Evolution and Expression Characteristics of Receptor-Like Cytoplasmic Protein Kinases in Maize, Rice and Arabidopsis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Identification of RLCKs in Maize, Rice and Arabidopsis
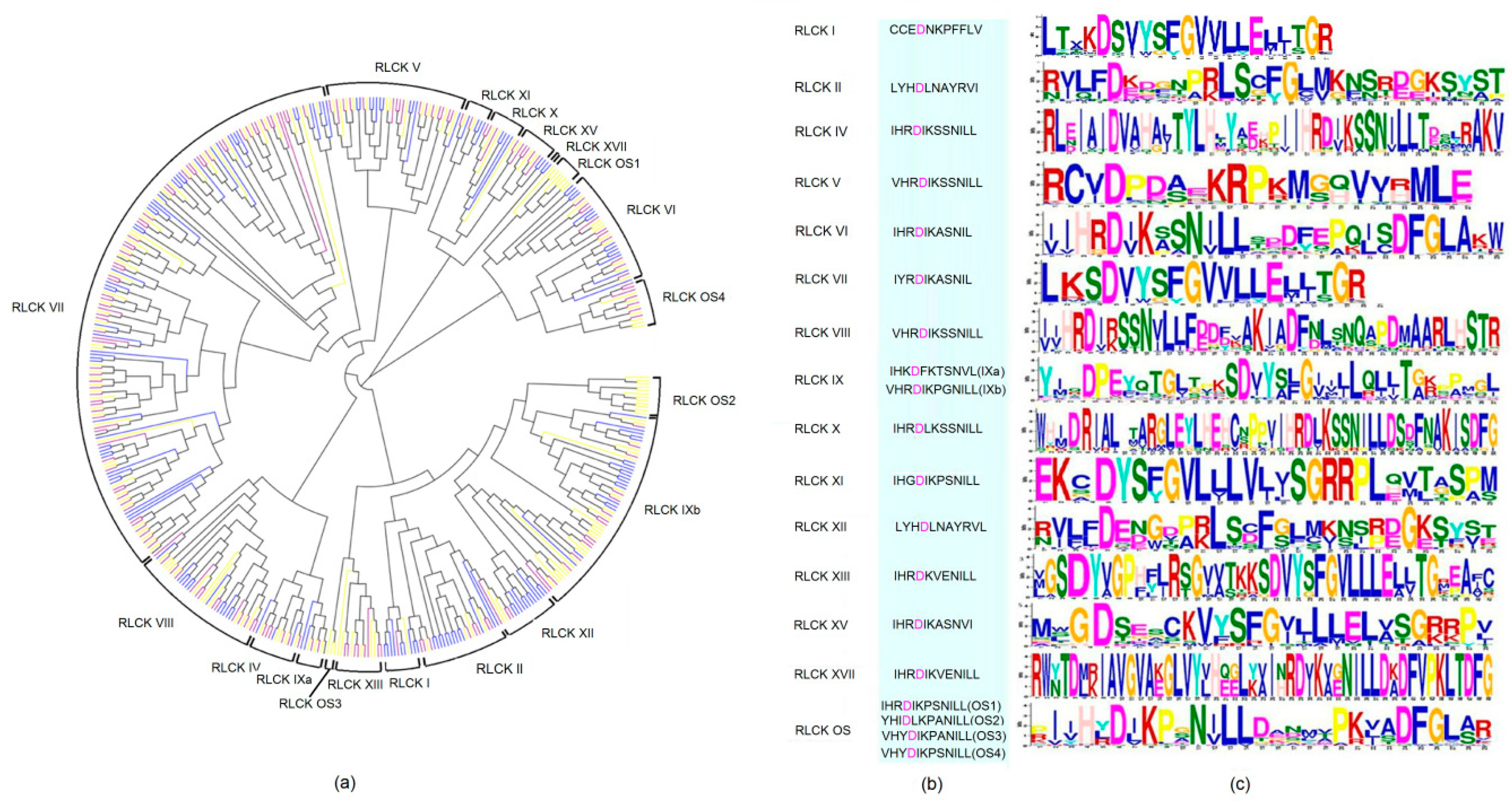
2.2. Conservative Evolution of RLCKs in Maize, Rice and Arabidopsis
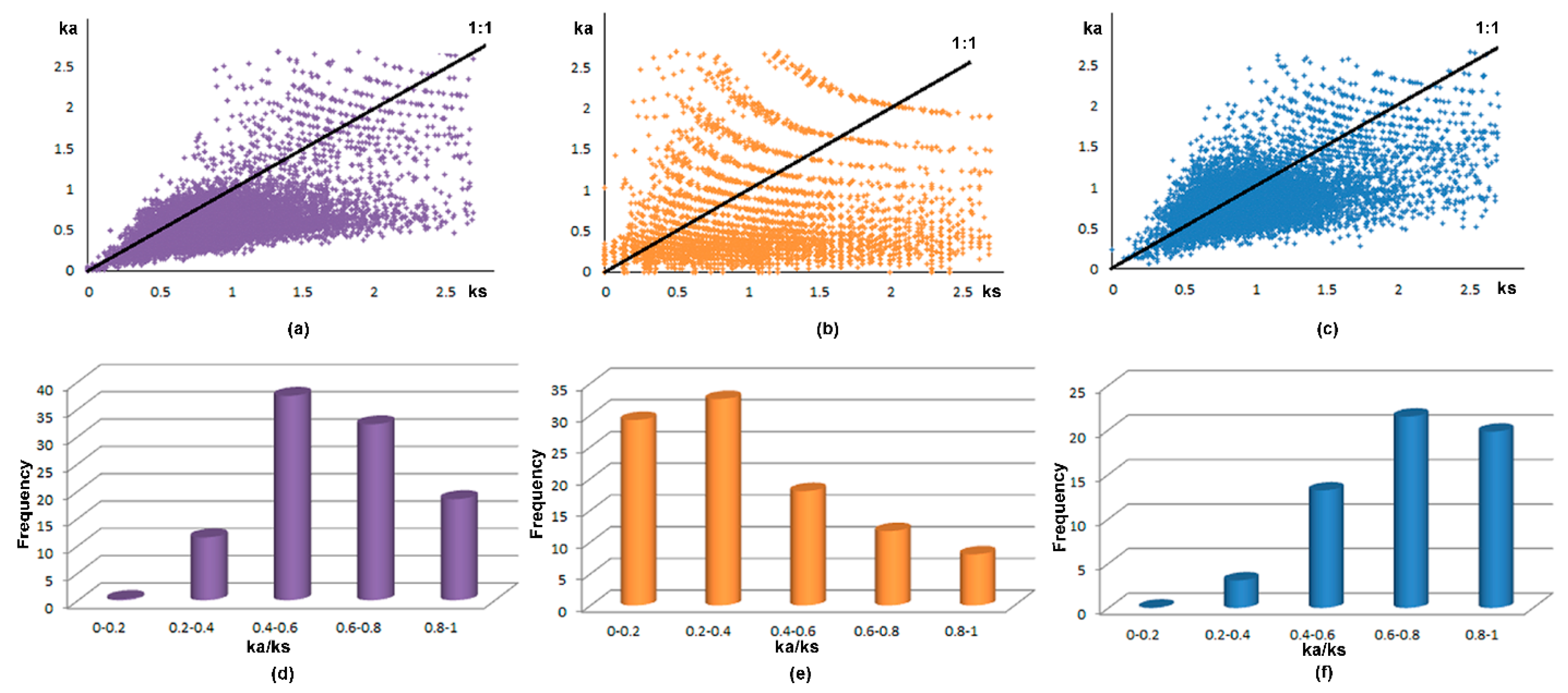
2.3. The Evolution Selection of RLCKs in Maize, Rice and Arabidopsis
2.4. Segmental Duplication of RLCKs in Maize, Rice and Arabidopsis
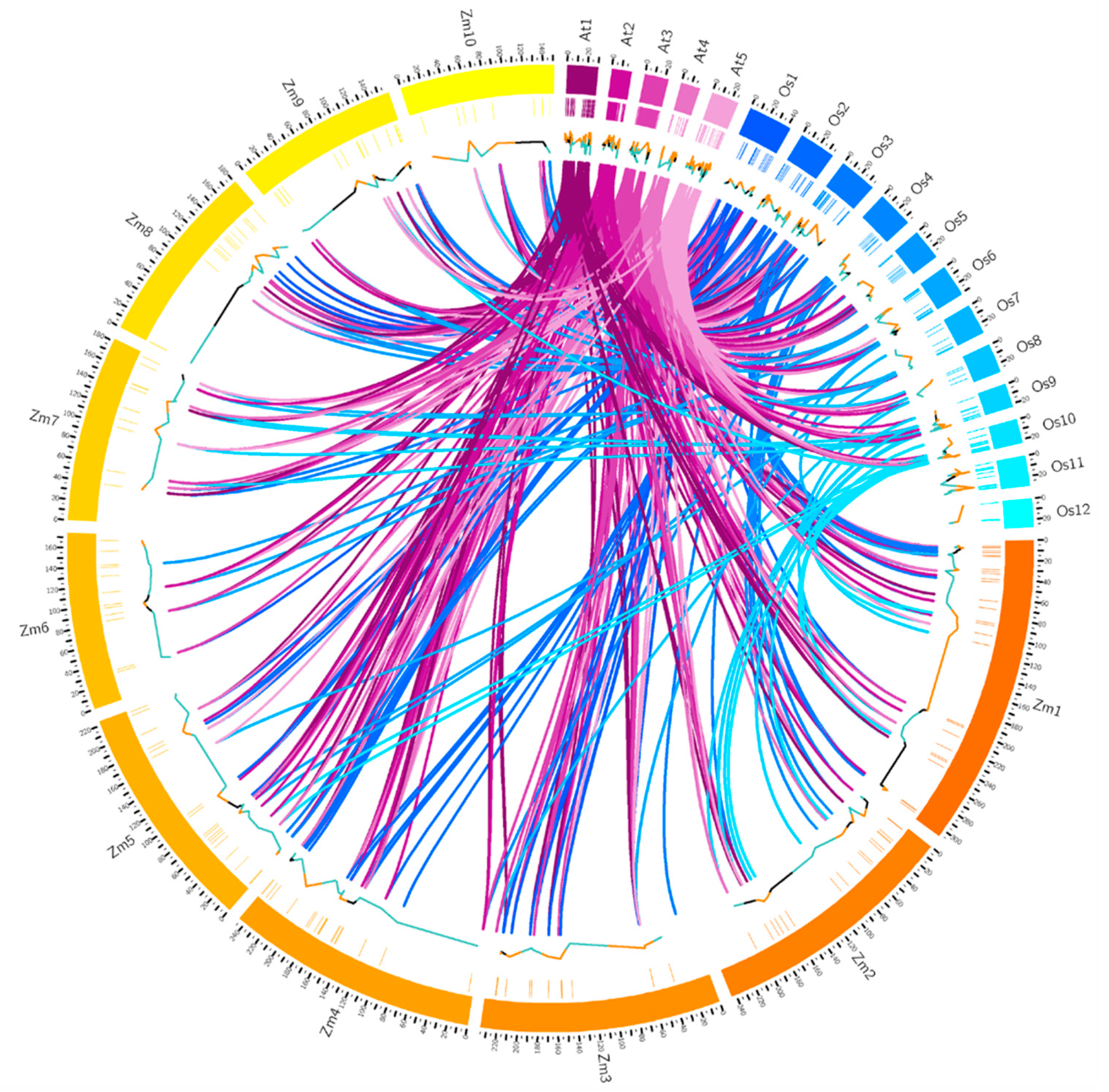
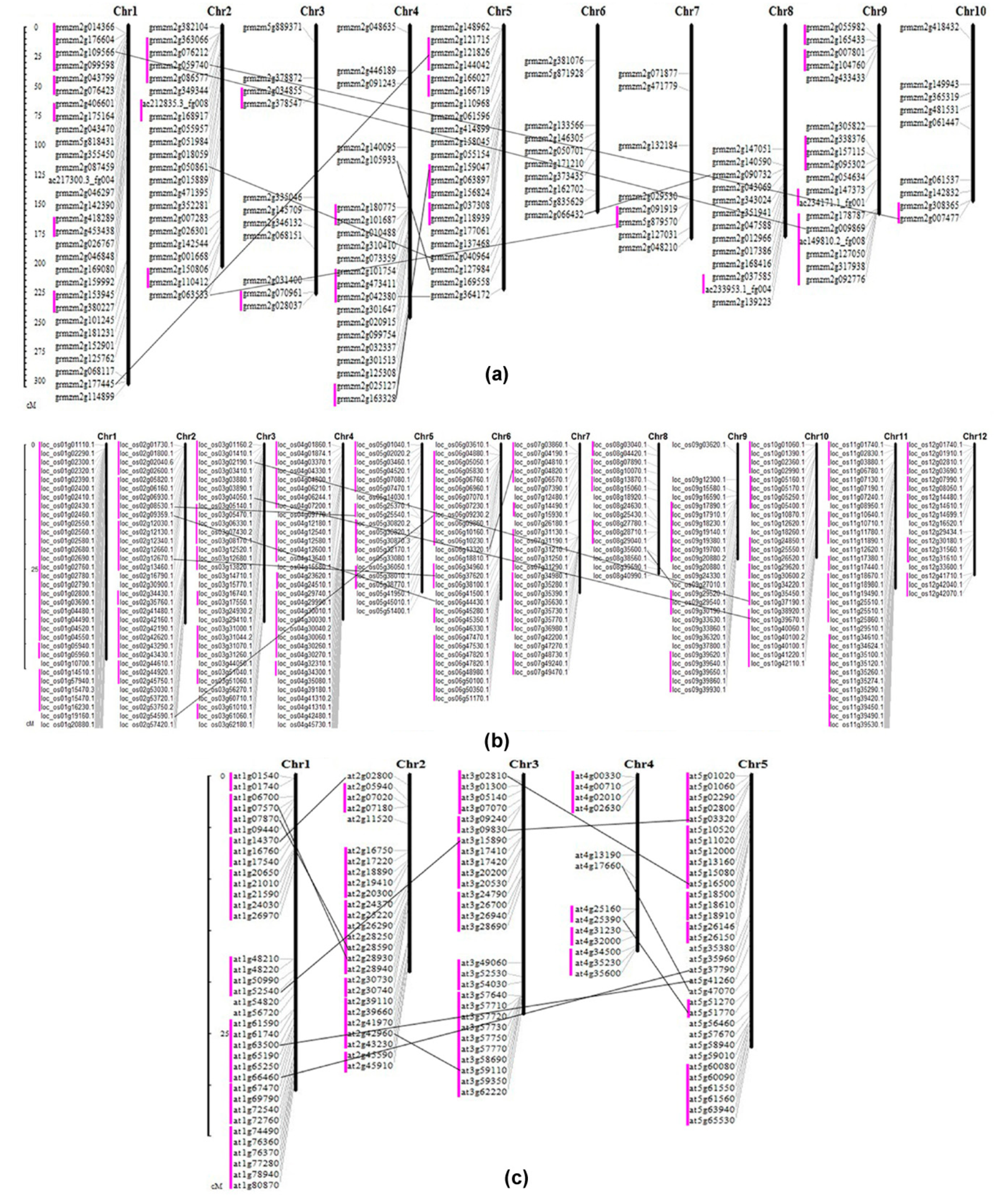
2.5. The Distribution and Synteny of RLCK Genes in Maize, Rice and Arabidopsis
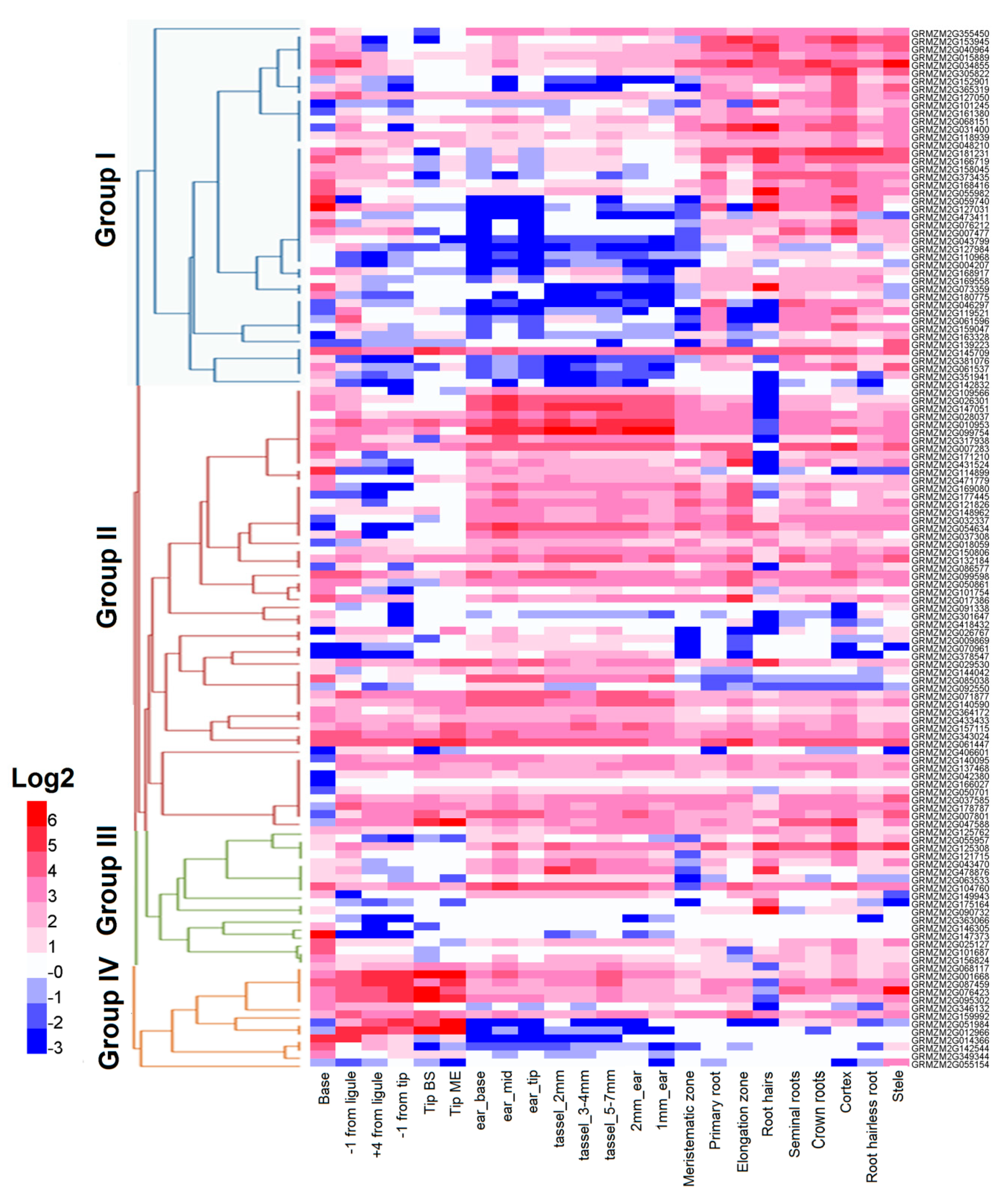
2.6. Dynamic Expression of RLCK Genes in Maize
3. Discussion
4. Materials and Methods
4.1. Identification and Protein Domain Prediction of RLCK Genes
4.2. The Phylogram and Selection of RLCKs
4.3. Distribution and Synteny of RLCK Genes
4.4. Differential Expression of Maize RLCK Genes
Supplementary Materials
Author contribution
Funding
Conflicts of Interest
References
- Morillo, S.A.; Tax, F.E. Functional analysis of receptor-like kinases in monocots and dicots. Curr. Opin. Plant Bio. 2006, 9, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Shiu, S.H.; Karlowski, W.M.; Pan, R.; Tzeng, Y.H.; Mayer, K.F. Comparative analysis of the receptor-like kinase family in Arabidopsis and rice. Plant Cell. 2004, 16, 1220–1234. [Google Scholar] [CrossRef] [PubMed]
- Vij, S.; Giri, J.; Dansana, P.K.; Kapoor, S.; Tyagi, A.K. The receptor-like cytoplasmic kinase (OsRLCK) gene family in rice: Organization, phylogenetic relationship, and expression during development and stress. Mol. Plant. 2008, 1, 732–750. [Google Scholar] [CrossRef] [PubMed]
- Jurca, M.E.; Bottka, S.; Fehér, A. Characterization of a family of Arabidopsis receptor-like cytoplasmic kinases (RLCK class VI). Plant Cell Rep. 2008, 27, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, M.M.; Pinto, S.; Kluth, J.; Wienand, U.; Lorbiecke, R. The PTI1-like kinase ZmPti1a from maize (Zea mays L.) co-localizes with callose at the plasma membrane of pollen and facilitates a competitive advantage to the male gametophyte. BMC Plant Biol. 2006, 6, 22. [Google Scholar] [CrossRef] [PubMed]
- Smet, I.D.; Vos, U.; Jürgens, G. Receptor-like kinases shape the plant. Nat. Cell Biol. 2009, 11, 1166–1173. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.C.; Zhang, R. Relationship of a putative receptor protein kinase from maize to the s-locus glycoproteins of brassica. Nature 1990, 345, 743–746. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Chaudhuri, S.; Yang, L.; Chen, Y.; Poovaiah, B.W. Calcium/calmodulin up-regulates a cytoplasmic receptor-like kinase in plants. J. Biol. Chem. 2004, 279, 42552–42559. [Google Scholar] [CrossRef] [PubMed]
- Dissanayake, K.; Castillo, C.; Takasaki, T.; Nakanishi, T.; Norioka, N. Molecular cloning, functional expression and characterization of two serine/threonine-specific protein kinases from nicotiana tabacum pollen. Sex. Plant Reprod. 2004, 17, 165–175. [Google Scholar] [CrossRef]
- Wang, S.; Sun, J.; Fan, F.; Tan, Z.; Zou, Y.; Lu, D. A Xanthomonas oryzae pv. oryzae effector, XopR, associates with receptor-like cytoplasmic kinases and suppresses PAMP-triggered stomatal closure. Sci. China Life Sci. 2016a, 59, 897–905. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wu, G.; Peng, C.; Zhou, X.; Li, W. The Receptor-Like Cytoplasmic Kinase OsRLCK102 Regulates XA21-Mediated Immunity and Plant Development in Rice. Plant Mol. Bio. Rep. 2016b, 34, 628–637. [Google Scholar] [CrossRef]
- Sun, X.; Sun, M.; Luo, X.; Ding, X.; Ji, W.; Cai, H.; Bai, X.; Liu, X.; Zhu, Y. A Glycine soja ABA-responsive receptor-like cytoplasmic kinase, GsRLCK, positively controls plant tolerance to salt and drought stresses. Planta 2013, 237, 1527–1545. [Google Scholar] [CrossRef] [PubMed]
- Panchy, N.; Lehti-Shiu, M.; Shiu, S.H. Evolution of gene duplication in plants. Plant Physiol. 2016, 171, 2294–2316. [Google Scholar] [CrossRef] [PubMed]
- Baumgarten, A.; Cannon, S.; Spangler, R.; May, G. Genome-level evolution of resistance genes in Arabidopsis thaliana. Genetics 2003, 165, 309–319. [Google Scholar] [PubMed]
- Eveland, A.L.; Goldshmidt, A.; Pautler, M.; Morohashi, K.; Liseronmonfils, C. Regulatory modules controlling maize inflorescence architecture. Genome Res. 2014, 24, 431–443. [Google Scholar] [CrossRef] [PubMed]
- Kondrashov, F.A. Gene duplication as a mechanism of genomic adaptation to a changing environment. Proc. Biol. Sci. 2012, 279, 5048–5057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galtier, N. Gene conversion drives GC content evolution in mammalian histones. Trends Genet. 2003, 19, 65–68. [Google Scholar] [CrossRef]
- Zhao, T.; Holmer, R.; Bruijn, S.D.; Angenent, G.C.; Van, H.A.; Schranz, M.E. Phylogenomic Synteny Network Analysis of MADS-Box Transcription Factor Genes Reveals Lineage-Specific Transpositions, Ancient Tandem Duplications, and Deep Positional Conservation. Plant Cell 2017, 29, 1278–1292. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Bowers, J.E.; Wang, X.; Ming, R.; Alam, M. Synteny and collinearity in plant genomes. Science 2008, 320, 486–488. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Conery, J.S. The evolutionary fate and consequences of duplicate genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef] [PubMed]
- Gaut, B.S.; Morton, B.R.; McCaig, B.C.; Clegg, M.T. Substitution rate comparisons between grasses and palms: synonymous rate differences at the nuclear gene Adh parallel rate differences at the plastid gene rbcL. Proc. Natl. Acad. Sci. USA 1996, 93, 10274–10279. [Google Scholar] [CrossRef] [PubMed]
- Koch, M.A.; Haubold, B.; Mitchell-Olds, T. Comparative evolutionary analysis of chalcone synthase and alcohol dehydrogenase loci in Arabidopsis, Arabis, and related genera (Brassicaceae). Mol. Biol. Evol. 2000, 17, 1483–1498. [Google Scholar] [CrossRef] [PubMed]
- Kozik, A.; Kochetkova, E.; Michelmore, R. GenomePixelizer-a visualization program for comparative genomics within and between species. Bioinformatics 2002, 18, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Higgins, D.G.; Bleasby, A.J.; Fuchs, R. CLUSTAL V: Improved software for multiple sequence alignment. CABIOS 1992, 8, 189–191. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Wei, K.; Wang, Y.; Xie, D. Identification and expression profile analysis of the protein kinase gene superfamily in maize development. Mol. Breed. 2014, 33, 155–172. [Google Scholar] [CrossRef]
- Wang, D.X.; Wang, H.; Irfan, M.; Fan, M.X.; Lin, F. Structure and evolution analysis of pollen receptor-like kinase in Zea mays and Arabidopsis thaliana. Comput. Biol. Chem. 2014, 51, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Juretic, N.; Hoen, D.R.; Huynh, M.L.; Harrison, P.M.; Bureau, T.E. The evolutionary fate of MULE-mediated duplications of host gene fragments in rice. Genome Res. 2005, 15, 1292–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leister, D. Tandem and segmental gene duplication and recombination in the evolution of plant disease resistance genes. Trends Genet. 2004, 20, 116–122. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, M.; Ma, W.; Liu, C.; Zhang, C.; Wu, S.; Chen, M.; Liu, K.; Cai, F.; Lin, F. Evolution and Expression Characteristics of Receptor-Like Cytoplasmic Protein Kinases in Maize, Rice and Arabidopsis. Int. J. Mol. Sci. 2018, 19, 3680. https://doi.org/10.3390/ijms19113680
Fan M, Ma W, Liu C, Zhang C, Wu S, Chen M, Liu K, Cai F, Lin F. Evolution and Expression Characteristics of Receptor-Like Cytoplasmic Protein Kinases in Maize, Rice and Arabidopsis. International Journal of Molecular Sciences. 2018; 19(11):3680. https://doi.org/10.3390/ijms19113680
Chicago/Turabian StyleFan, Mingxia, Wenjuan Ma, Chen Liu, Chunyu Zhang, Suwen Wu, Meiming Chen, Kuichen Liu, Fengchun Cai, and Feng Lin. 2018. "Evolution and Expression Characteristics of Receptor-Like Cytoplasmic Protein Kinases in Maize, Rice and Arabidopsis" International Journal of Molecular Sciences 19, no. 11: 3680. https://doi.org/10.3390/ijms19113680
APA StyleFan, M., Ma, W., Liu, C., Zhang, C., Wu, S., Chen, M., Liu, K., Cai, F., & Lin, F. (2018). Evolution and Expression Characteristics of Receptor-Like Cytoplasmic Protein Kinases in Maize, Rice and Arabidopsis. International Journal of Molecular Sciences, 19(11), 3680. https://doi.org/10.3390/ijms19113680