune system for cancer treatment. J. Immunol. Res. 2018, 2018, 9585614. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.L.; Jin, L.P. Immune cell population in ovarian tumor microenvironment. J. Cancer 2017, 8, 2915–2923. [Google Scholar] [CrossRef] [PubMed]
- Eddy, J.A.; Hood, L.; Price, N.D.; Geman, D. Identifying tightly regulated and variably expressed networks by differential rank conservation (dirac). PLoS Comput. Biol. 2010, 6, e1000792. [Google Scholar] [CrossRef] [PubMed]
- Robin, X.; Turck, N.; Hainard, A.; Tiberti, N.; Lisacek, F.; Sanchez, J.C.; Muller, M. Proc: An open-source package for r and s+ to analyze and compare roc curves. BMC Bioinform. 2011, 12, 77. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | Case | Control | Total | Case Mean (SD) | Control Mean (SD) | Corrected Case Mean | p Value * |
|---|---|---|---|---|---|---|---|
| I | 34 | 136 | 170 | 0.6195 (0.1035) | 0.6461 (0.1018) | 0.6214 | <0.05 |
| II | 39 | 136 | 175 | 0.6021 (0.1109) | 0.6459 (0.1017) | 0.6041 | <0.05 |
| III | 695 | 136 | 831 | 0.5748 (0.1205) | 0.6518 (0.1083) | 0.5715 | <0.05 |
| IV | 131 | 136 | 267 | 0.5588 (0.1154) | 0.6486 (0.1031) | 0.5583 | <0.05 |
| Binary Classification | K = | Sensitivity (Mean) | Sensitivity (SD) | Specificity (Mean) | Specificity (SD) | Accuracy (Mean) | Accuracy (SD) | AUC |
|---|---|---|---|---|---|---|---|---|
| stage I | 5 | 1.0000 | 0.0000 | 1.0000 | 0.0000 | 1.0000 | 0.0000 | 1.0000 |
| 3 | 0.9851 | 0.0313 | 1.0000 | 0.0000 | 0.9964 | 0.0073 | 0.9911 | |
| 2 | 0.9933 | 0.0210 | 1.0000 | 0.0000 | 0.9988 | 0.0037 | 0.9969 | |
| stage II | 5 | 0.9334 | 0.0839 | 1.0000 | 0.0000 | 0.9857 | 0.0150 | 0.9702 |
| 3 | 0.9607 | 0.0422 | 1.0000 | 0.0000 | 0.9913 | 0.0090 | 0.9803 | |
| 2 | 0.9711 | 0.0249 | 1.0000 | 0.0000 | 0.9931 | 0.0059 | 0.9852 | |
| stage III | 5 | 1.0000 | 0.0000 | 1.0000 | 0.0000 | 1.0000 | 0.0000 | 1.0000 |
| 3 | 1.0000 | 0.0000 | 0.9977 | 0.0071 | 0.9996 | 0.0011 | 0.9988 | |
| 2 | 0.9997 | 0.0009 | 0.9917 | 0.0148 | 0.9983 | 0.0025 | 0.9955 | |
| stage IV | 5 | 1.0000 | 0.0000 | 1.0000 | 0.0000 | 1.0000 | 0.0000 | 1.0000 |
| 3 | 0.9958 | 0.0087 | 0.9976 | 0.0073 | 0.9966 | 0.0054 | 0.9966 | |
| 2 | 0.9938 | 0.0103 | 0.9969 | 0.0097 | 0.9954 | 0.0063 | 0.9954 | |
| Multiclass classification | 5 | 0.9244 | 0.0227 | 1.0000 | 0.0000 | 0.9338 | 0.0195 | 0.9881 |
| Stage I | p Value | GO Index | |
| 1 | Positive regulation of B cell mediated immunity | 1.2874 × 10−14 | GO:0002714 |
| 2 | T cell differentiation involved in immune response | 1.7204 × 10−14 | GO:0002292 |
| 3 | Regulation of B cell mediated immunity | 3.5164 × 10−14 | GO:0002712 |
| 4 | Cytokine production involved in immune response | 3.5232 × 10−14 | GO:0002367 |
| 5 | Regulation of isotype switching | 2.4920 × 10−12 | GO:0045191 |
| 6 | Negative regulation of CD4 positive αβ T cell activation | 4.4483 × 10−12 | GO:2000515 |
| 7 | Negative regulation of αβ T cell differentiation | 5.4946 × 10−11 | GO:0046639 |
| 8 | Regulation of lymphocyte chemotaxis | 9.0873 × 10−11 | GO:1901623 |
| 9 | Positive regulation of immunoglobulin production | 3.1226 × 10−10 | GO:0002639 |
| 10 | Negative regulation of αβ T cell activation | 8.6150 × 10−10 | GO:0046636 |
| 11 | Positive regulation of activated T cell proliferation | 1.2196 × 10−09 | GO:0042104 |
| 12 | Negative regulation of adaptive immune response | 2.2585 × 10−09 | GO:0002820 |
| 13 | Positive regulation of adaptive immune response | 3.5157 × 10−09 | GO:0002821 |
| 14 | Regulation of immunoglobulin production | 3.7150 × 10−09 | GO:0002637 |
| 15 | Regulation of adaptive immune response | 4.6564 × 10−09 | GO:0002819 |
| 16 | T cell activation involved in immune response | 6.1231 × 10−09 | GO:0002286 |
| 17 | Regulation of macrophage activation | 1.0046 × 10−08 | GO:0043030 |
| 18 | Regulation of lymphocyte mediated immunity | 1.7703 × 10−08 | GO:0002706 |
| 19 | Regulation of activated T cell proliferation | 1.8826 × 10−08 | GO:0046006 |
| 20 | Positive regulation of lymphocyte mediated immunity | 6.7086 × 10−08 | GO:0002708 |
| Stage II | |||
| 1 | Regulation of B cell mediated immunity | 7.3615 × 10−16 | GO:0002712 |
| 2 | Positive regulation of B cell mediated immunity | 8.6942 × 10−16 | GO:0002714 |
| 3 | Regulation of isotype switching | 1.6517 × 10−14 | GO:0045191 |
| 4 | Cytokine production involved in immune response | 2.7748 × 10−13 | GO:0002367 |
| 5 | T cell differentiation involved in immune response | 1.8020 × 10−11 | GO:0002292 |
| 6 | Negative regulation of CD4 positive αβ T cell activation | 6.5398 × 10−11 | GO:2000515 |
| 7 | Positive regulation of immunoglobulin production | 6.5398 × 10−11 | GO:0002639 |
| 8 | Regulation of immunoglobulin production | 7.4799 × 10−11 | GO:0002637 |
| 9 | Regulation of adaptive immune response | 1.0861 × 10−10 | GO:0002819 |
| 10 | Negative regulation of adaptive immune response | 1.3364 × 10−10 | GO:0002820 |
| 11 | Positive regulation of adaptive immune response | 1.9411 × 10−10 | GO:0002821 |
| 12 | Regulation of lymphocyte chemotaxis | 1.9411 × 10−10 | GO:1901623 |
| 13 | Regulation of lymphocyte mediated immunity | 1.2403 × 10−09 | GO:0002706 |
| 14 | Negative regulation of αβ T cell activation | 7.1847 × 10−09 | GO:0046636 |
| 15 | Positive regulation of activated T cell proliferation | 8.7581 × 10−09 | GO:0042104 |
| 16 | Positive regulation of lymphocyte mediated immunity | 8.7581 × 10−09 | GO:0002708 |
| 17 | Regulation of humoral immune response | 8.7581 × 10−09 | GO:0002920 |
| 18 | Positive regulation of macrophage activation | 1.0451 × 10−08 | GO:0043032 |
| 19 | Negative regulation of humoral immune response | 1.8493 × 10−08 | GO:0002921 |
| 20 | Regulation of macrophage activation | 1.8493 × 10−08 | GO:0043030 |
| Stage III | |||
| 1 | Negative regulation of CD4 positive αβ T cell activation | 1.3948 × 10−62 | GO:2000515 |
| 2 | Negative regulation of αβ T cell activation | 3.3724 × 10−54 | GO:0046636 |
| 3 | Negative regulation of adaptive immune response | 7.1266 × 10−49 | GO:0002820 |
| 4 | Erythrocyte homeostasis | 1.0745 × 10−46 | GO:0034101 |
| 5 | Myeloid cell homeostasis | 2.2732 × 10−45 | GO:0002262 |
| 6 | T cell differentiation involved in immune response | 1.3835 × 10−43 | GO:0002292 |
| 7 | MyD88 dependent Toll like receptor signaling pathway | 2.4472 × 10−43 | GO:0002755 |
| 8 | Regulation of humoral immune response | 2.3395 × 10−41 | GO:0002920 |
| 9 | B cell proliferation | 3.3633 × 10−41 | GO:0042100 |
| 10 | Regulation of CD4 positive αβ T cell activation | 1.4256 × 10−40 | GO:2000514 |
| 11 | T cell activation involved in immune response | 7.6933 × 10−40 | GO:0002286 |
| 12 | Lymphocyte activation involved in immune response | 1.0932 × 10−39 | GO:0002285 |
| 13 | Regulation of T helper 1 type immune response | 1.2770 × 10−39 | GO:0002825 |
| 14 | Natural killer cell activation involved in immune response | 3.0394 × 10−39 | GO:0002323 |
| 15 | Regulation of αβ T cell proliferation | 3.0394 × 10−39 | GO:0046640 |
| 16 | Dendritic cell differentiation | 4.1689 × 10−39 | GO:0097028 |
| 17 | Regulation of lymphocyte chemotaxis | 4.1689 × 10−39 | GO:1901623 |
| 18 | Erythrocyte development | 6.5301 × 10−39 | GO:0048821 |
| 19 | Negative regulation of lymphocyte mediated immunity | 1.5062 × 10−38 | GO:0002707 |
| 20 | Thymic T cell selection | 4.8521 × 10−38 | GO:0045061 |
| Stage IV | |||
| 1 | Regulation of B cell mediated immunity | 4.2582 × 10−36 | GO:0002712 |
| 2 | Positive regulation of B cell mediated immunity | 1.3571 × 10−33 | GO:0002714 |
| 3 | Regulation of isotype switching | 1.1468 × 10−32 | GO:0045191 |
| 4 | Positive regulation of immunoglobulin production | 2.3588 × 10−29 | GO:0002639 |
| 5 | Regulation of immunoglobulin production | 2.3588 × 10−29 | GO:0002637 |
| 6 | Negative regulation of adaptive immune response | 5.4043 × 10−29 | GO:0002820 |
| 7 | Regulation of adaptive immune response | 1.4043 × 10−28 | GO:0002819 |
| 8 | T cell differentiation involved in immune response | 2.3196 × 10−28 | GO:0002292 |
| 9 | Cytokine production involved in immune response | 2.4258 × 10−28 | GO:0002367 |
| 10 | Regulation of humoral immune response | 2.4258 × 10−28 | GO:0002920 |
| 11 | Negative regulation of CD4 positive αβ T cell activation | 8.1639 × 10−28 | GO:2000515 |
| 12 | Regulation of lymphocyte mediated immunity | 2.6040 × 10−27 | GO:0002706 |
| 13 | Negative regulation of lymphocyte mediated immunity | 8.0954 × 10−26 | GO:0002707 |
| 14 | Positive regulation of adaptive immune response | 8.0954 × 10−26 | GO:0002821 |
| 15 | Regulation of acute inflammatory response | 8.0954 × 10−26 | GO:0002673 |
| 16 | Regulation of T helper 1 type immune response | 1.3689 × 10−25 | GO:0002825 |
| 17 | Regulation of macrophage activation | 2.7349 × 10−25 | GO:0043030 |
| 18 | Negative regulation of αβ T cell activation | 3.6163 × 10−25 | GO:0046636 |
| 19 | Regulation of immune effector process | 1.6392 × 10−24 | GO:0002697 |
| 20 | Positive regulation of lymphocyte mediated immunity | 1.7601 × 10−24 | GO:0002708 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, C.-C.; Su, K.-M.; Lu, K.-H.; Lin, C.-K.; Wang, P.-H.; Li, H.-Y.; Wang, M.-L.; Lin, C.-K.; Yu, M.-H.; Chang, C.-M. Key Immunological Functions Involved in the Progression of Epithelial Ovarian Serous Carcinoma Discovered by the Gene Ontology-Based Immunofunctionome Analysis. Int. J. Mol. Sci. 2018, 19, 3311. https://doi.org/10.3390/ijms19113311
Chang C-C, Su K-M, Lu K-H, Lin C-K, Wang P-H, Li H-Y, Wang M-L, Lin C-K, Yu M-H, Chang C-M. Key Immunological Functions Involved in the Progression of Epithelial Ovarian Serous Carcinoma Discovered by the Gene Ontology-Based Immunofunctionome Analysis. International Journal of Molecular Sciences. 2018; 19(11):3311. https://doi.org/10.3390/ijms19113311
Chicago/Turabian StyleChang, Cheng-Chang, Kuo-Min Su, Kai-Hsi Lu, Chi-Kang Lin, Peng-Hui Wang, Hsin-Yang Li, Mong-Lien Wang, Cheng-Kuo Lin, Mu-Hsien Yu, and Chia-Ming Chang. 2018. "Key Immunological Functions Involved in the Progression of Epithelial Ovarian Serous Carcinoma Discovered by the Gene Ontology-Based Immunofunctionome Analysis" International Journal of Molecular Sciences 19, no. 11: 3311. https://doi.org/10.3390/ijms19113311
APA StyleChang, C. -C., Su, K. -M., Lu, K. -H., Lin, C. -K., Wang, P. -H., Li, H. -Y., Wang, M. -L., Lin, C. -K., Yu, M. -H., & Chang, C. -M. (2018). Key Immunological Functions Involved in the Progression of Epithelial Ovarian Serous Carcinoma Discovered by the Gene Ontology-Based Immunofunctionome Analysis. International Journal of Molecular Sciences, 19(11), 3311. https://doi.org/10.3390/ijms19113311