Cathepsin S Alters the Expression of Pro-Inflammatory Cytokines and MMP-9, Partially through Protease—Activated Receptor-2, in Human Corneal Epithelial Cells

 , , and
, , and 
Abstract
:1. Introduction
2. Results
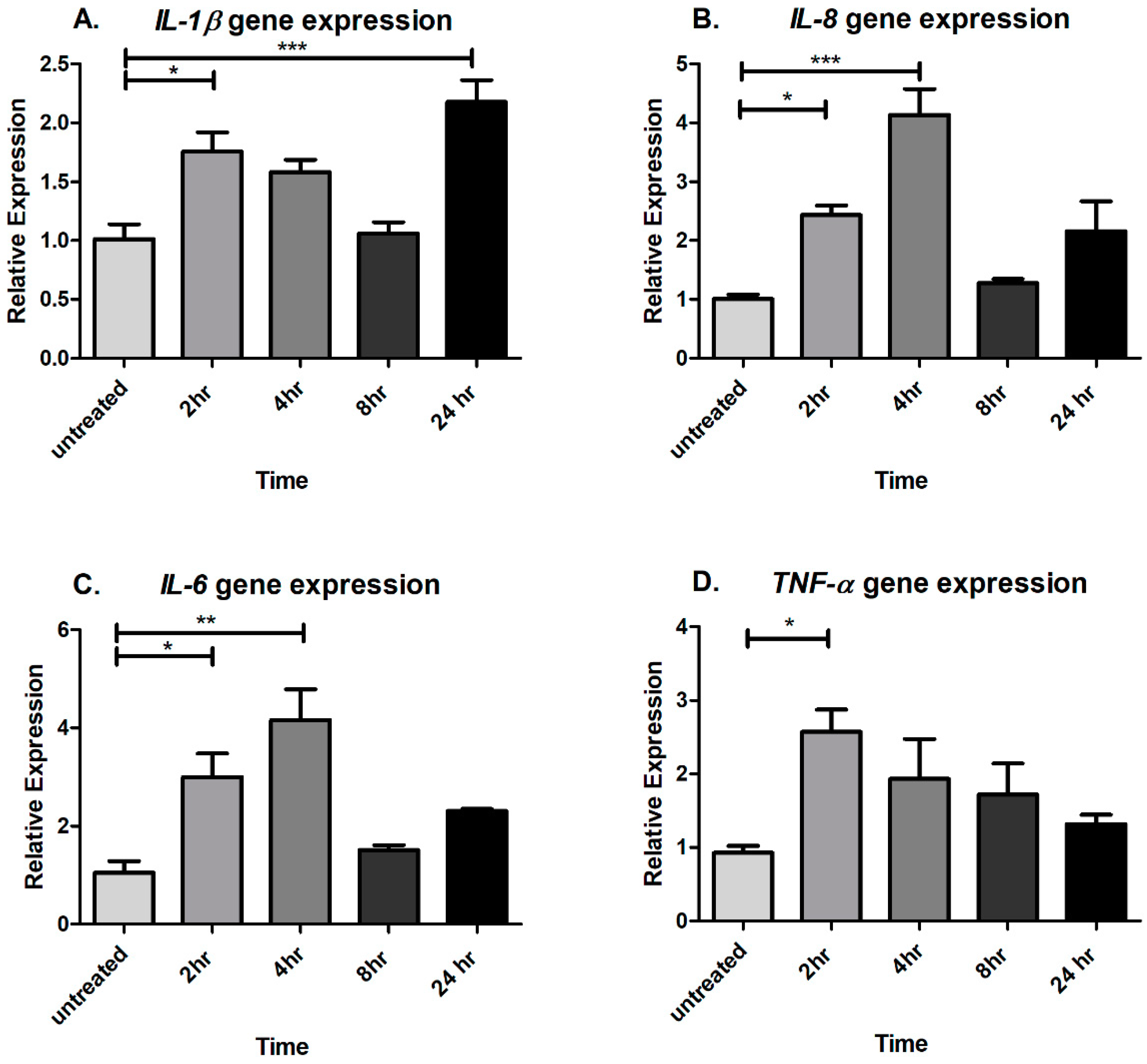
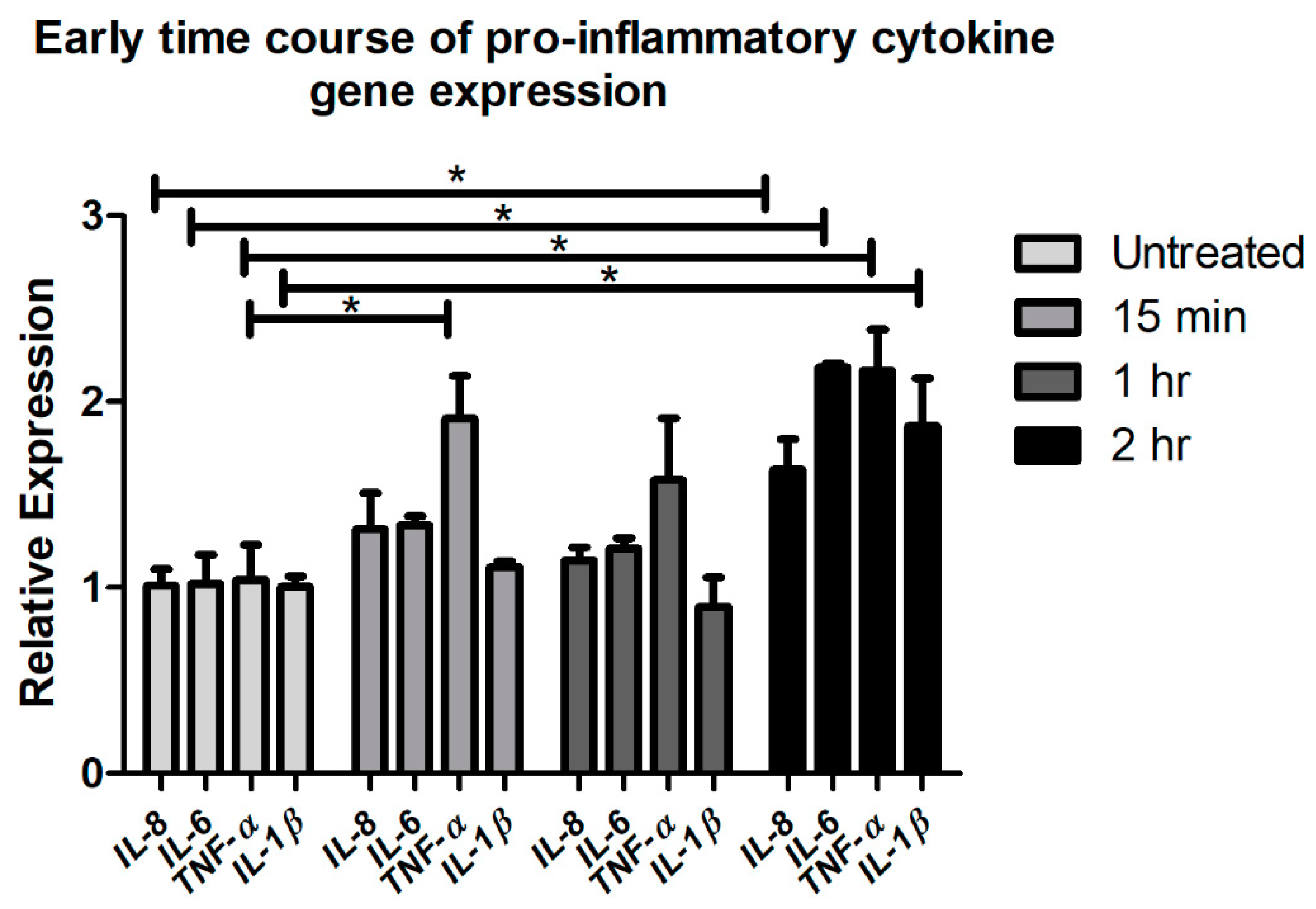
2.1. Acute CTSS Exposure Significantly Increases IL-1β, IL-8, IL-6, and TNF-α Gene and Protein Expression in Cell Culture Medium and Cell Lysates of Human Corneal Epithelial Cells
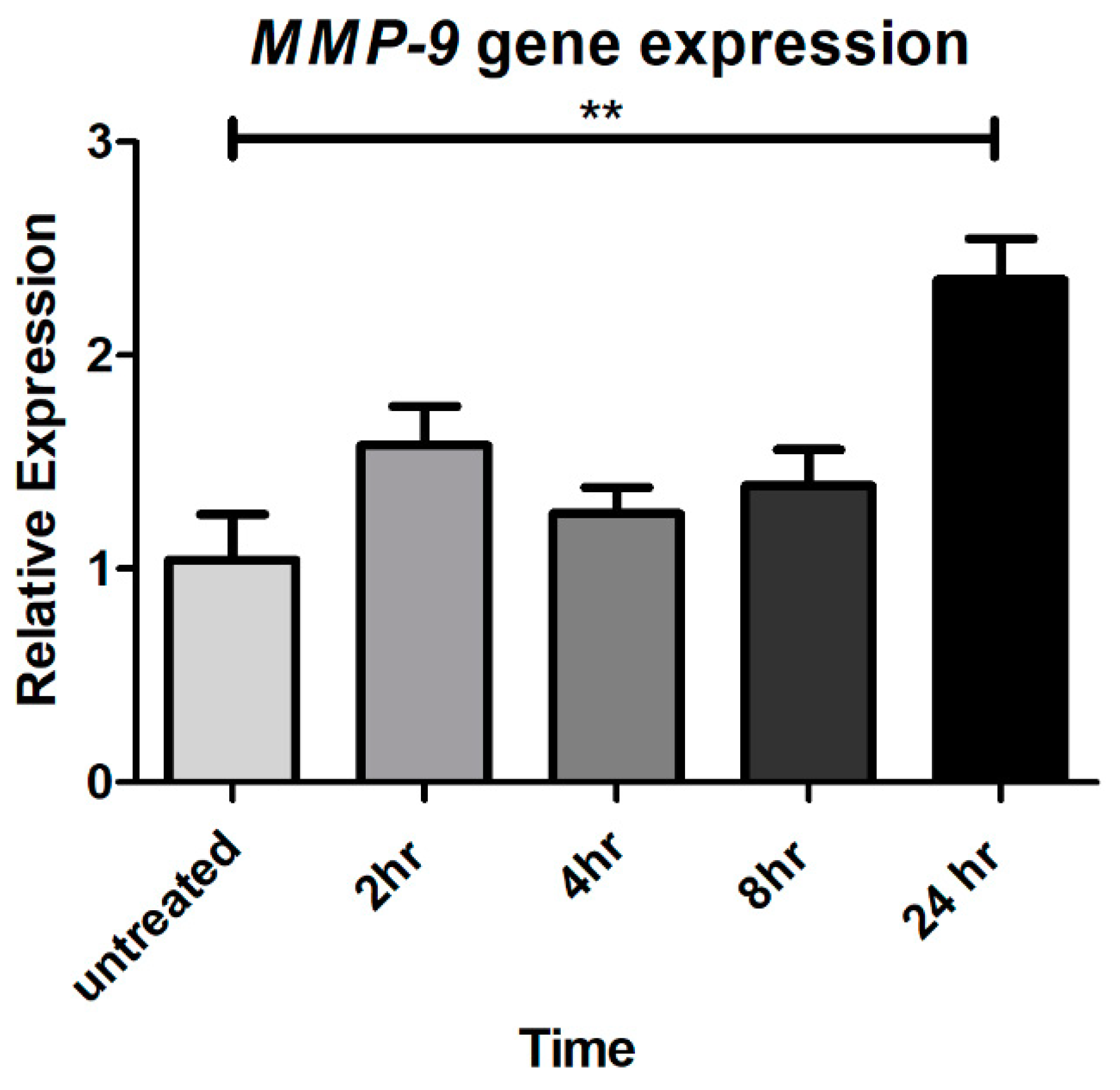
2.2. Chronic CTSS Exposure Significantly Increases MMP-9 Gene Expression
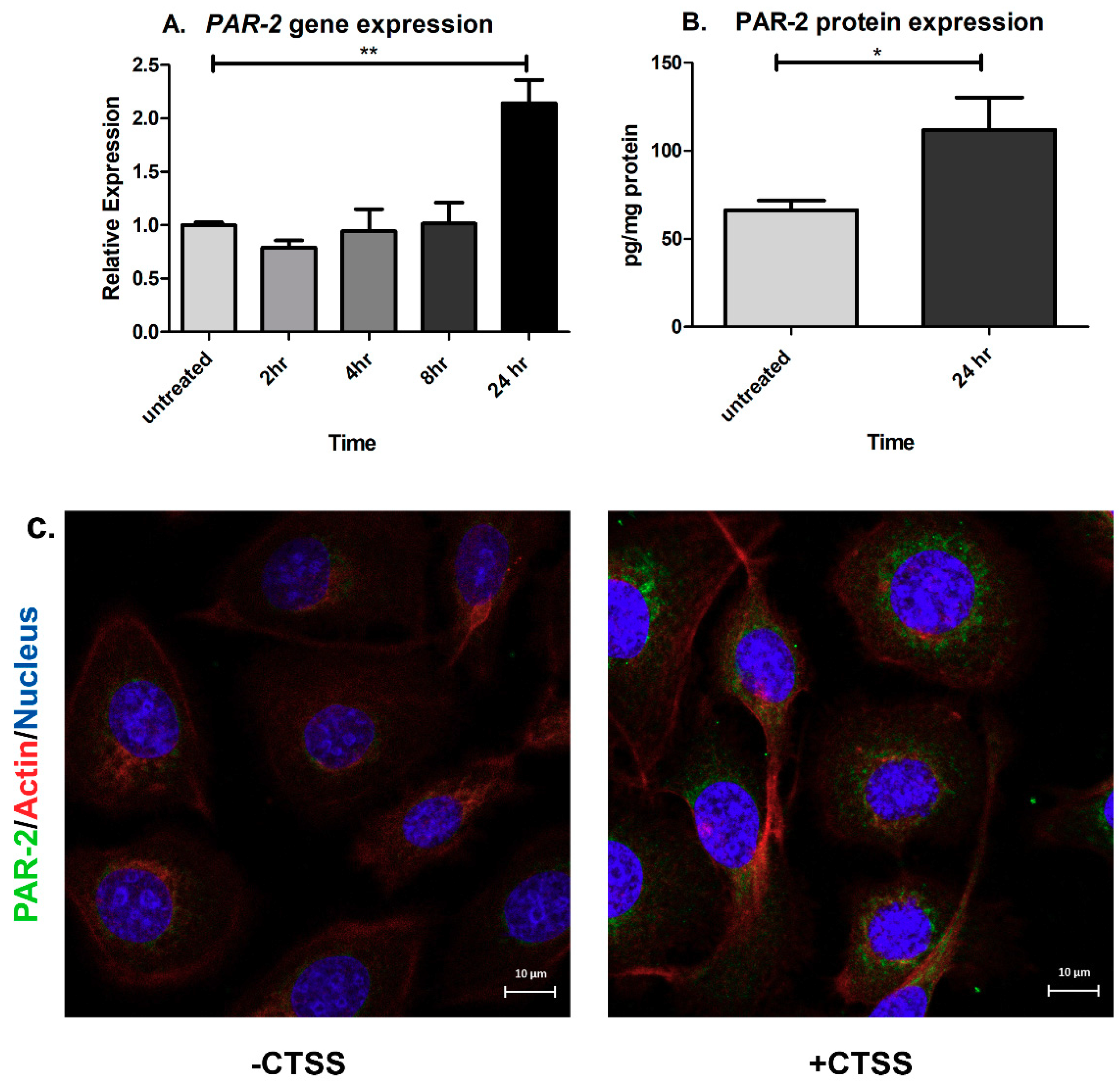
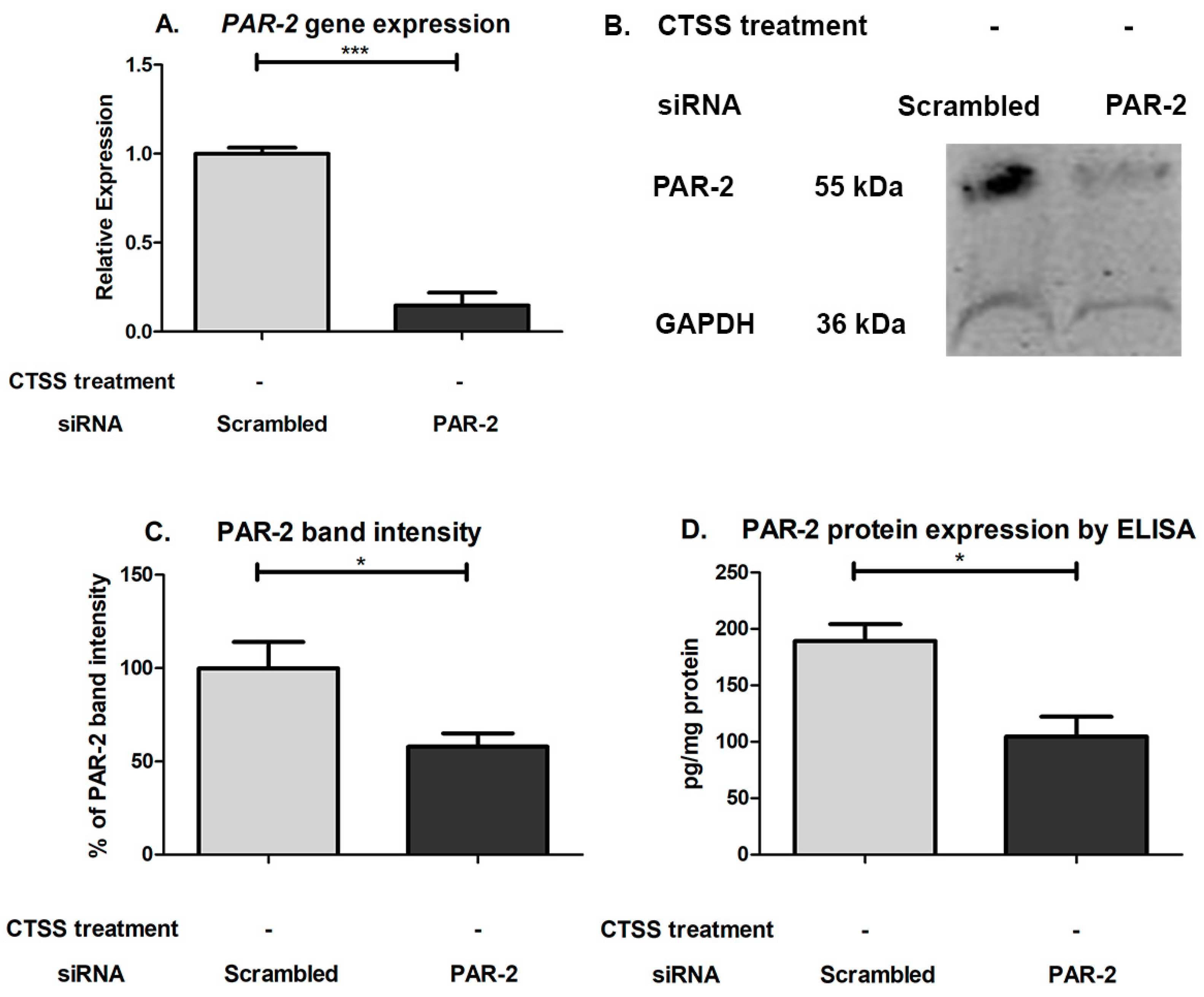
2.3. Chronic CTSS Exposure Significantly Increases PAR-2 Gene and Protein Expression
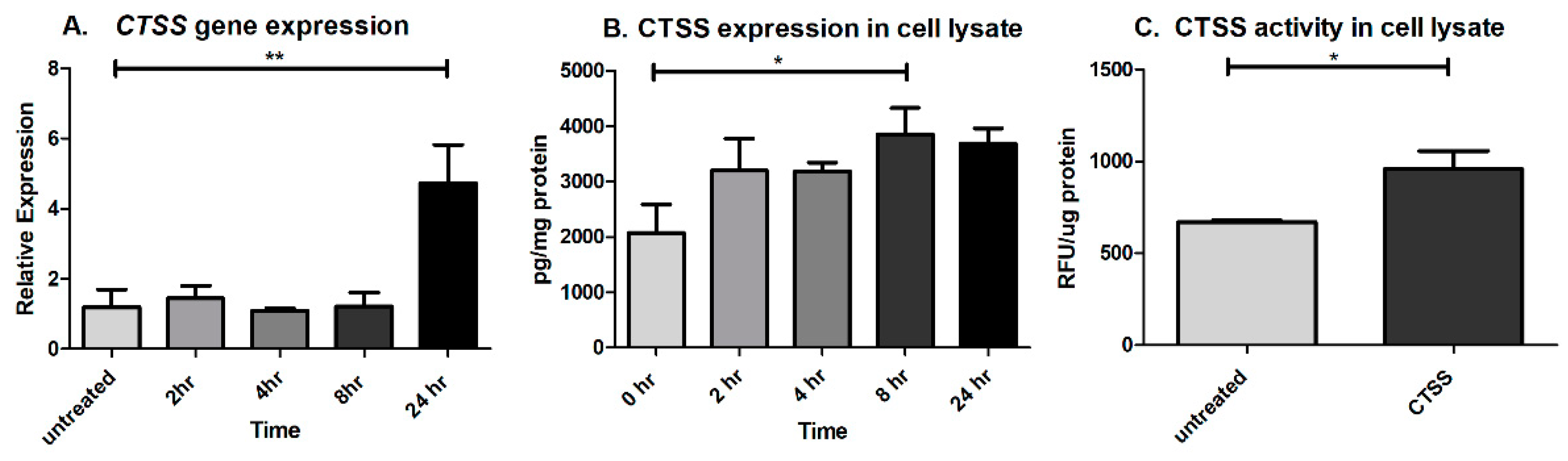
2.4. CTSS Exposure Significantly Increases CTSS Gene and Protein Expression at 8- and 24-h
2.5. Acute Exposure to Heat-Inactivated CTSS Does not Induce Pro-Inflammatory Cytokines
2.6. CTSS-Dependent Activation of PAR-2 Activation Is Involved in Induction of Pro-Inflammatory Cytokines and Proteases in Human Corneal Epithelial Cells
2.7. An Initial PAR-2 Dependent Increase in TNF-α after Acute CTSS Exposure May Drive IL-6 and IL-1β Gene Expression in Human Corneal Epithelial Cells
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Calculation of Human Recombinant CTSS Dosage for Treatment of Human Corneal Epithelial Cells
4.3. Cell Culture
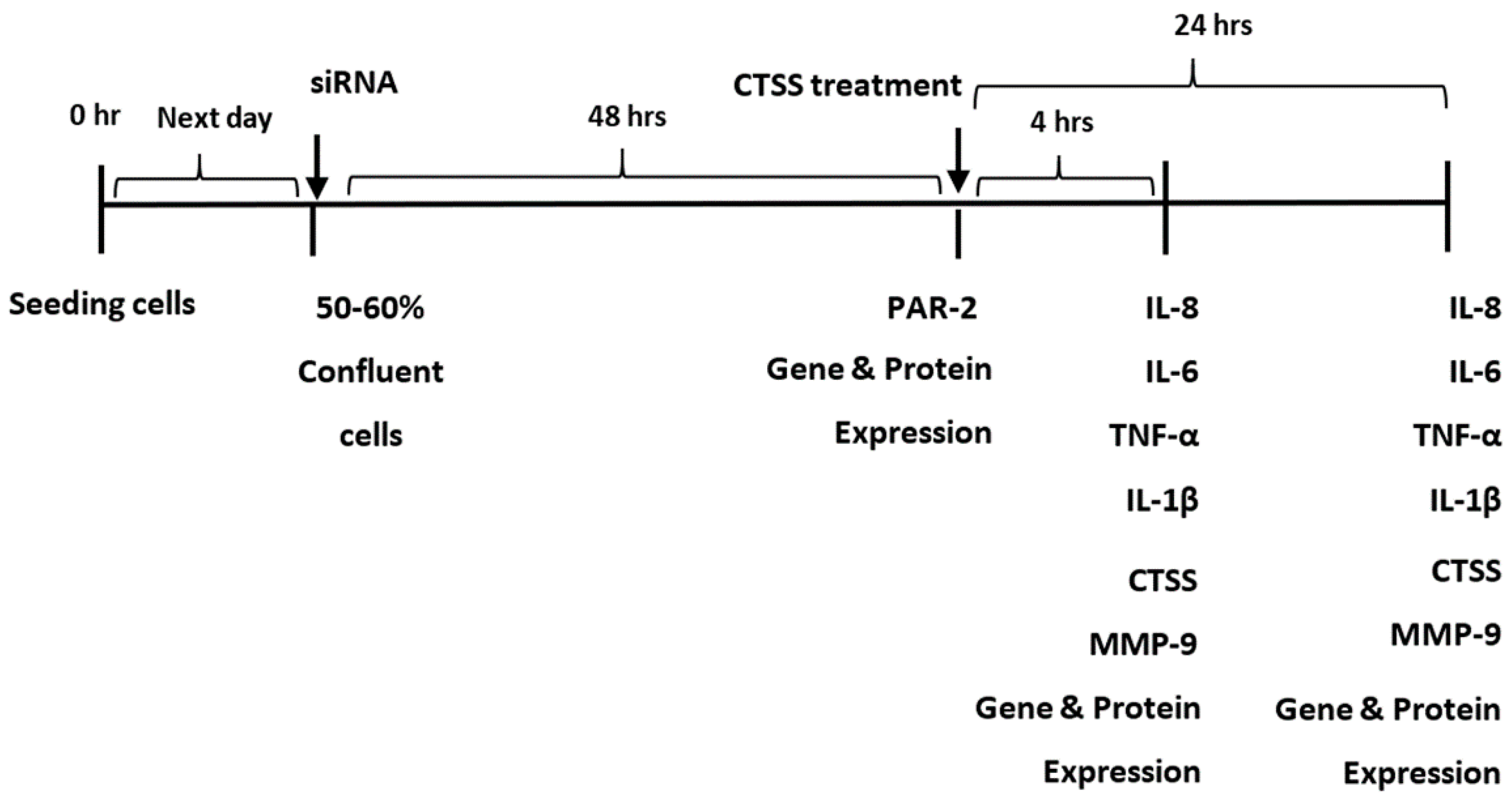
4.4. siRNA Transfection
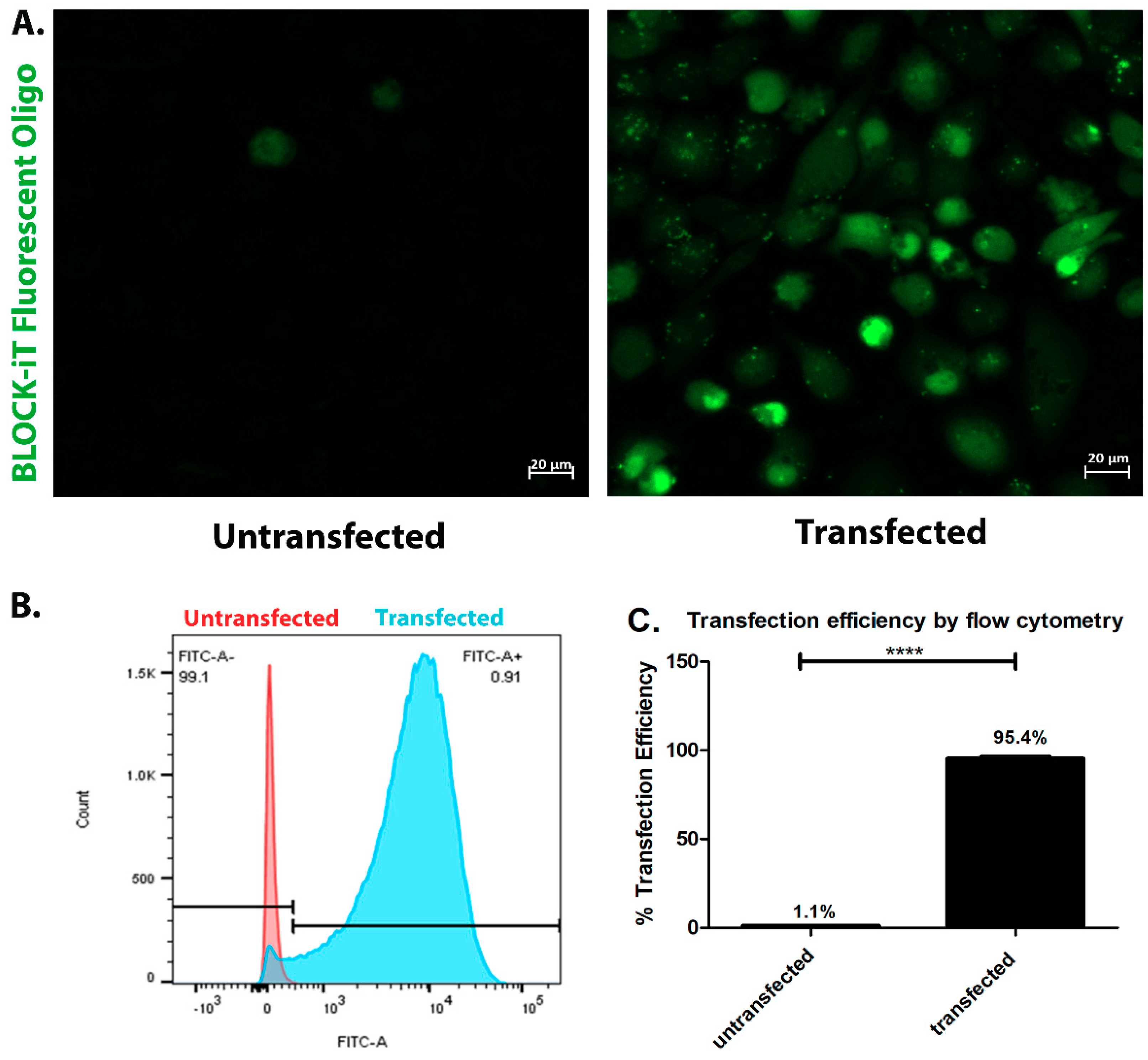
4.5. Transfection Efficiency
4.6. Treatment of Cells with Recombinant Human CTSS
4.7. Gene Expression
4.8. Multiplex Assay and ELISA Methods for Measurement of Pro-Inflammatory Cytokines in HCE-T Cell Medium and Lysates
4.9. ELISA Method for Measurement of CTSS in HCE-T Cell Lysates
4.10. ELISA Method for Measurement PAR-2 in HCE-T Cell Lysates
4.11. Multiplex Assay and ELISA Methods for Measurement of Pro-Inflammatory Cytokines, CTSS, and MMP-9 in siRNA Transfected HCE-T Cells
4.12. Immunofluorescence of PAR-2
4.13. CTSS Activity in HCE-T Cell Lysates
4.14. Western Blotting for PAR-2
4.15. Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| SS | Sjögren’s syndrome |
| LG | Lacrimal gland |
| CTSS | Cathepsin S |
| IL | Interleukin |
| TNF | Tumor necrosis factor |
| TGF | Transforming growth factor |
| IFN | Interferon |
| MMP | Matrix metalloproteinase |
| NOD | Non-obese diabetic |
| PAR | Protease-activated receptor |
| GPCR | G-protein-coupled receptor |
| HCE-T | Human corneal epithelial cell line |
| SEM | Standard error of the mean |
| Th | T helper cell |
| siRNA | Small interfering RNA |
| CD | Cluster of differentiation |
References
- Brito-Zeron, P.; Baldini, C.; Bootsma, H.; Bowman, S.J.; Jonsson, R.; Mariette, X.; Sivils, K.; Theander, E.; Tzioufas, A.; Ramos-Casals, M. Sjogren syndrome. Nat. Rev. Dis. Prim. 2016, 2, 16047. [Google Scholar] [CrossRef] [PubMed]
- Hyon, J.Y.; Lee, Y.J.; Yun, P.Y. Management of ocular surface inflammation in Sjogren syndrome. Cornea 2007, 26, S13–S15. [Google Scholar] [CrossRef] [PubMed]
- Stern, M.E.; Pflugfelder, S.C. Inflammation in dry eye. Ocul. Surf. 2004, 2, 124–130. [Google Scholar] [CrossRef]
- Byun, Y.S.; Lee, H.J.; Shin, S.; Chung, S.H. Elevation of autophagy markers in Sjogren syndrome dry eye. Sci. Rep. 2017, 7, 17280. [Google Scholar] [CrossRef] [PubMed]
- Foulks, G.N.; Forstot, S.L.; Donshik, P.C.; Forstot, J.Z.; Goldstein, M.H.; Lemp, M.A.; Nelson, J.D.; Nichols, K.K.; Pflugfelder, S.C.; Tanzer, J.M.; et al. Clinical guidelines for management of dry eye associated with Sjogren disease. Ocul. Surf. 2015, 13, 118–132. [Google Scholar] [CrossRef] [PubMed]
- Foulks, G.N.; Bunya, V.Y.; Hammitt, K.M.; Nichols, K.K.; Pflugfelder, S.C.; Vivino, F.B. Improving diagnosis and outcomes of Sjogren’s disease through targeting dry eye patients. A Continuing medical education enduring material. Ocul. Surf. 2015, 13 (Suppl. 1), S1–S33. [Google Scholar] [CrossRef]
- Shi, G.; Villadangos, J.A.; Dranoff, G.; Small, C.; Gu, L.; Haley, K.J.; Riese, R.; Ploegh, H.L.; Chapman, H.A. Cathepsin S required for normal MHC class II peptide loading and germinal center development. Immunity 1999, 10, 197–206. [Google Scholar] [CrossRef]
- Riese, R.J.; Wolf, P.R.; Bromme, D.; Natkin, L.R.; Villadangos, J.A.; Ploegh, H.L.; Chapman, H.A. Essential role for cathepsin S in MHC class II-associated invariant chain processing and pepetide loading. Immunity 1996, 4, 357–366. [Google Scholar] [CrossRef]
- Edman, M.C.; Janga, S.R.; Meng, Z.; Bechtold, M.; Chen, A.F.; Kim, C.; Naman, L.; Sarma, A.; Teekappanavar, N.; Kim, A.Y.; et al. Increased Cathepsin S activity associated with decreased protease inhibitory capacity contributes to altered tear proteins in Sjogren’s syndrome patients. Sci. Rep. 2018, 8, 11044. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.M.; Troutt, J.S.; Knierman, M.D.; Siegel, R.W.; Qian, Y.W.; Ackermann, B.L.; Konrad, R.J. Determination of Cathepsin S abundance and activity in human plasma and implications for clinical investigation. Anal. Biochem. 2012, 430, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Helske, S.; Syvaranta, S.; Lindstedt, K.A.; Lappalainen, J.; Oorni, K.; Mayranpaa, M.I.; Lommi, J.; Turto, H.; Werkkala, K.; Kupari, M.; et al. Increased expression of elastolytic cathepsins S, K, and V and their inhibitor cystatin C in stenotic aortic valves. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 1791–1798. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.S.; Li, W.; Keyszer, G.; Weber, E.; Levy, R.; Klein, M.J.; Gravallese, E.M.; Goldring, S.R.; Bromme, D. Comparison of cathepsins K and S expression within the rheumatoid and osteoarthritic synovium. Arthritis Rheum. 2002, 46, 663–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haves-Zburof, D.; Paperna, T.; Gour-Lavie, A.; Mandel, I.; Glass-Marmor, L.; Miller, A. Cathepsins and their endogenous inhibitors cystatins: Expression and modulation in multiple sclerosis. J. Cell. Mol. Med. 2011, 15, 2421–2429. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, T.; Nakamura, H.; Owen, C.A.; Yoshida, S.; Tsuduki, K.; Chubachi, S.; Shirahata, T.; Mashimo, S.; Nakamura, M.; Takahashi, S.; et al. Plasma Cathepsin S and Cathepsin S/Cystatin C Ratios Are Potential Biomarkers for COPD. Dis. Mark. 2016, 2016, 4093870. [Google Scholar] [CrossRef] [PubMed]
- Pflugfelder, S.C.; Jones, D.; Ji, Z.; Afonso, A.; Monroy, D. Altered cytokine balance in the tear fluid and conjunctiva of patients with Sjögren’s syndrome keratoconjunctivitis sicca. Curr. Eye Res. 2009, 19, 201–211. [Google Scholar] [CrossRef]
- Yoon, K.; Jeong, I.; Park, Y.; Yang, S. Interleukin-6 and tumor necrosis factor-α levels in tears of patients with dry eye syndrome. Cornea 2007, 26, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Solomon, A.; Dursun, D.; Liu, Z.; Xie, Y.; Macri, A.; Pflugfelder, S.C. Pro- and anti-inflammatory forms of Interleukin-1 in the tear fluid and conjunctiva of Patients with dry-eye disease. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2283–2292. [Google Scholar]
- Liu, R.; Gao, C.; Chen, H.; Li, Y.; Jin, Y.; Qi, H. Analysis of Th17-associated cytokines and clinical correlations in patients with dry eye disease. PLoS ONE 2017, 12, e0173301. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, S.; Tong, L. Practical issues concerning tear protein assays in dry eye. Eye Vis. 2014, 1, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ram, M.; Sherer, Y.; Shoenfeld, Y. Matrix metalloproteinase-9 and autoimmune diseases. J. Clin. Immunol. 2006, 26, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Aluri, H.S.; Kublin, C.L.; Thotakura, S.; Armaos, H.; Samizadeh, M.; Hawley, D.; Thomas, W.M.; Leavis, P.; Makarenkova, H.P.; Zoukhri, D. Role of Matrix Metalloproteinases 2 and 9 in Lacrimal Gland Disease in Animal Models of Sjogren’s syndrome. Investig. Ophthalmol. Vis. Sci. 2015, 56, 5218–5228. [Google Scholar] [CrossRef] [PubMed]
- Kossakowska, A.; Edwards, D.R.; Prusinkiewicz, C.; Zhang, M.C.; Guo, D.; Urbanski, S.J.; Grogan, T.; Marquez, L.A.; Janowska-Wieczorek, J. Interleukin-6 regulation of matrix metalloproteinase (MMP-2 and MMP-9) and tissue inhibitor of metalloproteinase (TIMP-1) expression in malignant Non-Hodgkin’s lymphomas. Blood 1999, 94, 2080–2089. [Google Scholar] [PubMed]
- Chakrabarti, S.; Patel, K.D. Regulation of matrix metalloproteinase-9 release from IL-8-stimulated human neutrophils. J. Leukoc. Biol. 2005, 78, 279–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, G.M.; Ledee, D.R.; Feuer, W.J.; Fini, M.E. Cytokines and signaling pathways regulating matrix metalloproteinase-9 (MMP-9) expression in corneal epithelial cells. J. Cell. Physiol. 2009, 221, 402–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.Q.; Lokeshwar, B.L.; Solomon, A.; Monroy, D.; Ji, Z.; Pflugfelder, S.C. Regulation of MMP-9 production by human corneal epithelial cells. Exp. Eye Res. 2001, 73, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Chotikavanich, S.; de Paiva, C.S.; Li, D.Q.; Chen, J.J.; Bian, F.; Farley, W.J.; Pflugfelder, S.C. Production and activity of matrix metalloproteinase-9 on the ocular surface increase in dysfunctional tear syndrome. Investig. Ophthalmol. Vis. Sci. 2009, 50, 3203–3209. [Google Scholar] [CrossRef] [PubMed]
- Yau, M.K.; Liu, L.; Fairlie, D.P. Toward drugs for protease-activated receptor 2 (PAR2). J. Med. Chem. 2013, 56, 7477–7497. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, R.; Noorbakhsh, F.; Defea, K.; Hollenberg, M.D. Targeting proteinase-activated receptors: Therapeutic potential and challenges. Nat. Rev. Drug Discov. 2012, 11, 69–86. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Kim, J.M.; Jeong, S.K.; Jeon, J.E.; Yoon, H.J.; Jeong, M.K.; Lee, S.H. Protease-activated receptor-2 mediates the expression of inflammatory cytokines, antimicrobial peptides, and matrix metalloproteinases in keratinocytes in response to Propionibacterium acnes. Arch. Dermatol. Res. 2010, 302, 745–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikolic-Paterson, D.J. Cathepsin S-Dependent Protease-Activated Receptor-2 Activation: A New Mechanism of Endothelial Dysfunction. J. Am. Soc. Nephrol. 2016, 27, 1577–1579. [Google Scholar] [CrossRef] [PubMed]
- Gruber, B.L.; Marchese, M.J.; Santiago-Schwarz, F.; Martin, C.A.; Zhang, J.; Kew, R.R. Protease-activated receptor-2 (PAR-2) expression in human fibroblasts is regulated by growth factors and extracellular matrix. J. Investig. Dermatol. 2004, 123, 832–839. [Google Scholar] [CrossRef] [PubMed]
- Vergnolle, N.; Bunnett, N.W.; Sharkey, K.A.; Brussee, V.; Compton, S.J.; Grady, E.F.; Cirino, G.; Gerard, N.; Basbaum, A.I.; Andrade-Gordon, P.; et al. Proteinase-activated receptor-2 and hyperalgesia: A novel pain pathway. Nat. Med. 2001, 7, 821–826. [Google Scholar] [CrossRef] [PubMed]
- Ramelli, G.; Fuertes, S.; Narayan, S.; Busso, N.; Acha-Orbea, H.; So, A. Protease-activated receptor 2 signalling promotes dendritic cell antigen transport and T-cell activation in vivo. Immunology 2010, 129, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Lang, R.; Song, P.I.; Legat, F.J.; Lavker, R.M.; Harten, B.; Kalden, H.; Grady, E.F.; Bunnett, N.W.; Armstrong, C.A.; Ansel, J.C. Human Corneal Epithelial Cells Express Functional PAR-1 and PAR-2. Investig. Ophthalmol. Vis. Sci. 2003, 44. [Google Scholar] [CrossRef]
- Iablokov, V.; Hirota, C.L.; Peplowski, M.A.; Ramachandran, R.; Mihara, K.; Hollenberg, M.D.; MacNaughton, W.K. Proteinase-activated receptor 2 (PAR2) decreases apoptosis in colonic epithelial cells. J. Biol. Chem. 2014, 289, 34366–34377. [Google Scholar] [CrossRef] [PubMed]
- Dulon, S.; Cande, C.; Bunnett, N.W.; Hollenberg, M.D.; Chignard, M.; Pidard, D. Proteinase-activated receptor-2 and human lung epithelial cells: Disarming by neutrophil serine proteinases. Am. J. Respir. Cell Mol. Biol. 2003, 28, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Lieu, T.; Barlow, N.; Metcalf, M.; Veldhuis, N.A.; Jensen, D.D.; Kocan, M.; Sostegni, S.; Haerteis, S.; Baraznenok, V.; et al. Cathepsin S causes inflammatory pain via biased agonism of PAR2 and TRPV4. J. Biol. Chem. 2014, 289, 27215–27234. [Google Scholar] [CrossRef] [PubMed]
- Kumar Vr, S.; Darisipudi, M.N.; Steiger, S.; Devarapu, S.K.; Tato, M.; Kukarni, O.P.; Mulay, S.R.; Thomasova, D.; Popper, B.; Demleitner, J.; et al. Cathepsin S Cleavage of Protease-Activated Receptor-2 on Endothelial Cells Promotes Microvascular Diabetes Complications. J. Am. Soc. Nephrol. 2016, 27, 1635–1649. [Google Scholar] [CrossRef] [PubMed]
- Tato, M.; Kumar, S.V.; Liu, Y.; Mulay, S.R.; Moll, S.; Popper, B.; Eberhard, J.N.; Thomasova, D.; Rufer, A.C.; Gruner, S.; et al. Cathepsin S inhibition combines control of systemic and peripheral pathomechanisms of autoimmune tissue injury. Sci. Rep. 2017, 7, 2775. [Google Scholar] [CrossRef] [PubMed]
- Cattaruzza, F.; Lyo, V.; Jones, E.; Pham, D.; Hawkins, J.; Kirkwood, K.; Valdez-Morales, E.; Ibeakanma, C.; Vanner, S.J.; Bogyo, M.; et al. Cathepsin S is activated during colitis and causes visceral hyperalgesia by a PAR2-dependent mechanism in mice. Gastroenterology 2011, 141, 1864–1874. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Bae, K.B.; Kim, M.O.; Yu, D.H.; Kim, H.J.; Yuh, H.S.; Ji, Y.R.; Park, S.J.; Kim, S.; Son, K.H.; et al. Overexpression of cathepsin S induces chronic atopic dermatitis in mice. J. Investig. Dermatol. 2012, 132, 1169–1176. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.K.; Cheung, V.C.; Lu, P.; Lau, E.Y.; Ma, S.; Tang, K.H.; Tong, M.; Lo, J.; Ng, I.O. Blockade of CD47-mediated cathepsin S/protease-activated receptor 2 signaling provides a therapeutic target for hepatocellular carcinoma. Hepatology 2014, 60, 179–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lourbakos, A.; Potempa, J.; Travis, J.; D’Andrea, M.R.; Andrade-Gordon, P.; Santulli, R.; Mackie, E.J.; Pike, R.N. Arginine-specific protease from Porphyromonas gingivalis activates protease-activated receptors on human oral epithelial cells and induces interleukin-6 secretion. Infect. Immun. 2001, 69, 5121–5130. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, C.; Tsuda, T.; Konishi, H.; Nakagawa, N.; Yamanishi, K. Tetracyclines modulate protease-activated receptor 2-mediated proinflammatory reactions in epidermal keratinocytes. Antimicrob. Agents Chemother. 2009, 53, 1760–1765. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, T.; Alizadeh, H. Role of protease-activated receptors 2 (PAR2) in ocular infections and inflammation. Recept. Clin. Investig. 2014, 1. [Google Scholar] [CrossRef]
- Lee, S.E.; Kim, J.M.; Jeong, S.K.; Choi, E.H.; Zouboulis, C.C.; Lee, S.H. Expression of Protease-Activated Receptor-2 in SZ95 Sebocytes and its Role in Sebaceous Lipogenesis, Inflammation, and Innate Immunity. J. Investig. Dermatol. 2015, 135, 2219–2227. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Yang, J.; Zhang, H.; Chen, X.; Zhang, C. Effect of PAR2 in regulating TNF-alpha and NAD(P)H oxidase in coronary arterioles in type 2 diabetic mice. Basic Res. Cardiol. 2011, 106, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Napoli, C.; Cicala, C.; Wallace, J.L.; De Nigris, F.; Santagada, V.; Caliendo, G.; Franoni, F.; Ignarro, L.J.; Cirino, G. Protease-activated receptor-2 modulates myocardial ischemia-reperfusion injury in the rat heart. Proc. Natl. Acad. Sci. USA 2000, 97, 3678–3683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vliagoftis, H.; Schwingshackl, A.; Milne, C.D.; Duszyk, M.; Hollenberg, M.D.; Wallace, J.L.; Befus, A.D.; Moqbel, R. Proteinase-activated receptor-2-mediated matrix metalloproteinase-9 release from airway epithelial cells. J. Allergy Clin. Immunol. 2000, 106, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Cenac, N.; Garcia-Villar, R.; Ferrier, L.; Larauche, M.; Vergnolle, N.; Bunnett, N.W.; Coelho, A.M.; Fioramonti, J.; Bueno, L. Proteinase-Activated Receptor-2-Induced Colonic Inflammation in Mice: Possible Involvement of Afferent Neurons, Nitric Oxide, and Paracellular Permeability. J. Immunol. 2003, 170, 4296–4300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coelho, A.M.; Vergnolle, N.; Guiard, B.; Fioramonti, J.; Bueno, L. Proteinases and proteinase-activated receptor 2: A possible role to promote visceral hyperalgesia in rats. Gastroenterology 2002, 122, 1035–1047. [Google Scholar] [CrossRef] [PubMed]
- Steinhoff, M.; Vergnolle, N.; Young, S.H.; Tognetto, M.; Amadesi, S.; Ennes, H.S.; Trevisani, M.; Hollenbreg, M.D.; Wallace, J.L.; Caughey, G.H.; et al. Agonist of proteinase-activated receptor 2 induce inflammation by a neurogenic mechanism. Nat. Med. 2000, 6, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Dekita, M.; Wu, Z.; Ni, J.; Zhang, X.; Liu, Y.; Yan, X.; Nakanishi, H.; Takahashi, I. Cathepsin S Is Involved in Th17 Differentiation Through the Upregulation of IL-6 by Activating PAR-2 after Systemic Exposure to Lipopolysaccharide from Porphyromonas gingivalis. Front. Pharmacol. 2017, 8, 470. [Google Scholar] [CrossRef] [PubMed]
- Ainscough, J.S.; Macleod, T.; McGonagle, D.; Brakefield, R.; Baron, J.M.; Alase, A.; Wittmann, M.; Stacey, M. Cathepsin S is the major activator of the psoriasis-associated proinflammatory cytokine IL-36gamma. Proc. Natl. Acad. Sci. USA 2017, 114, E2748–E2757. [Google Scholar] [CrossRef] [PubMed]
- Turner, N.A.; Mughal, R.S.; Warburton, P.; O’Regan, D.J.; Ball, S.G.; Porter, K.E. Mechanism of TNFalpha-induced IL-1alpha, IL-1beta and IL-6 expression in human cardiac fibroblasts: Effects of statins and thiazolidinediones. Cardiovasc. Res. 2007, 76, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.R.; Soares, R. Inflammation in Sjogren’s syndrome: Cause or consequence? Autoimmunity 2017, 50, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, M.V.; Delaleu, N.; Brokstad, K.A.; Berggreen, E.; Skarstein, K. Impaired salivary gland function in NOD mice: Association with changes in cytokine profile but not with histopathologic changes in the salivary gland. Arthritis Rheum. 2006, 54, 2300–2305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Wu, K.; Edman, M.; Schenke-Layland, K.; MacVeigh-Aloni, M.; Janga, S.R.; Schulz, B.; Hamm-Alvarez, S.F. Increased expression of cathepsins and obesity-induced proinflammatory cytokines in lacrimal glands of male NOD mouse. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5019–5029. [Google Scholar] [CrossRef] [PubMed]
- Hamm-Alvarez, S.F.; Janga, S.R.; Edman, M.C.; Madrigal, S.; Shah, M.; Frousiakis, S.E.; Renduchintala, K.; Zhu, J.; Bricel, S.; Silka, K.; et al. Tear cathepsin S as a candidate biomarker for Sjogren’s syndrome. Arthritis Rheumatol. 2014, 66, 1872–1881. [Google Scholar] [CrossRef] [PubMed]
- Schonbeck, U.; Mach, F.; Libby, P. Generation of biologically active IL-1 beta by matrix metalloproteinases: A novel caspase-1-independent pathway of IL-1 beta processing. J. Immunol. 1998, 161, 3340–3346. [Google Scholar] [PubMed]
- Charousos, G.P.; Flier, J.S.; Underhill, L.H. The hypothalamic-pituitary-adrenal axis and immune-mediated inflammation. N. Engl. J. Med. 1995, 332, 1351–1362. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.; Arend, W.; Sims, J.; Smith, D.; Blumberg, H.; O’Neill, L.; Goldbach-Mansky, R.; Pizarro, T.; Hoffman, H.; Bufler, P.; et al. IL-1 family nomenclature. Nat. Immunol. 2010, 11, 973. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Wang, X.; Hong, X.; Lu, L.; Liu, D. IL-36 cytokines in autoimmunity and inflammatory disease. Oncotarget 2018, 9, 2895–2901. [Google Scholar] [CrossRef] [PubMed]
- Rupanagudi, K.V.; Kulkarni, O.P.; Lichtnekert, J.; Darisipudi, M.N.; Mulay, S.R.; Schott, B.; Gruner, S.; Haap, W.; Hartmann, G.; Anders, H.J. Cathepsin S inhibition suppresses systemic lupus erythematosus and lupus nephritis because cathepsin S is essential for MHC class II-mediated CD4 T cell and B cell priming. Ann. Rheum. Dis. 2015, 74, 452–463. [Google Scholar] [CrossRef] [PubMed]
- Lieu, T.; Savage, E.; Zhao, P.; Edgington-Mitchell, L.; Barlow, N.; Bron, R.; Poole, D.P.; McLean, P.; Lohman, R.J.; Fairlie, D.P.; et al. Antagonism of the proinflammatory and pronociceptive actions of canonical and biased agonists of protease-activated receptor-2. Br. J. Pharmacol. 2016, 173, 2752–2765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araki-Sasaki, K.; Ohashi, Y.; Sasabe, T.; Hayashi, K.; Watanabe, H.; Tano, Y.; Handa, H. An SV40-immortalized human corneal epithelial cell line and its characterization. Investig. Ophthalmol. Vis. Sci. 1995, 36, 614–621. [Google Scholar]
- Liu, H.; Qin, Y.; Kong, Z.; Shao, Q.; Su, Z.; Wang, S.; Chen, J. siRNA Targeting the 2Apro Genomic Region Prevents Enterovirus 71 Replication In Vitro. PLoS ONE 2016, 11, e0149470. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.; Klinngam, W.; Edman, M.C.; Hamm-Alvarez, S.F. Interferon-gamma treatment in vitro elicits some of the changes in Cathepsin S and antigen presentation characteristic of lacrimal glands and corneas from the NOD mouse model of Sjogren’s syndrome. PLoS ONE 2017, 12, e0184781. [Google Scholar] [CrossRef] [PubMed]
- Liu, W. On sample size determination of Dunnett’s procedure for comparing several treatments with a control. J. Stat. Plan. Inference 1997, 62, 255–261. [Google Scholar] [CrossRef]










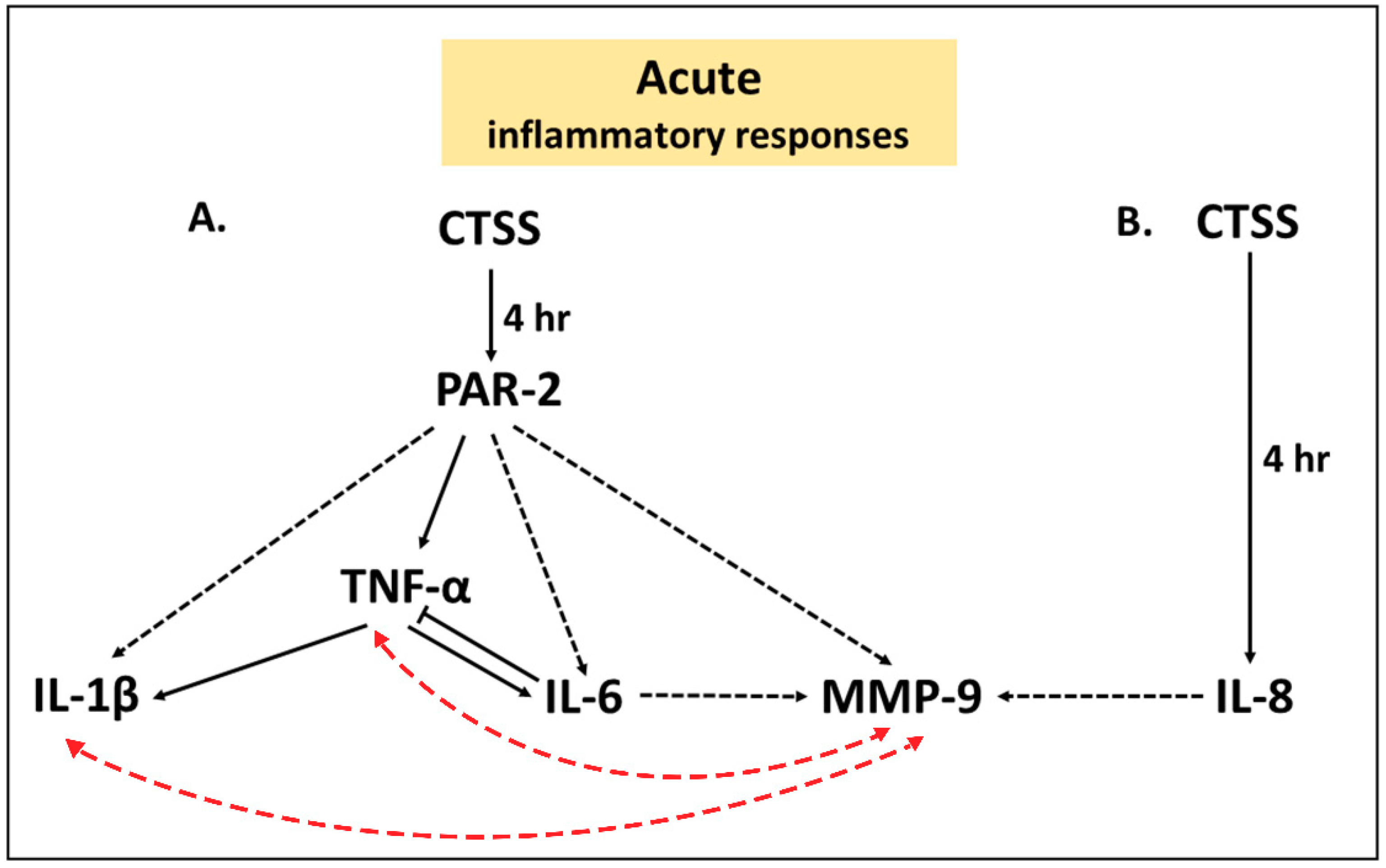
 ) represents a proposed inducible pathway, an one-way black dashed arrow (
) represents a proposed inducible pathway, an one-way black dashed arrow (  ) represents a possible one-way inducible pathway, a two-way red dashed arrow (
) represents a possible one-way inducible pathway, a two-way red dashed arrow (  ) represents a possible two-way inducible pathway and a solid line with blunt end (
) represents a possible two-way inducible pathway and a solid line with blunt end (  ) represents a negative feedback pathway.
) represents a proposed inducible pathway, an one-way black dashed arrow ( ) represents a possible one-way inducible pathway, a two-way red dashed arrow ( ) represents a possible two-way inducible pathway and a solid line with blunt end ( ) represents a negative feedback pathway.
) represents a negative feedback pathway.
) represents a proposed inducible pathway, an one-way black dashed arrow ( ) represents a possible one-way inducible pathway, a two-way red dashed arrow ( ) represents a possible two-way inducible pathway and a solid line with blunt end ( ) represents a negative feedback pathway.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Expression | Relative Expression | |||||
|---|---|---|---|---|---|---|
| 4 h of CTSS Treatment | 24 h of CTSS Treatment | |||||
| Scrambled siRNA | PAR-2 siRNA | p Value | Scrambled siRNA | PAR-2 siRNA | p Value | |
| IL-8 | 1.007 ± 0.084 | 1.109 ± 0.130 | ns | 1.029 ± 0.180 | 0.348 ± 0.057 | * |
| IL-6 | 1.007 ± 0.085 | 0.525 ± 0.079 | * | 1.000 ± 0.021 | 0.090 ± 0.007 | **** |
| IL-1β | 1.187 ± 0.493 | 0.546 ± 0.153 | ns | 1.032 ± 0.183 | 0.222 ± 0.068 | * |
| TNF-α | 1.048 ± 0.224 | 0.382 ± 0.083 | * | 1.007 ± 0.087 | 0.267 ± 0.040 | ** |
| CTSS | 1.012 ± 0.115 | 1.156 ± 0.263 | ns | 1.017 ± 0.131 | 0.289 ± 0.059 | ** |
| MMP-9 | 1.050 ± 0.243 | 0.723 ± 0.112 | ns | 1.011 ± 0.104 | 0.099 ± 0.034 | ** |
| Protein Expression | Protein Expression in Cell Lysates (pg/mg protein) | |||||
|---|---|---|---|---|---|---|
| 4 h of CTSS Treatment | 24 h of CTSS Treatment | |||||
| Scrambled siRNA | PAR-2 siRNA | p Value | Scrambled siRNA | PAR-2 siRNA | p Value | |
| Cell Culture Medium | ||||||
| IL-8 | 18759 ± 61.6 | 19800 ± 472.2 | ns | 15254 ± 445 | 20377 ± 289.2 | ns |
| IL-6 a | 79003 ± 12649 | 26608 ± 9138 | * | 42730 ± 2075 | 17810 ± 844.4 | *** |
| IL-1β a | 1.240 ± 0.1052 | 0.738 ± 0.095 | * | 1.826 ± 0.235 | 1.302 ± 0.072 | ns |
| TNF-α | 105.8 ± 7.221 | 43.52 ± 1.785 | ** | 44.64 ± 2.173 | 22.09 ± 0.662 | *** |
| MMP-9 | 10834 ± 881 | 5613 ± 48.90 | ** | 34666 ± 836 | 19702 ± 3355 | * |
| Cell Lysates | ||||||
| IL-8 | 147.5 ± 16.61 | 115.7 ± 10.92 | ns | 108.8 ± 18.13 | 113.8 ± 21.64 | ns |
| IL-6 | 8.784 ± 0.560 | 3.862 ± 1.133 | * | 3.155 ± 0.746 | 1.548 ± 0.231 | ns |
| IL-1β | 138.0 ± 7.153 | 94.75 ± 15.88 | ns | 62.99 ± 11.64 | 36.33 ± 7.632 | ns |
| TNF-α | 0.836 ± 0.056 | 0.778 ± 0.145 | ns | 0.794 ± 0.039 | 0.669 ± 0.117 | ns |
| CTSS | 25.16 ± 5.543 | 33.53 ± 6.185 | ns | 35.60 ± 6.494 | 34.30 ± 8.386 | ns |
| MMP-9 | 6160 ± 329.0 | 3322 ± 309.5 | ** | 10628 ± 1137 | 5980 ± 1390 | ns |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klinngam, W.; Fu, R.; Janga, S.R.; Edman, M.C.; Hamm-Alvarez, S.F. Cathepsin S Alters the Expression of Pro-Inflammatory Cytokines and MMP-9, Partially through Protease—Activated Receptor-2, in Human Corneal Epithelial Cells. Int. J. Mol. Sci. 2018, 19, 3530. https://doi.org/10.3390/ijms19113530
Klinngam W, Fu R, Janga SR, Edman MC, Hamm-Alvarez SF. Cathepsin S Alters the Expression of Pro-Inflammatory Cytokines and MMP-9, Partially through Protease—Activated Receptor-2, in Human Corneal Epithelial Cells. International Journal of Molecular Sciences. 2018; 19(11):3530. https://doi.org/10.3390/ijms19113530
Chicago/Turabian StyleKlinngam, Wannita, Runzhong Fu, Srikanth R. Janga, Maria C. Edman, and Sarah F. Hamm-Alvarez. 2018. "Cathepsin S Alters the Expression of Pro-Inflammatory Cytokines and MMP-9, Partially through Protease—Activated Receptor-2, in Human Corneal Epithelial Cells" International Journal of Molecular Sciences 19, no. 11: 3530. https://doi.org/10.3390/ijms19113530
APA StyleKlinngam, W., Fu, R., Janga, S. R., Edman, M. C., & Hamm-Alvarez, S. F. (2018). Cathepsin S Alters the Expression of Pro-Inflammatory Cytokines and MMP-9, Partially through Protease—Activated Receptor-2, in Human Corneal Epithelial Cells. International Journal of Molecular Sciences, 19(11), 3530. https://doi.org/10.3390/ijms19113530