Differential Responses of OsMPKs in IR56 Rice to Two BPH Populations of Different Virulence Levels


Abstract
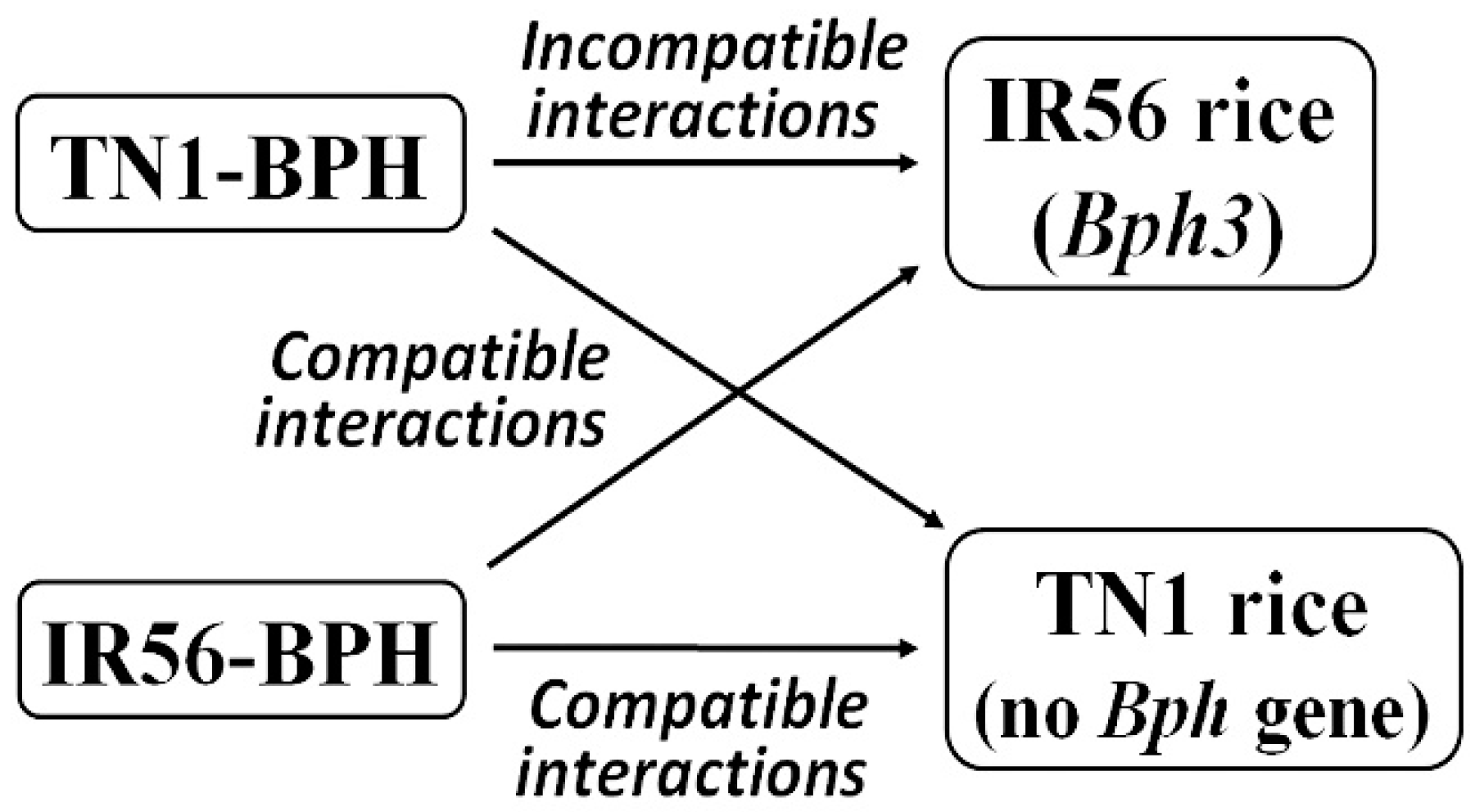
:1. Introduction
2. Results
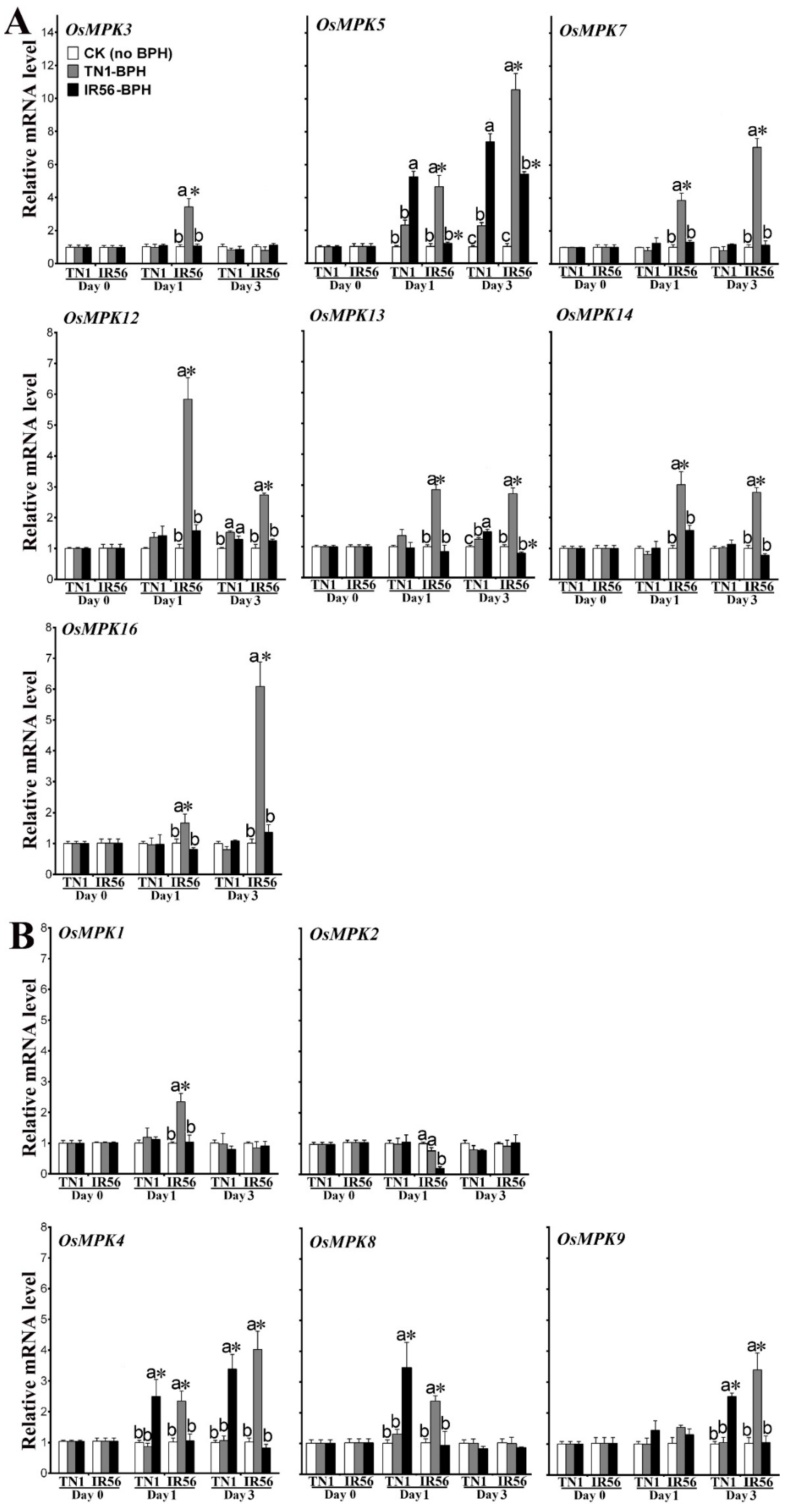
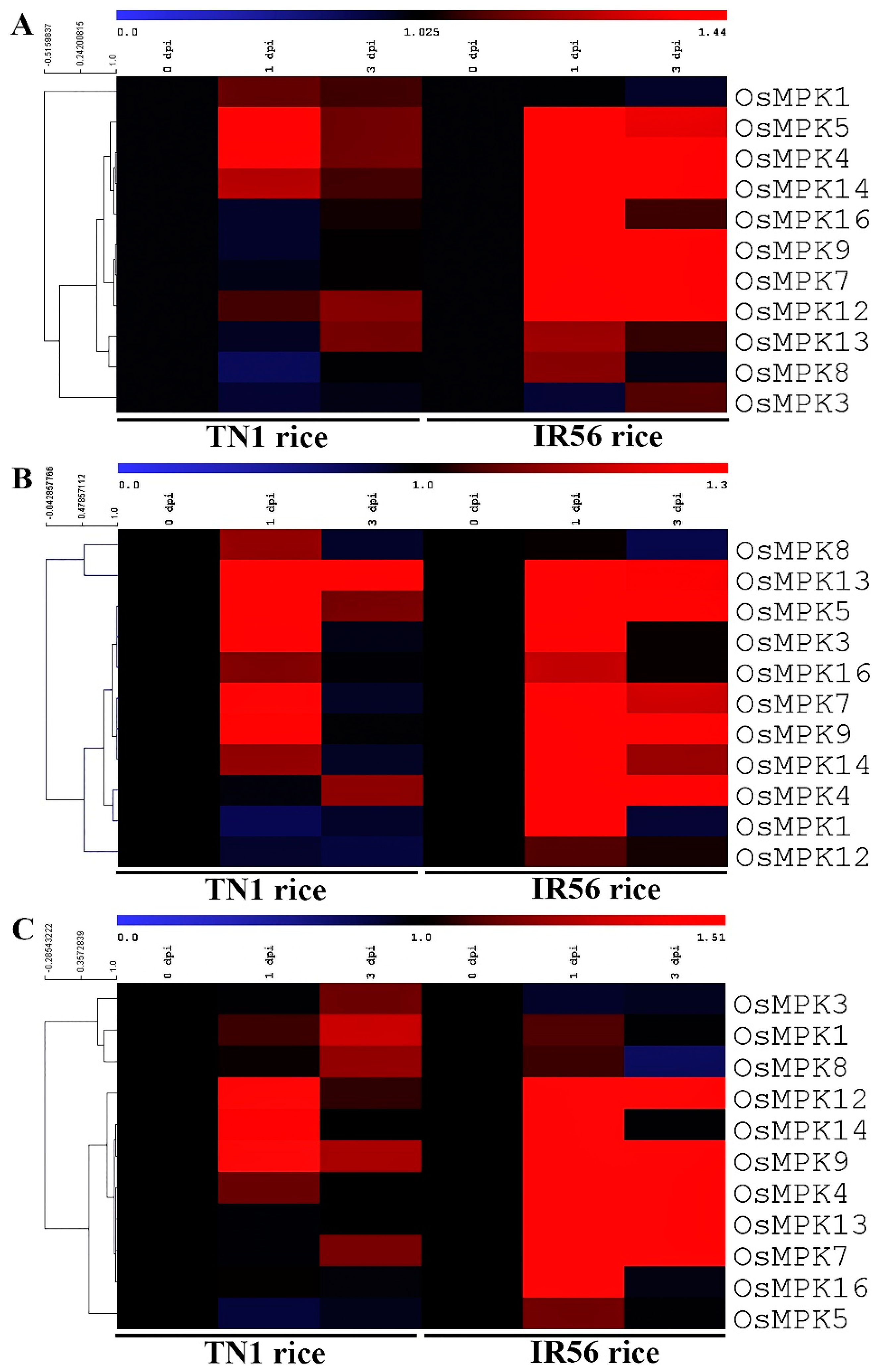
2.1. Expression Profiling of OsMPKs in Response to BPH Infestations
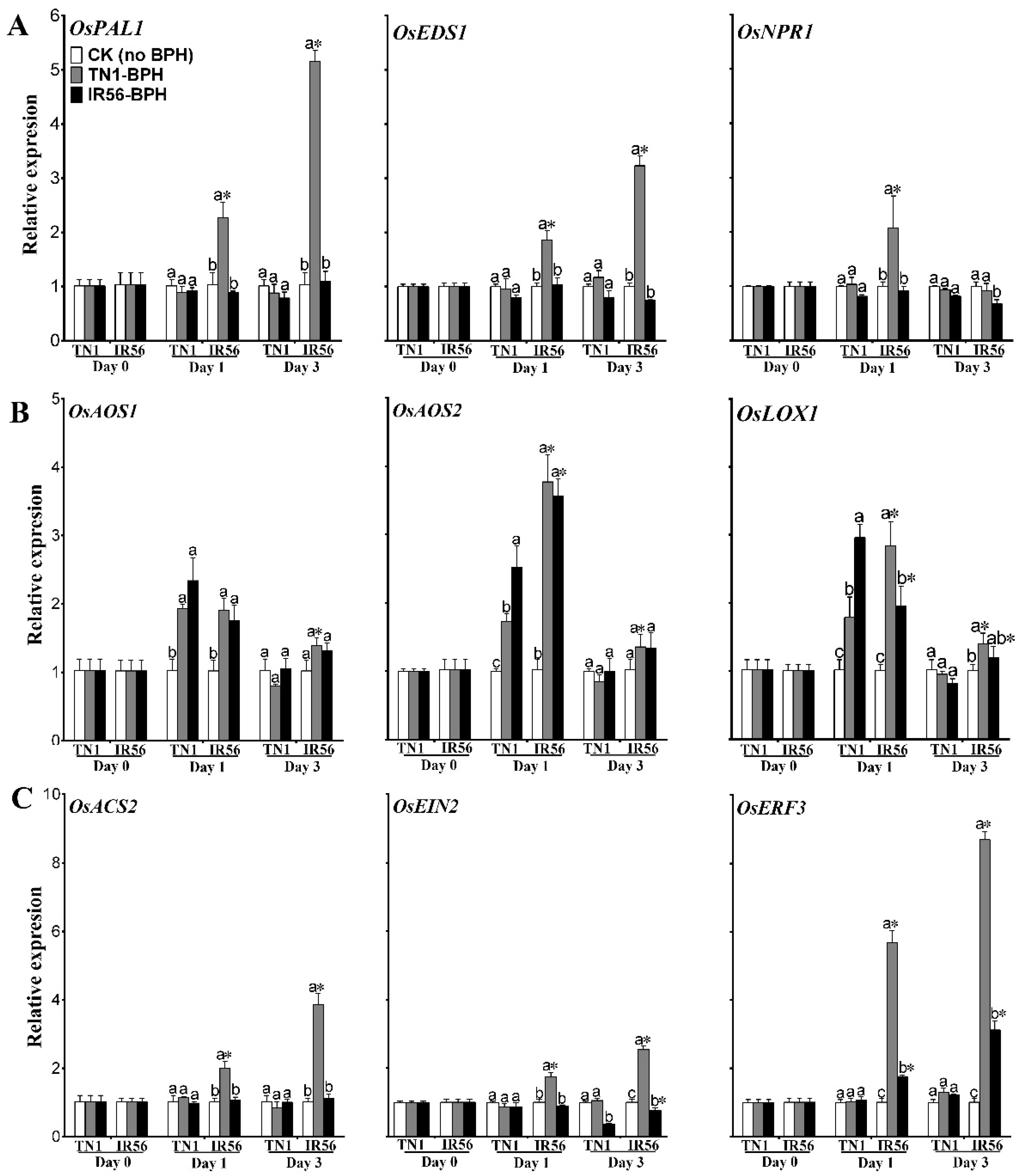
2.2. Induced Expression of Genes Associated with Phytohormone Signaling and Phenylpropanoid Synthesis under BPH Infestation
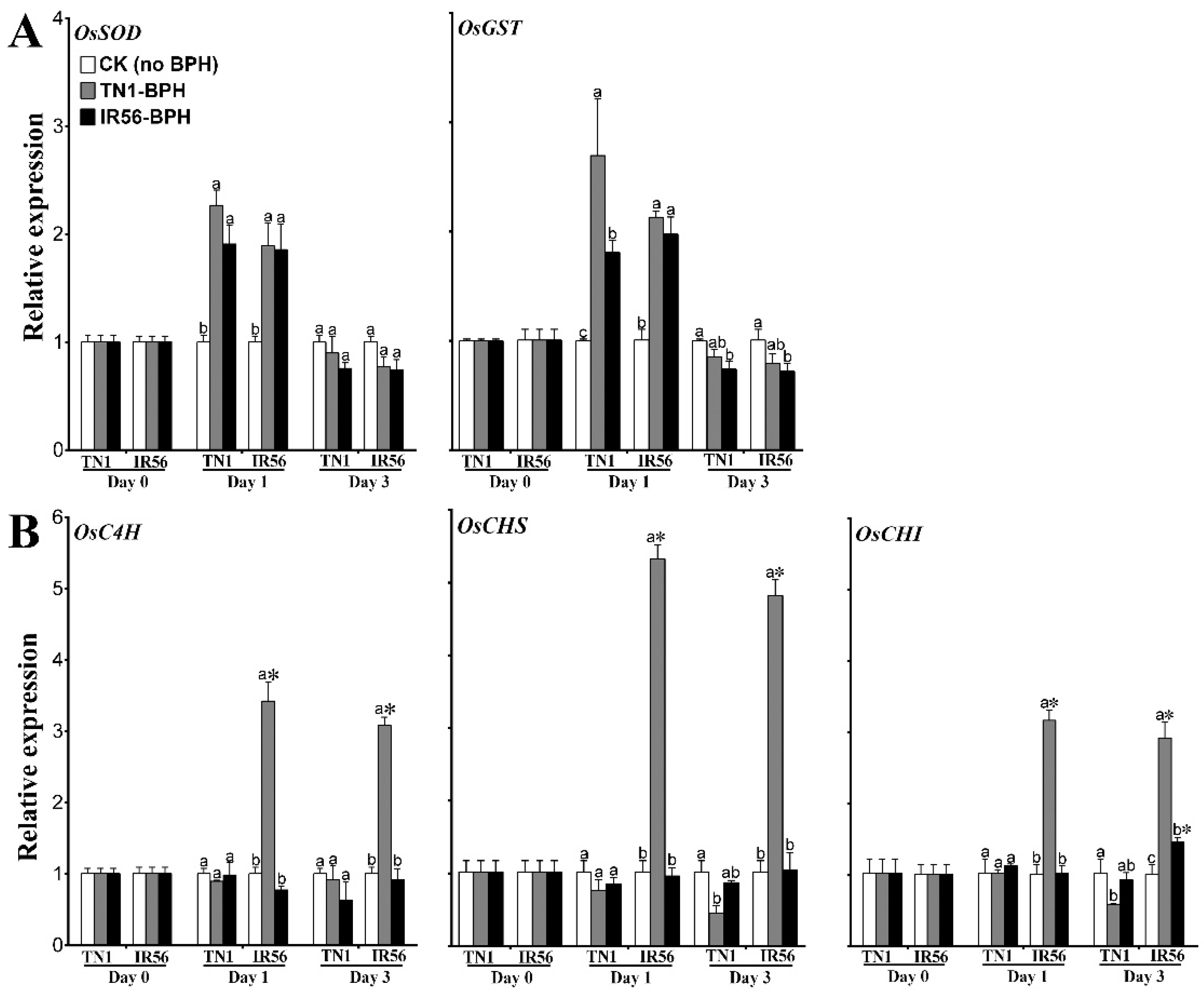
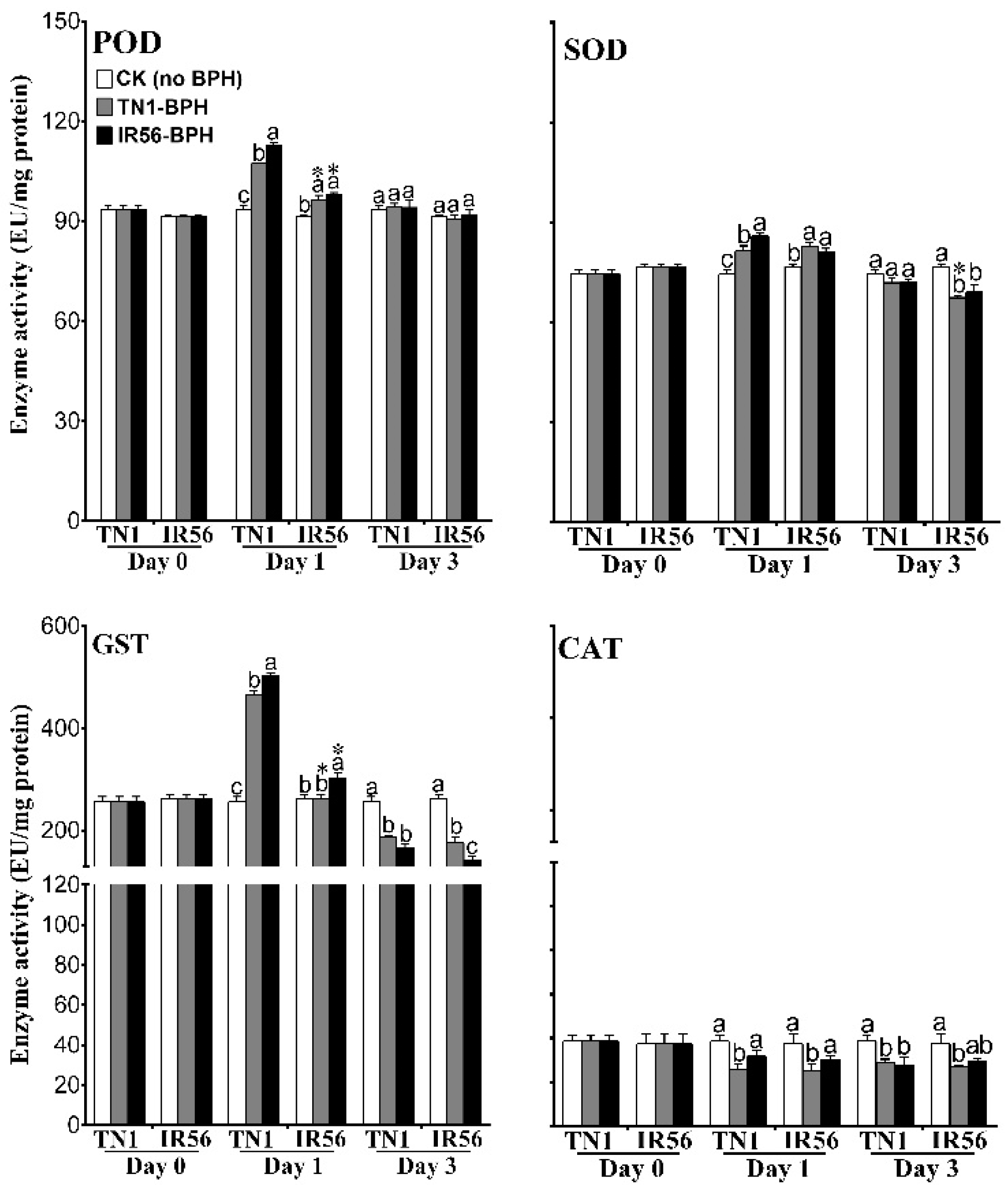
2.3. Expression Profiling and Enzyme Activity of ROS-Signaling Genes under BPH Infestation
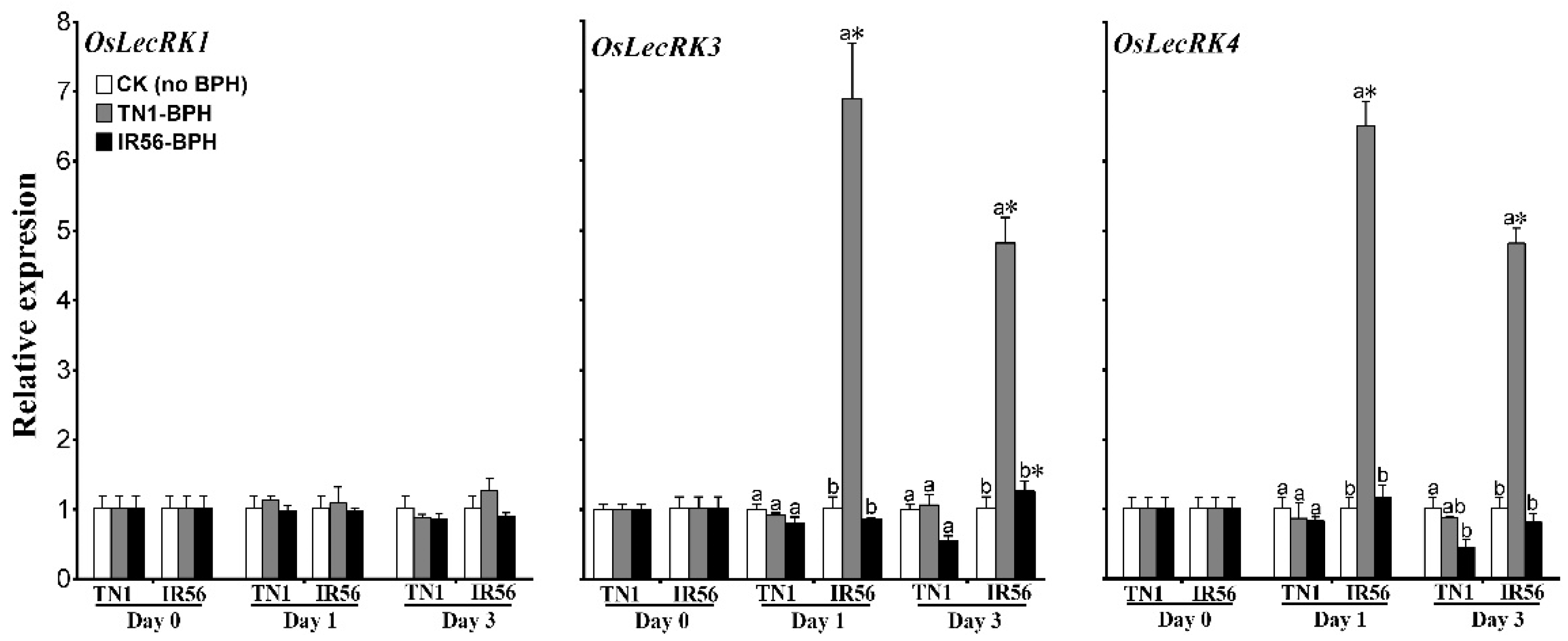
2.4. Differential Transcription of OsLecRKs in the Two Rice Varieties under BPH Infestation
2.5. Expression Modulation of the BPH-Induced OsMPKs in Response to the Exogenous Treatment of Phytohormones
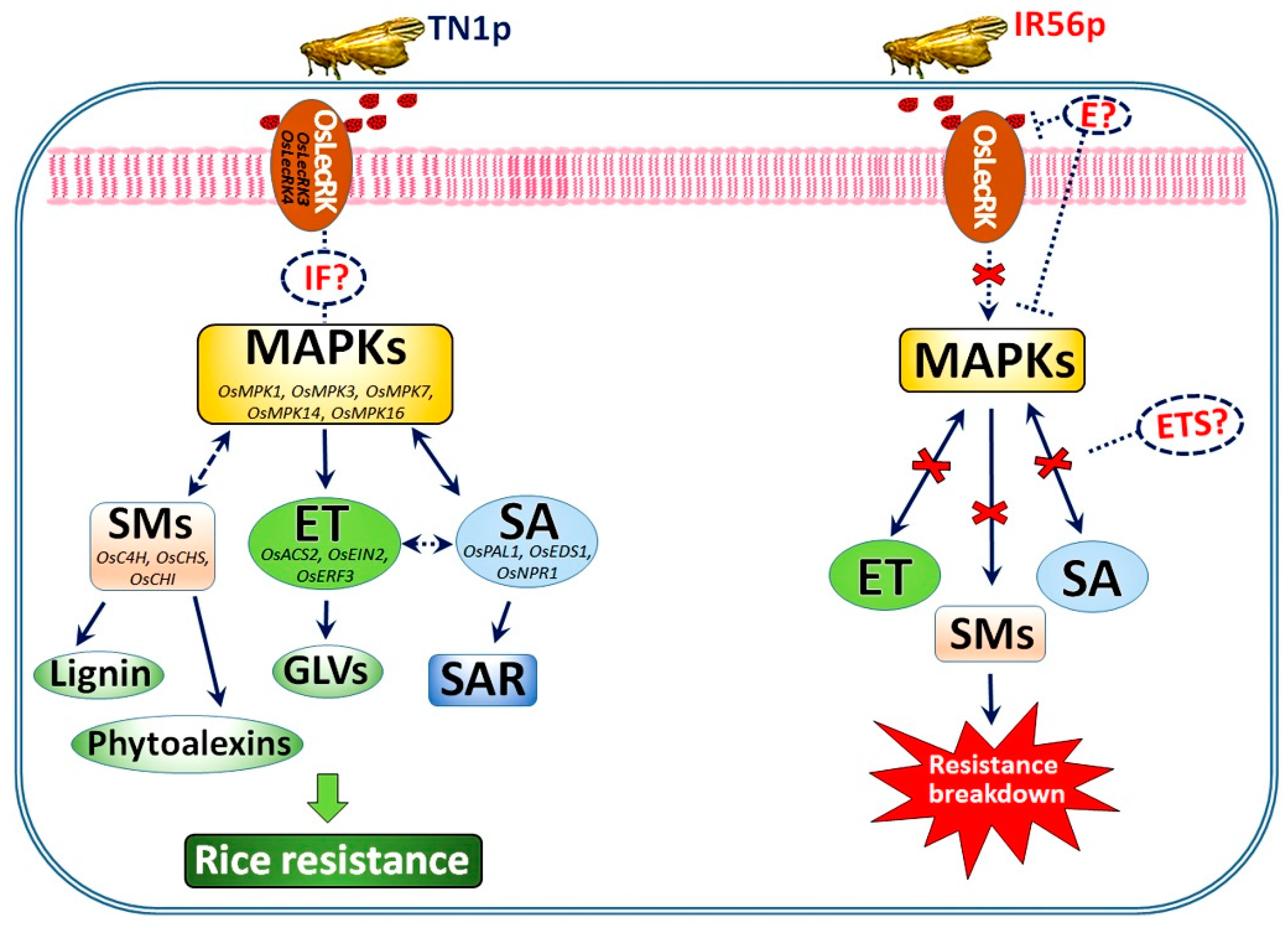
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Insect Materials
4.3. BPH Bioassays
4.4. Chemical Treatments
4.5. RNA Isolation and cDNA Synthesis
4.6. Quantitative Real-Time PCR (qPCR)
4.7. Enzyme Activity Assays
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BPH | Brown planthopper |
| TN1 | Taichung Native 1 |
| MAPK | Mitogen-activated protein kinase |
| LecRK | Lectin receptor kinase |
| ANOVA | Analysis of variance |
| SA | Salicylic acid |
| JA | Jasmonic acid |
| ET | Ethylene |
| ROS | Reactive oxygen species |
| GST | Glutathione S–transferase |
| CAT | Catalase |
| SOD | Superoxide dismutase |
| POD | Peroxidase |
| PAL1 | Phenylalanine ammonia lyase 1 |
| EDS1 | Enhanced disease susceptibility 1 |
| NPR1 | Nonexpresser of pathogenesis-related genes 1 |
| AOS1 | Allene oxide synthase 1 |
| AOS2 | allene oxide synthase 2 |
| LOX1 | Lipoxygenase 1 |
| ACS2 | 1-aminocyclopropane-1-carboxylate synthase 2 |
| EIN2 | Ethylene-insensitive gene 2 |
| ERF3 | Ethylene-responsive transcription factor |
| C4H | Cinnamate-4-hydroxylase |
| CHS | Chalcone synthase |
| CHI | Chalcone isomerase |
References
- Cheng, X.; Zhu, L.; He, G. Towards understanding of molecular interactions between rice and the brown planthopper. Mol. Plant 2013, 6, 621–634. [Google Scholar] [CrossRef] [PubMed]
- Pathak, M.D.; Cheng, C.H.; Fortuno, M.E. Resistance to Nephotettix impicticeps and Nilaparvata lugens in varieties of rice. Nature 1969, 223, 502–504. [Google Scholar] [CrossRef]
- Athwal, D.S.; Pathak, M.D.; Bacalangco, E.H.; Pura, C.D. Genetics of resistance to brown planthoppers and green leafhoppers in Oryza sativa L.1. Crop Sci. 1971, 11, 747–750. [Google Scholar] [CrossRef]
- Sogawa, K. Planthopper outbreaks in different paddy ecosystems in Asia: Man-made hopper plagues that threatened the green revolution in rice. In Rice Planthoppers: Ecology, Management, Socio Economics and Policy; Heong, K.L., Cheng, J., Escalada, M.M., Eds.; Springer: Dordrecht, The Netherlands, 2015. [Google Scholar] [CrossRef]
- Khush, G.S. Genetics of and breeding for resistance to the brown planthopper. In Brown Planthopper: Threat to Rice Production in Asia; International Rice Research Institute: Los Baños, Philippines, 1979; pp. 321–332. [Google Scholar]
- Mochida, O.; Heinrichs, E. Strategies for the development of an integrated approach to rice brown planthopper control. In Rice Research Strategies for the Future; International Rice Research Institute: Los Baños, Philippines, 1982; p. 99. [Google Scholar]
- Liu, Y.; Wu, H.; Chen, H.; Liu, Y.; He, J.; Kang, H.; Sun, Z.; Pan, G.; Wang, Q.; Hu, J.; et al. A gene cluster encoding lectin receptor kinases confers broad-spectrum and durable insect resistance in rice. Nat. Biotechnol. 2014, 33, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Sōgawa, K. The rice brown olanthopper: Feeding physiology and host plant interactions. Annu. Rev. Entomol. 1982, 27, 49–73. [Google Scholar] [CrossRef]
- Zheng, Y.; He, J.; Wan, P.; Lai, F.; Sun, Y.; Lin, J.; Fu, Q. Virulence characteristics of Nilaparvata lugens (Stål) reared on resistant rice variety IR56. Chin. J. Rice Sci. 2016, 30, 552–558. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, L.; Liu, Y.; Dai, H.; He, J.; Kang, H.; Pan, G.; Huang, J.; Qiu, Z.; Wang, Q.; et al. Marker assisted pyramiding of two brown planthopper resistance genes, Bph3 and Bph27(t), into elite rice Cultivars. Rice 2016, 9, 27. [Google Scholar] [CrossRef] [PubMed]
- International Rice Research Institute. Annual Report for 1975; International Rice Research Institute: Los Baños, Philippines, 1976; p. 479. [Google Scholar]
- Fu, Q.; Zhang, Z.; Hu, C.; Lai, F. The differentiation of amino acid requirements in three host-related populations of the brown planthopper, Nilaparvata lugens (Stål). Entomol. Sin. 2001, 8, 361–369. [Google Scholar]
- Han, Y.; Wu, C.; Yang, L.; Zhang, D.; Xiao, Y. Resistance to Nilaparvata lugens in rice lines introgressed with the resistance genes Bph14 and Bph15 and related resistance types. PLoS ONE 2018, 13, e0198630. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.; Sarao, P.S.; Bhatia, D.; Neelam, K.; Kaur, A.; Mangat, G.S.; Brar, D.S.; Singh, K. High-resolution genetic mapping of a novel brown planthopper resistance locus, Bph34 in Oryza sativa L. X Oryza nivara (Sharma & Shastry) derived interspecific F2 population. Theor. Appl. Genet. 2018, 131, 1163–1171. [Google Scholar] [CrossRef] [PubMed]
- Nanda, S.; Rout, E.; Joshi, R.K. Curcuma longa mitogen-activated protein kinase 6 (ClMPK6) stimulates the defense response pathway and enhances the resistance to necrotrophic fungal infection. Plant Mol. Biol. Rep. 2016, 34, 886–898. [Google Scholar] [CrossRef]
- Hettenhausen, C.; Schuman, M.C.; Wu, J. MAPK signaling: A key element in plant defense response to insects. Insect Sci. 2014, 22, 157–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Song, N.; Zhang, Q.; Wang, N.; Kang, Z. TaMAPK4 acts as a positive regulator in defense of wheat stripe-rust infection. Front. Plant Sci. 2018, 9, 152. [Google Scholar] [CrossRef] [PubMed]
- Husaini, A.M.; Sakina, A.; Cambay, S.R. Host–pathogen interaction in Fusarium oxysporum infections: Where do we stand? Mol. Plant Microbe Interact. 2018, 31, 889–898. [Google Scholar] [CrossRef] [PubMed]
- Rohila, J.S.; Yang, Y. Rice mitogen-activated protein kinase gene family and its role in biotic and abiotic stress response. J. Integr. Plant Biol. 2007, 49, 751–759. [Google Scholar] [CrossRef]
- Hamel, L.P.; Nicole, M.C.; Sritubtim, S.; Morency, M.J.; Ellis, M.; Ehlting, J.; Beaudoin, N.; Barbazuk, B.; Klessig, D.; Lee, J.; et al. Ancient signals: Comparative genomics of plant MAPK and MAPKK gene families. Trends Plant Sci. 2006, 11, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Rao, K.P.; Richa, T.; Kumar, K.; Raghuram, B.; Sinha, A.K. In silico analysis reveals 75 members of mitogen-activated protein kinase kinase kinase gene family in rice. DNA Res. 2010, 17, 139–153. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Li, J.; Hu, L.; Zhang, T.; Zhang, G.; Lou, Y. OsMPK3 positively regulates the JA signaling pathway and plant resistance to a chewing herbivore in rice. Plant Cell Rep. 2013, 32, 1075–1084. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.-Y.; You, M.K.; Jeung, J.U.; Shin, J.S. OsMPK3 is a TEY-type rice MAPK in Group C and phosphorylates OsbHLH65, a transcription factor binding to the E-box element. Plant Cell Rep. 2014, 33, 1343–1353. [Google Scholar] [CrossRef]
- Kandoth, P.K.; Ranf, S.; Pancholi, S.S.; Jayanty, S.; Walla, M.D.; Miller, W.; Howe, G.A.; Lincoln, D.E.; Stratmann, J.W. Tomato MAPKs LeMPK1, LeMPK2, and LeMPK3 function in the systemin-mediated defense response against herbivorous insects. Proc. Natl. Acad. Sci. USA 2007, 104, 12205. [Google Scholar] [CrossRef]
- Fu, Z.Q.; Yan, S.; Saleh, A.; Wang, W.; Ruble, J.; Oka, N.; Mohan, R.; Spoel, S.H.; Tada, Y.; Zheng, N.; et al. NPR3 and NPR4 are receptors for the immune signal salicylic acid in plants. Nature 2012, 486, 228–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Li, J.; Ju, H.; Liu, X.; Erb, M.; Wang, X.; Lou, Y. Contrasting effects of ethylene biosynthesis on induced plant resistance against a chewing and a piercing-sucking herbivore in rice. Mol. Plant 2014, 7, 1670–1682. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Pan, C.; Wang, Y.; Ye, L.; Wu, J.; Chen, L.; Zou, T.; Lu, G. Genome-wide identification of MAPK, MAPKK, and MAPKKK gene families and transcriptional profiling analysis during development and stress response in cucumber. BMC Genom. 2015, 16, 386. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhou, J.; Peng, X.; Xu, H.; Liu, C.; Du, B.; Yuan, B.; Zhu, L.; He, G. The Bphi008a gene interacts with the ethylene pathway and transcriptionally regulates MAPK genes in the response of rice to brown planthopper feeding. Plant Physiol. 2011, 156, 856–872. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Hettenhausen, C.; Meldau, S.; Baldwin, I.T. Herbivory rapidly activates MAPK signaling in attacked and unattacked leaf regions but not between leaves of Nicotiana attenuata. Plant Cell 2007, 19, 1096–1122. [Google Scholar] [CrossRef]
- Hu, L.; Ye, M.; Li, R.; Zhang, T.; Zhou, G.; Wang, Q.; Lu, J.; Lou, Y. The rice transcription factor WRKY53 suppresses herbivore-induced defenses by acting as a negative feedback modulator of mitogen-activated protein kinase activity. Plant Physiol. 2015, 169, 2907–2921. [Google Scholar] [CrossRef]
- Singh, I.K.; Kumar, S.; Singh, S.; Singh, A. Expression profiling of mitogen-activated protein kinase genes from chickpea (Cicer arietinum L.) in response to Helicoverpa armigera, wounding and signaling compounds. J. Asia Pac. Entomol. 2017, 20, 942–948. [Google Scholar] [CrossRef]
- Reyna, N.S.; Yang, Y. Molecular analysis of the rice map kinase gene family in relation to Magnaporthe grisea infection. Mol. Plant Microbe Interact. 2006, 19, 530–540. [Google Scholar] [CrossRef]
- Damien, L.; Nguyen, P.T.; Ayako, N.; Kenji, U.; Tsutomu, K.; Ko, S. A sphingolipid elicitor-inducible mitogen-activated protein kinase is regulated by the small GTPase OsRac1 and heterotrimeric G-protein in rice. Plant Physiol. 2005, 138, 1644–1652. [Google Scholar] [CrossRef]
- Liu, X.; Li, J.; Xu, L.; Wang, Q.; Lou, Y. Expressing OsMPK4 impairs plant growth but enhances the resistance of rice to the striped stem borer Chilo suppressalis. Int. J. Mol. Sci. 2018, 1182. [Google Scholar] [CrossRef]
- Jalmi, S.K.; Sinha, A.K. Functional involvement of a mitogen activated protein kinase module, OsMKK3-OsMPK7-OsWRK30 in mediating resistance against Xanthomonas oryzae in rice. Sci. Rep. 2016, 6, 37974. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, G.K.; Tamogami, S.; Iwahashi, H.; Agrawal, V.P.; Rakwal, R. Transient regulation of jasmonic acid-inducible rice MAP kinase gene (OsBWMK1) by diverse biotic and abiotic stresses. Plant Physiol. Biochem. 2003, 41, 355–361. [Google Scholar] [CrossRef]
- Ji, R.; Ye, W.; Chen, H.; Zeng, J.; Li, H.; Yu, H.; Li, J.; Lou, Y. A salivary endo-β-1,4-glucanase acts as an effector that enables the brown planthopper to feed on rice. Plant Physiol. 2017, 173, 1920–1932. [Google Scholar] [CrossRef] [PubMed]
- Ye, W.; Yu, H.; Jian, Y.; Zeng, J.; Ji, R.; Chen, H.; Lou, Y. A salivary EF-hand calcium-binding protein of the brown planthopper Nilaparvata lugens functions as an effector for defense responses in rice. Sci. Rep. 2017, 7, 40498. [Google Scholar] [CrossRef] [PubMed]
- Rudd, J.J.; Keon, J.; Hammond-Kosack, K.E. The wheat mitogen-activated protein kinases TaMPK3 and TaMPK6 are differentially regulated at multiple levels during compatible disease interactions with Mycosphaerella graminicola. Plant Physiol. 2008, 147, 802–815. [Google Scholar] [CrossRef] [PubMed]
- Stuart, J. Insect effectors and gene-for-gene interactions with host plants. Curr. Opin. Insect Sci. 2015, 9, 56–61. [Google Scholar] [CrossRef]
- Dempsey, D.M.A.; Vlot, A.C.; Wildermuth, M.C.; Klessig, D.F. Salicylic acid biosynthesis and metabolism. Arabidopsis Book Am. Soc. Plant Biol. 2011, 9, e0156. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.Z.; Xue, Y.J.; Zhou, J.J.; Zhang, B.; Chang, H.; Takano, M. Phytochromes regulate SA and JA signaling pathways in rice and are required for developmentally controlled resistance to Magnaporthe grisea. Mol. Plant 2011, 4, 688–696. [Google Scholar] [CrossRef]
- Li, X.; Yang, D.L.; Sun, L.; Li, Q.; Mao, B.; He, Z. The systemic acquired resistance regulator OsNPR1 attenuates growth by repressing auxin signaling through promoting IAA-amido synthase expression. Plant Physiol. 2016, 172, 546. [Google Scholar] [CrossRef]
- Lu, J.; Ju, H.; Zhou, G.; Zhu, C.; Erb, M.; Wang, X.; Wang, P.; Lou, Y. An EAR-motif-containing ERF transcription factor affects herbivore-induced signaling, defense and resistance in rice. Plant J. 2011, 68, 583–596. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Li, W.; Cao, J.; Meng, F.; Yu, Y.; Huang, J.; Jiang, L.; Liu, M.; Zhang, Z.; Chen, X.; et al. Activation of ethylene signaling pathways enhances disease resistance by regulating ROS and phytoalexin production in rice. Plant J. 2016, 89, 338–353. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Zhang, W.; Liu, B.; Hu, J.; Wei, Z.; Shi, Z.; He, R.; Zhu, L.; Chen, R.; Han, B.; et al. Identification and characterization of Bph14, a gene conferring resistance to brown planthopper in rice. Proc. Natl. Acad. Sci. USA 2009, 106, 22163–22168. [Google Scholar] [CrossRef] [PubMed]
- Zhan, L.P.; Peng, D.L.; Wang, X.L.; Kong, L.A.; Peng, H.; Liu, S.M.; Liu, Y.; Huang, W.K. Priming effect of root-applied silicon on the enhancement of induced resistance to the root-knot nematode Meloidogyne graminicola in rice. BMC Plant Biol. 2018, 18, 50. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, X.; Yuan, H.; Chen, R.; Zhu, L.; He, R.; He, G. Responses of two contrasting genotypes of rice to brown planthopper. Mol. Plant Microbe Interact. 2007, 21, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Qi, J.; Ren, N.; Cheng, J.; Erb, M.; Mao, B.; Lou, Y. Silencing OsHI-LOX makes rice more susceptible to chewing herbivores, but enhances resistance to a phloem feeder. Plant J. 2009, 60, 638–648. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Luo, C.; Zhou, Z.; Wang, R.; Ling, F.; Xiao, L.; Lin, Y.; Chen, H. Gene expression and plant hormone levels in two contrasting rice genotypes responding to brown planthopper infestation. BMC Plant Biol. 2017, 17, 57. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Malook, S.; Shen, G.; Gao, L.; Zhang, C.; Li, J.; Zhang, J.; Wang, L.; Wu, J. Current understanding of maize and rice defense against insect herbivores. Plant Divers. 2018, 40, 189–195. [Google Scholar] [CrossRef]
- Liu, C.; Hao, F.; Hu, J.; Zhang, W.; Wan, L.; Zhu, L.; Tang, H.; He, G. Revealing different systems responses to brown planthopper infestation for pest susceptible and resistant rice plants with the combined metabonomic and gene-expression analysis. J. Proteome Res. 2010, 9, 6774–6785. [Google Scholar] [CrossRef]
- Luo, X.; Xu, N.; Huang, J.; Gao, F.; Zou, H.; Boudsocq, M.; Coaker, G.; Liu, J. A lectin receptor-like kinase mediates pattern-triggered salicylic acid signaling. Plant Physiol. 2017, 174, 2501–2514. [Google Scholar] [CrossRef]
- Woo, J.Y.; Jeong, K.J.; Kim, Y.J.; Paek, K.-H. CaLecRK-S.5, a pepper L-type lectin receptor kinase gene, confers broad-spectrum resistance by activating priming. J. Exp. Bot. 2016, 67, 5725–5741. [Google Scholar] [CrossRef]
- Zhang, X.; Mi, X.; Chen, C.; Wang, H.; Guo, W. Identification on mitogen-activated protein kinase signaling cascades by integrating protein interaction with transcriptional profiling analysis in cotton. Sci. Rep. 2018, 8, 8178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, P.; Mohanta, T.K.; Sinha, A.K. Unraveling the intricate nexus of molecular mechanisms governing rice root development: OsMPK3/6 and auxin-cytokinin interplay. PLoS ONE 2015, 10, e0123620. [Google Scholar] [CrossRef] [PubMed]
- Alessi, D.R.; Cuenda, A.; Cohen, P.; Dudley, D.T.; Saltiel, A.R. PD 098059 is a specific inhibitor of the activation of mitogen-activated protein kinase kinase in vitro and in vivo. J. Biol. Chem. 1995, 270, 27489–27494. [Google Scholar] [CrossRef] [PubMed]
- Bjornson, M.; Benn, G.; Song, X.; Comai, L.; Franz, A.K.; Dandekar, A.M.; Drakakaki, G.; Dehesh, K. Distinct roles for Mitogen-activated protein kinase signaling and Calmodulin-binding transcriptional activator3 in regulating the peak time and amplitude of the plant general stress response. Plant Physiol. 2014, 166, 988–996. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.; Savatin, D.; Sicilia, F.; Gramegna, G.; Cervone, F.; De Lorenzo, G. Oligogalacturonides: Plant damage-associated molecular patterns and regulators of growth and development. Front. Plant Sci. 2013, 4, 49. [Google Scholar] [CrossRef] [PubMed]
- Dang, T.T.; Shimatani, Z.; Kawano, Y.; Terada, R.; Shimamoto, K. Gene editing a constitutively active OsRac1 by homologous recombination-based gene targeting induces immune responses in rice. Plant Cell Physiol. 2013, 54, 2058–2070. [Google Scholar] [CrossRef]
- Jannoey, P.; Channei, D.; Kotcharerk, J.; Pongprasert, W.; Nomura, M. Expression analysis of genes related to rice resistance against brown planthopper, Nilaparvata lugens. Rice Sci. 2017, 24, 163–172. [Google Scholar] [CrossRef]
- Mentlak, T.A.; Kombrink, A.; Shinya, T.; Ryder, L.S.; Otomo, I.; Saitoh, H.; Terauchi, R.; Nishizawa, Y.; Shibuya, N.; Thomma, B.P.H.J.; Talbot, N.J. Effector-mediated suppression of chitin-triggered immunity by Magnaporthe oryzae is necessary for rice blast disease. Plant Cell 2012, 24, 322–335. [Google Scholar] [CrossRef]
- Nahar, K.; Kyndt, T.; De Vleesschauwer, D.; Höfte, M.; Gheysen, G. The jasmonate pathway is a key player in systemically induced defense against root knot nematodes in rice. Plant Physiol. 2011, 157, 305–316. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Tang, Q.Y.; Zhang, C.X. Data Processing System (DPS) software with experimental design, statistical analysis and data mining developed for use in entomological research. Insect Sci. 2012, 20, 254–260. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Biotic Stress Type |
|---|---|
| OsMPK1 | Magnaporthe grisea [33] |
| OsMPK2 | M. grisea [32] |
| OsMPK3 | M. grisea and BPH [23] |
| OsMPK4 | M. grisea [32] and Chilo suppressalis infestation [34] |
| OsMPK5 | M. grisea [32], C. suppressalis [22], and BPH infestation [28] |
| OsMPK6 | C. suppressalis infestation [30] |
| OsMPK7 | M. grisea [32] and Xanthomonas oryzae [35] |
| OsMPK8 | M. grisea [32] |
| OsMPK12 | Fungal elicitor chitosan [36], M. grisea [32], and BPH infestation [28] |
| OsMPK13 | M. grisea [32] and BPH infestation [28] |
| OsMPK15 | M. grisea [32] |
| OsMPK17 | M. grisea [32] and BPH infestation [28] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nanda, S.; Wan, P.-J.; Yuan, S.-Y.; Lai, F.-X.; Wang, W.-X.; Fu, Q. Differential Responses of OsMPKs in IR56 Rice to Two BPH Populations of Different Virulence Levels. Int. J. Mol. Sci. 2018, 19, 4030. https://doi.org/10.3390/ijms19124030
Nanda S, Wan P-J, Yuan S-Y, Lai F-X, Wang W-X, Fu Q. Differential Responses of OsMPKs in IR56 Rice to Two BPH Populations of Different Virulence Levels. International Journal of Molecular Sciences. 2018; 19(12):4030. https://doi.org/10.3390/ijms19124030
Chicago/Turabian StyleNanda, Satyabrata, Pin-Jun Wan, San-Yue Yuan, Feng-Xiang Lai, Wei-Xia Wang, and Qiang Fu. 2018. "Differential Responses of OsMPKs in IR56 Rice to Two BPH Populations of Different Virulence Levels" International Journal of Molecular Sciences 19, no. 12: 4030. https://doi.org/10.3390/ijms19124030
APA StyleNanda, S., Wan, P. -J., Yuan, S. -Y., Lai, F. -X., Wang, W. -X., & Fu, Q. (2018). Differential Responses of OsMPKs in IR56 Rice to Two BPH Populations of Different Virulence Levels. International Journal of Molecular Sciences, 19(12), 4030. https://doi.org/10.3390/ijms19124030