T-2 Toxin Exposure Induces Apoptosis in TM3 Cells by Inhibiting Mammalian Target of Rapamycin/Serine/Threonine Protein Kinase(mTORC2/AKT) to Promote Ca2+Production


Abstract
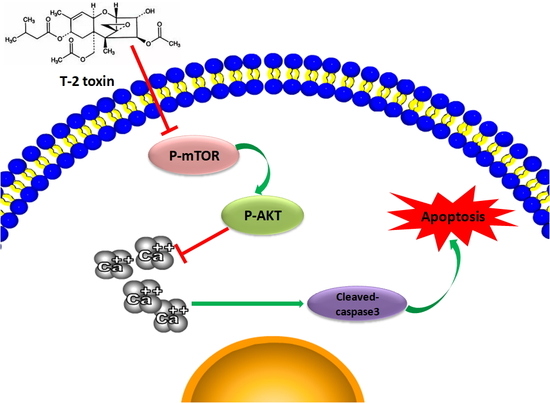
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. TM3 Cell Viability
2.2. T-2 Toxin Induces Intrinsic Apoptosis in TM3 Cells
2.3. Induction of Ca2+Is Involved in T-2 Toxin-Mediated TM3 Cells Apoptosis
2.4. The mTOR/Akt Pathway Inhibition Is Related to T-2 Toxin-Mediated Apoptosis
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Drug Treatment
4.4. Growth Inhibition Assay
4.5. Western Blotting
4.6. Detection of Apoptosis
4.7. Measurement of Intracellular Ca2+ Concentration
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zain, M.E. Impact of mycotoxins on humans and animals. J. Saudi Chem. Soc. 2011, 15, 129–144. [Google Scholar] [CrossRef]
- Adhikari, M.; Negi, B.; Kaushik, N.; Adhikari, A.; Al-Khedhairy, A.A.; Kaushik, N.K.; Choi, E.H. T-2 mycotoxin: Toxicological effects and decontamination strategies. Oncotarget 2017, 8, 33933–33952. [Google Scholar] [CrossRef] [PubMed]
- Marin, S.; Ramos, A.J.; Cano-Sancho, G.; Sanchis, V. Mycotoxins: Occurrence, toxicology, and exposure assessment. Food Chem. Toxicol. 2013, 60, 218–237. [Google Scholar] [CrossRef] [PubMed]
- Salimian, J.; Arefpour, M.A.; Riazipour, M.; Poursasan, N. Immunomodulatory effects of selenium and vitamin e on alterations in t lymphocyte subsets induced by T-2 toxin. Immunopharmacol. Immunotoxicol. 2014, 36, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Shinozuka, J.; Suzuki, M.; Noguchi, N.; Sugimoto, T.H.; Uetsuka, K.; Nakayama, H.; Doi, K. T-2 toxin-induced apoptosis in hematopoietic tissues of mice. Toxicol. Pathol. 1998, 26, 674–681. [Google Scholar] [CrossRef] [PubMed]
- Afsah-Hejri, L.; Jinap, S.; Hajeb, P.; Radu, S.; Shakibazadeh, S. A review on mycotoxins in food and feed: Malaysia case study. Compr. Rev. Food Sci. Food Saf. 2013, 12, 629–651. [Google Scholar] [CrossRef]
- Kiessling, K.H. Biochemical mechanism of action of mycotoxins. Pure Appl. Chem. 1986, 58, 327–338. [Google Scholar] [CrossRef]
- Sehata, S.; Kiyosawa, N.; Makino, T.; Atsumi, F.; Ito, K.; Yamoto, T.; Teranishi, M.; Baba, Y.; Uetsuka, K.; Nakayama, H.; et al. Morphological and microarray analysis of t-2 toxin-induced rat fetal brain lesion. Food Chem. Toxicol. 2004, 42, 1727–1736. [Google Scholar] [CrossRef] [PubMed]
- Doi, K.; Ishigami, N.; Sehata, S. T-2 toxin-induced toxicity in pregnant mice and rats. Int. J. Mol. Sci. 2008, 9, 2146–2158. [Google Scholar] [CrossRef] [PubMed]
- Canady, R.A.; Coker, R.D.; Egan, S.K.; Krska, R.; Olsen, M.; Resnik, S.; Schlatter, J. T-2 and HT-2 Toxins. International Programme on Chemical Safty. Available online: http://www.inchem.org/documents/jecfa/jecmono/v47je06.htm (accessed on 20 September 2001).
- Chen, J.H.; Cao, J.L.; Chu, Y.L.; Wang, Z.L.; Yang, Z.T.; Wang, H.L. T-2 toxin-induced apoptosis involving fas, p53, bcl-xl, bcl-2, bax and caspase-3 signaling pathways in human chondrocytes. J. Zhejiang Univ.-Sci. B (Biomed. Biotechnol.) 2008, 9, 455. [Google Scholar] [CrossRef] [PubMed]
- Nagase, M.; Alam, M.M.; Tsushima, A.; Yoshuzawa, T.; Sakato, N. Apoptosis induction by T-2 toxin: Activation of caspase-9, caspase-3 and DEF-40/CAD through cytosolic release of cytochrome c in HL-60 cells. Biosci. Biotechnol. Biochem. 2001, 65, 1741–1747. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Z.; Yang, D.; Huang, Y.; Wang, S. Study on the apoptosis mechanism induced by t-2 toxin. PLoS ONE 2013, 8, e83105. [Google Scholar] [CrossRef] [PubMed]
- Mu, Y.P.; Lin, D.C.; Yan, F.R.; Jiao, H.X.; Gui, L.X.; Lin, M.J. Alterations in caveolin-1 expression and receptor-operated ca2+ entry in the aortas of rats with pulmonary hypertension. Cell. Physiol. Biochem. 2016, 39, 438–452. [Google Scholar] [CrossRef] [PubMed]
- Pinton, P.; Giorgi, C.; Siviero, R.; Zecchini, E.; Rizzuto, R. Calcium and apoptosis: ER-mitochondria Ca2+ transfer in the control of apoptosis. Oncogene 2008, 27, 6407. [Google Scholar] [CrossRef] [PubMed]
- Holme, J.A.; Morrison, E.; Samuelsen, J.T.; Wiger, R.; Låg, M.; Schwarze, P.E.; Bernhoft, A.; Refsnes, M. Mechanisms involved in the induction of apoptosis by T-2 and HT-2 toxins in HL-60 human promyelocytic leukemia cells. Cell Biol. Toxicol. 2003, 19, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.M.; Gulhati, P.; Rampy, B.A.; Han, Y.; Rychahou, P.G.; Doan, H.Q.; Weiss, H.L.; Evers, B.M. Novel expression patterns of PI3K/AKT/mTOR signaling pathway components in colorectal cancer. J. Am. Coll. Surg. 2010, 210, 767–776. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Liu, Y.; Li, Y.; Zhe, X.; Zhang, S.; Zhang, L. Neuroglobin promotes the proliferation and suppresses the apoptosis of glioma cells by activating the PI3K/AKT pathway. Mol. Med. Rep. 2018, 17, 2757–2763. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.T.; Wang, W.W.; Ren, L.H.; Zhao, X.X.; Wang, Z.H.; Zhuang, D.L.; Bai, Y.N. The mTORC2/Akt/NFκB pathway-mediated activation of trpc6 participates in adriamycin-induced podocyte apoptosis. Cell. Physiol. Biochem. 2016, 40, 1079–1093. [Google Scholar] [CrossRef] [PubMed]
- Ding, F.; Zhang, X.; Li, X.; Zhang, Y.; Li, B.; Ding, J. Mammalian target of rapamycin complex 2 signaling pathway regulates transient receptor potential cation channel 6 in podocytes. PLoS ONE 2014, 9, e112972. [Google Scholar] [CrossRef] [PubMed]
- Tobaben, S.; Grohm, J.; Seiler, A.; Conrad, M.; Plesnila, N.; Culmsee, C. Bid-mediated mitochondrial damage is a key mechanism in glutamate-induced oxidative stress and aif-dependent cell death in immortalized ht-22 hippocampal neurons. Cell Death Differ. 2011, 18, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Grahammer, F.; Wanner, N.; Huber, T.B. mTOR controls kidney epithelia in health and disease. Nephrol. Dial. Transplant. 2014, 29 (Suppl. 1). [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.J.; Park, Y.J.; Park, J.Y.; Jeong, H.O.; Kim, D.H.; Ha, Y.M.; Kim, J.M.; Song, Y.M.; Heo, H.S.; Yu, B.P.; et al. Inhibitory effect of mTOR activator MHY1485 on autophagy: Suppression of lysosomal fusion. PLoS ONE 2012, 7, e43418. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Zhang, Y.F.; Liang, A.M.; Kong, X.F.; Li, Y.X.; Ma, K.W.; Jing, A.H.; Feng, S.Y.; Qiao, X.L.; et al. Toxic effects of t-2 toxin on reproductive system in male mice. Toxicol. Ind. Health 2010, 26, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Matias, F.B.; Yi, J.E.; Wu, J. T-2 toxin-induced cytotoxicity and damage on tm3 leydig cells. Comp. Biochem. Physiol. Part C 2016, 181–182, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, Y.; Jing, A.; Ma, K.; Gong, Q.; Qin, C. Effects of t-2 toxin on testosterone biosynthesis in mouse leydig cells. Toxicol. Ind. Health 2012, 30, 873–877. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Zhang, Y.F.; Li, Y.X.; Guan, G.P.; Kong, X.F.; Liang, A.M.; Ma, K.W.; Da, L.G.; Bai, X.F. Effects of t-2 toxin on the regulation of steroidogenesis in mouse leydig cells. Toxicol. Ind. Health 2016, 32, 1801–1807. [Google Scholar] [CrossRef] [PubMed]
- Jian, Y.Y.; Yong, F.Z.; Xiang, P.M.; Yuan, X.L.; Kai, W.M.; Xue, F.B. T-2 toxin inhibits gene expression and activity of key steroidogenesis enzymes in mouse leydig cells. Toxicol. In Vitro 2015, 29, 1166–1171. [Google Scholar] [CrossRef]
- Wu, J.; Tu, D.; Yuan, L.Y.; Yi, J.E.; Tian, Y. T-2 toxin regulates steroid hormone secretion of rat ovarian granulosa cells through camp-pka pathway. Toxicol. Lett. 2015, 232, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.B.; Sun, Y.Y.; Zheng, Y.L.; Yu, C.Q.; Lin, H.Q.; Pang, J.Y. A study on blocking store-operated Ca2+ entry in pulmonary arterial smooth muscle cells with xyloketals from marine fungi. Acta Pharm. 2017, 67. [Google Scholar] [CrossRef] [PubMed]
- Decuypere, J.P.; Paudel, R.C.; Parys, J.B.; Bultynck, G. Intracellular Ca2+ signaling: A novel player in the canonical mtor-controlled autophagy pathway. Commun. Integr. Biol. 2013, 6, e25429. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.H.; Shih, Y.L.; Ko, W.C.; Wei, Y.H.; Shih, C.M. Cadmium-induced autophagy and apoptosis are mediated by a calcium signaling pathway. Cell. Mol. Life Sci. 2008, 65, 3640–3652. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Tian, X.; Huang, X.; Fang, Y.; Qiao, D. Resveratrol-induced mitochondrial dysfunction and apoptosis are associated with Ca2+, and mcicr-mediated mpt activation in hepg2 cells. Mol. Cell. Biochem. 2007, 302, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Carafoli, E. The calcium cycle of mitochondria. Febs Lett. 1979, 104, 1. [Google Scholar] [CrossRef]
- Masini, A.; Trenti, T.; Ceccarelli-Stanzani, D.; Ventura, E. The effect of ferric iron complex on isolated rat liver mitochondria. II. Ion movements. Biochim. Biophys. Acta 1985, 810, 20–26. [Google Scholar] [CrossRef]
- Ma, X.M.; Blenis, J. Molecular mechanisms of mTOR-mediated translational control. Nat. Rev. Mol. Cell Boil. 2009, 10, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Laplante, M.; Sabatini, D.M. mTOR signaling in growth control and disease. Cell 2012, 149, 274–293. [Google Scholar] [CrossRef] [PubMed]
- Sarbassov, D.D.; Guertin, D.A.; Ali, S.M.; Sabatini, D.M. Phosphorylation and regulation of akt/pkb by the rictor-mtor complex. Science 2005, 307, 1098. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chiu, J.; Zhang, H.; Qi, T.; Tang, Q.; Ma, K.; Lu, H.; Li, G. Autophagic cell death induced by resveratrol depends on the Ca(2+)/ampk/mtor pathway in a549 cells. Biochem. Pharmacol. 2013, 86, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Gulati, P.; Gaspers, L.D.; Dann, S.G.; Joaquin, M.; Nobukuni, T.; Natt, F.; Kozma, S.C.; Thomas, A.P.; Thomas, G. Amino acids activate mtor complex 1 via ca/cam signaling to hvps34. Cell Metab. 2008, 7, 456. [Google Scholar] [CrossRef] [PubMed]
- Shin, C.S.; Huh, W.K. Bidirectional regulation between TORC1 and autophagy in Saccharomyces cerevisiae. Autophagy 2011, 7, 854–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Zhou, Y.; Yuan, Z.; Yi, J.; Chen, J.; Wang, N.; Tian, Y. Crosstalk between Autophagy and Apoptosis Modulates T-2 Toxin-Induced Toxicity in Liver Cells. Preprints 2018, 2018100034. [Google Scholar] [CrossRef]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Yang, C.; Yuan, Z.; Yi, J.; Wu, J. T-2 Toxin Exposure Induces Apoptosis in TM3 Cells by Inhibiting Mammalian Target of Rapamycin/Serine/Threonine Protein Kinase(mTORC2/AKT) to Promote Ca2+Production. Int. J. Mol. Sci. 2018, 19, 3360. https://doi.org/10.3390/ijms19113360
Wang J, Yang C, Yuan Z, Yi J, Wu J. T-2 Toxin Exposure Induces Apoptosis in TM3 Cells by Inhibiting Mammalian Target of Rapamycin/Serine/Threonine Protein Kinase(mTORC2/AKT) to Promote Ca2+Production. International Journal of Molecular Sciences. 2018; 19(11):3360. https://doi.org/10.3390/ijms19113360
Chicago/Turabian StyleWang, Ji, Chenglin Yang, Zhihang Yuan, Jine Yi, and Jing Wu. 2018. "T-2 Toxin Exposure Induces Apoptosis in TM3 Cells by Inhibiting Mammalian Target of Rapamycin/Serine/Threonine Protein Kinase(mTORC2/AKT) to Promote Ca2+Production" International Journal of Molecular Sciences 19, no. 11: 3360. https://doi.org/10.3390/ijms19113360
APA StyleWang, J., Yang, C., Yuan, Z., Yi, J., & Wu, J. (2018). T-2 Toxin Exposure Induces Apoptosis in TM3 Cells by Inhibiting Mammalian Target of Rapamycin/Serine/Threonine Protein Kinase(mTORC2/AKT) to Promote Ca2+Production. International Journal of Molecular Sciences, 19(11), 3360. https://doi.org/10.3390/ijms19113360