Altered Circadian Timing System-Mediated Non-Dipping Pattern of Blood Pressure and Associated Cardiovascular Disorders in Metabolic and Kidney Diseases


Abstract
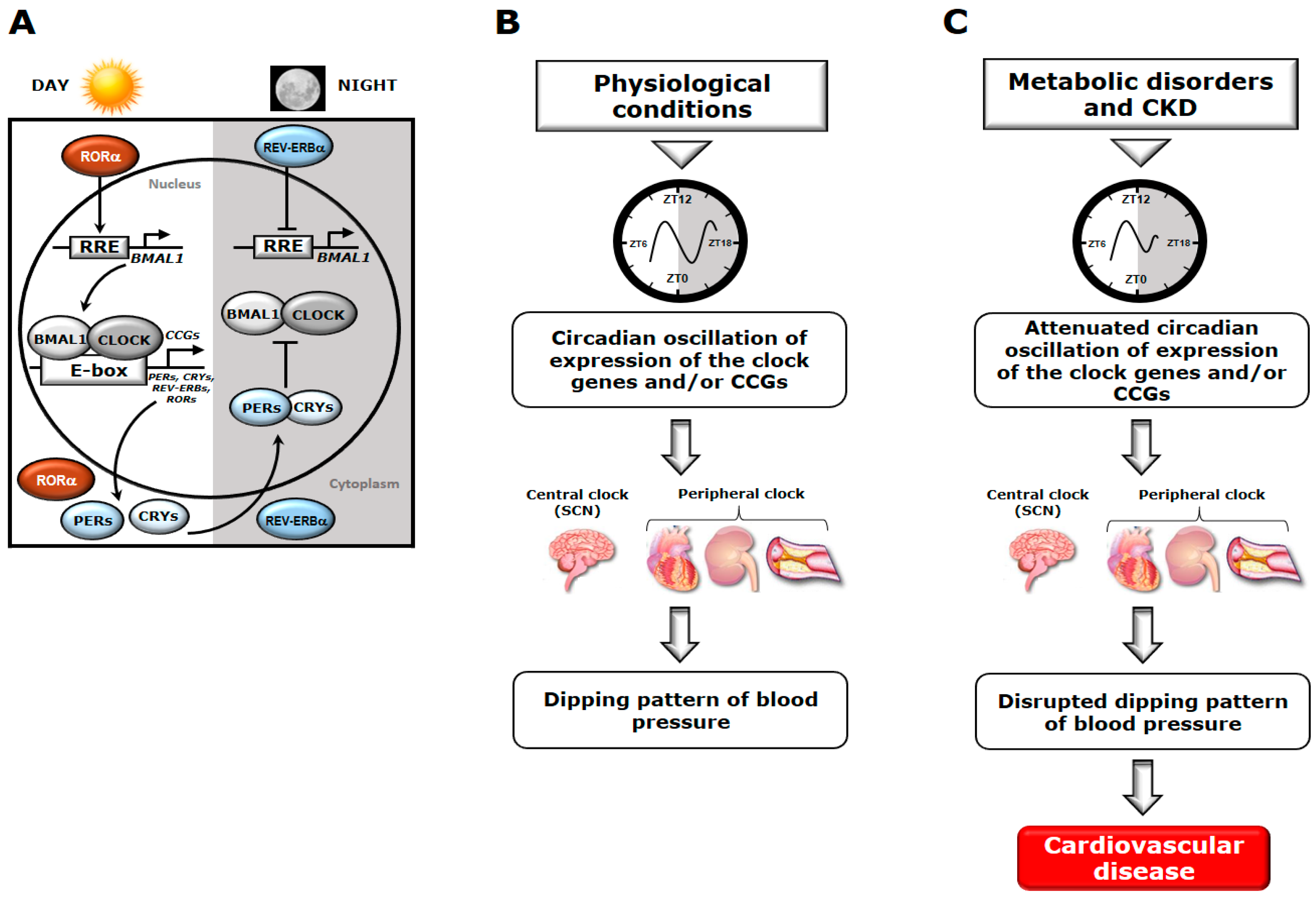
:1. Introduction
2. Circadian Timing System and BP Regulation
3. The Circadian Timing System and Diurnal Variation of BP in Metabolic Dysfunction
3.1. Glucose Homeostasis and Insulin Function
3.2. Cholesterol and Triglycerides
3.3. Endothelial Function and Vascular Tone
3.4. Autonomic Nervous Function
3.5. Sleep Disturbances
3.6. Genetic Involvement
4. The Circadian Timing System and Diurnal Variation of BP in CKD
4.1. Renal Function
4.2. Endothelin and Renin Angiotensin System
4.3. Water and Electrolyte Homeostasis
4.4. Renal Fibrosis
4.5. Melatonin
5. Altered Circadian Timing System and Disrupted Dipping Pattern of BP in the Development of CV Disorders
6. Circadian Timing System-Based Therapies for the Non-Dipping Pattern of Hypertension in Metabolic and Kidney Diseases
6.1. Established Therapeutics Modulating the Dipping Pattern of BP Based on the Time of Administration
6.2. Therapeutics Modulating the Circadian Timing System in Metabolic Disorders and Kidney Disease
6.2.1. Melatonin
6.2.2. Novel Compounds Targeting Molecular Components of the Circadian Timing System
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- White, W.B. Ambulatory blood pressure monitoring: Dippers compared with non-dippers. Blood Press. Monit. 2000, 5 (Suppl. 1), S17–S23. [Google Scholar] [CrossRef] [PubMed]
- Karaagac, K.; Tenekecioglu, E.; Yontar, O.C.; Kuzeytemiz, M.; Vatansever, F.; Tutuncu, A.; Ozluk, O.A.; Yilmaz, M.; Demir, M. Effect of non-dipper and dipper blood pressure patterns on Tp-Te interval and Tp-Te/QT ratio in patients with metabolic syndrome. Int. J. Clin. Exp. Med. 2014, 7, 1397–1403. [Google Scholar] [PubMed]
- Richards, J.; Diaz, A.N.; Gumz, M.L. Clock genes in hypertension: Novel insights from rodent models. Blood Press. Monit. 2014, 19, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, I.; Yoshida, Y.; Minamino, T. A role for circadian clock in metabolic disease. Hypertens. Res. 2016, 39, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Takeda, N.; Maemura, K. Circadian clock and vascular disease. Hypertens. Res. 2010, 3368, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Rudic, R.D.; Fulton, D.J. Pressed for time: The circadian clock and hypertension. J. Appl. Physiol. 2009, 107, 1328–1338. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, R. Regulation of circadian blood pressure—From mice to astronauts. Curr. Opin. Nephrol. Hypertens. 2010, 19, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.; Gumz, M.L. Mechanism of the circadian clock in physiology. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 302, R1053–R1064. [Google Scholar] [CrossRef] [PubMed]
- Gekakis, N.; Staknis, D.; Nguyen, H.B.; Davis, F.C.; Wilsbacher, L.D.; King, D.P.; Takahashi, J.S.; Weitz, C.J. Role of the CLOCK protein in the mammalian circadian mechanism. Science 1998, 280, 1564–1569. [Google Scholar] [CrossRef] [PubMed]
- Kume, K.; Zylka, M.J.; Sriram, S.; Shearman, L.P.; Weaver, D.R.; Jin, X.; Maywood, E.S.; Hastings, M.H.; Reppert, S.M. mCRY1 and mCRY2 are essential components of the negative limb of the circadian clock feedback loop. Cell 1999, 98, 193–205. [Google Scholar] [CrossRef]
- Preitner, N.; Damiola, F.; Lopez-Molina, L.; Zakany, J.; Duboule, D.; Albrecht, U.; Schibler, U. The orphan nuclear receptor REV-ERBalpha controls circadian transcription within the positive limb of the mammalian circadian oscillator. Cell 2002, 110, 251–260. [Google Scholar] [CrossRef]
- Sato, T.K.; Panda, S.; Miraglia, L.J.; Reyes, T.M.; Rudic, R.D.; McNamara, P.; Naik, K.A.; FitzGerald, G.A.; Kay, S.A.; Hogenesch, J.B. A functional genomics strategy reveals Rora as a component of the mammalian circadian clock. Neuron 2004, 43, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Witte, K.; Schnecko, A.; Buijs, R.M.; van der Vliet, J.; Scalbert, E.; Delagrange, P.; Guardiola-Lemaître, B.; Lemmer, B. Effects of SCN lesions on circadian blood pressure rhythm in normotensive and transgenic hypertensive rats. Chronobiol. Int. 1998, 15, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Storch, K.-F.; Lipan, O.; Leykin, I.; Viswanathan, N.; Davis, F.C.; Wong, W.H.; Weitz, C.J. Extensive and divergent circadian gene expression in liver and heart. Nature 2002, 417, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Perk, G.; Mekler, J.; Ben Ishay, D.; Bursztyn, M. Non-dipping in diabetic patients: Insights from the siesta. J. Hum. Hypertens. 2002, 16, 435–438. [Google Scholar] [CrossRef] [PubMed]
- Poantă, L.; Cerghizan, A.; Pop, D. Blood pressure pattern and heart rate variability in normotensive patients with type 2 diabetes mellitus. Rom. J. Intern. Med. 2010, 48, 321–327. [Google Scholar] [PubMed]
- Su, W.; Guo, Z.; Randall, D.C.; Cassis, L.; Brown, D.R.; Gong, M.C. Hypertension and disrupted blood pressure circadian rhythm in Type 2 diabetic db/db mice. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H1634–H1641. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Xie, Z.; Guo, Z.; Duncan, M.J.; Lutshumba, J.; Gong, M.C. Altered clock gene expression and vascular smooth muscle diurnal contractile variations in type 2 diabetic db/db mice. Am. J. Physiol. Heart Circ. Physiol. 2012, 302, H621–H633. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, M.C.; Yang, S.C.; Tseng, H.L.; Hwang, L.L.; Chen, C.T.; Shieh, K.R. Abnormal expressions of circadian-clock and circadian clock-controlled genes in the livers and kidneys of long-term, high-fat-diet-treated mice. Int. J. Obes. 2010, 34, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Kudo, T.; Akiyama, M.; Kuriyama, K.; Sudo, M.; Moriya, T.; Shibata, S. Night-time restricted feeding normalises clock genes and Pai-1 gene expression in the db/db mouse liver. Diabetologia 2004, 47, 1425–1436. [Google Scholar] [CrossRef] [PubMed]
- Oishi, K.; Ohkura, N.; Amagai, N.; Ishida, N. Involvement of circadian clock gene Clock in diabetes-induced circadian augmentation of plasminogen activator inhibitor-1 (PAI-1) expression in the mouse heart. FEBS Lett. 2005, 579, 3555–3559. [Google Scholar] [CrossRef] [PubMed]
- Fonken, L.K.; Aubrecht, T.G.; Meléndez-Fernández, O.H.; Weil, Z.M.; Nelson, R.J. Dim Light at Night Disrupts Molecular Circadian Rhythms and Increases Body Weight. J. Biol. Rhythms 2013, 28, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Woon, P.Y.; Kaisaki, P.J.; Bragança, J.; Rè Se Bihoreau, M.T.; Levy, J.C.; Farrall, M.; Gauguier, D.; Takahashi, J.S. Aryl hydrocarbon receptor nuclear translocator-like (BMAL1) is associated with susceptibility to hypertension and type 2 diabetes. Proc. Natl. Acad. Sci. USA 2007, 104, 14412–14417. [Google Scholar] [CrossRef] [PubMed]
- Corella, D.; Asensio, E.M.; Coltell, O.; Sorlí, J.V.; Estruch, R.; Martínez González, M.Á.; Salas-Salvadó, J.; Castañer, O.; Arós, F.; Lapetra, J.; et al. CLOCK gene variation is associated with incidence of type-2 diabetes and cardiovascular diseases in type-2 diabetic subjects: Dietary modulation in the PREDIMED randomized trial. Cardiovasc. Diabetol. 2016, 15, 4. [Google Scholar] [CrossRef] [PubMed]
- Sookoian, S.; Gianotti, T.F.; Burgueño, A.; Pirola, C.J.; Pirola, C.J. Gene-gene interaction between serotonin transporter (SLC6A4) and clock modulates the risk of metabolic syndrome in rotating shiftworkers. Chronobiol. Int. 2010, 27, 1202–1218. [Google Scholar] [CrossRef] [PubMed]
- Young, M.E.; Wilson, C.R.; Razeghi, P.; Guthrie, P.H.; Taegtmeyer, H. Alterations of the Circadian Clock in the Heart by Streptozotocin-induced Diabetes. J. Mol. Cell. Cardiol. 2002, 34, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Pistrosch, F.; Reissmann, E.; Wildbrett, J.; Koehler, C.; Hanefeld, M. Relationship between Diurnal Blood Pressure Variation and Diurnal Blood Glucose Levels in Type 2 Diabetic Patients. Am. J. Hypertens. 2007, 20, 541–545. [Google Scholar] [CrossRef] [PubMed]
- Saini, C.; Petrenko, V.; Pulimeno, P.; Giovannoni, L.; Berney, T.; Hebrok, M.; Howald, C.; Dermitzakis, E.T.; Dibner, C. A functional circadian clock is required for proper insulin secretion by human pancreatic islet cells. Diabetes Obes. MeTable 2016, 18, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Marcheva, B.; Ramsey, K.M.; Buhr, E.D.; Kobayashi, Y.; Su, H.; Ko, C.H.; Ivanova, G.; Omura, C.; Mo, S.; Vitaterna, M.H.; et al. Disruption of the clock components CLOCK and BMAL1 leads to hypoinsulinaemia and diabetes. Nature 2010, 466, 627–631. [Google Scholar] [CrossRef] [PubMed]
- Anan, F.; Takahashi, N.; Ooie, T.; Yufu, K.; Saikawa, T.; Yoshimatsu, H. Role of insulin resistance in nondipper essential hypertensive patients. Hypertens. Res. 2003, 26, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Laakso, M.; Kuusisto, J. Insulin resistance and hyperglycaemia in cardiovascular disease development. Nat. Rev. Endocrinol. 2014, 10, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Rudic, R.D.; McNamara, P.; Curtis, A.-M.; Boston, R.C.; Panda, S.; Hogenesch, J.B.; Fitzgerald, G.A. BMAL1 and CLOCK, two essential components of the circadian clock, are involved in glucose homeostasis. PLoS Biol. 2004, 2, e377. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Ansari, T.S.; McGuinness, O.P.; Wasserman, D.H.; Johnson, C.H. Circadian disruption leads to insulin resistance and obesity. Curr. Biol. 2013, 23, 372–381. [Google Scholar] [CrossRef] [PubMed]
- Mergeani, A.C.; Antochi, F.; Rusu, O.; Ciobotaru, A.; Coclitu, C.; Bajenaru, O.A. Correlations of cognitive impairment with circadian blood pressure pattern and intima-media thickness in hypertensive patients. Maedica (Buchar) 2015, 10, 325–330. [Google Scholar] [PubMed]
- Akashi, M.; Matsumura, R.; Matsuo, T.; Kubo, Y.; Komoda, H.; Node, K. Hypercholesterolemia causes circadian dysfunction: A potential risk factor for cardiovascular disease. EBioMedicine 2017, 20, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Vasunta, R.-L.; Kesäniemi, Y.A.; Ylitalo, A.; Ukkola, O. Nondipping pattern and carotid atherosclerosis in a middle-aged population: OPERA Study. Am. J. Hypertens. 2012, 25, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Pourcet, B.; Zecchin, M.; Vanhoutte, J.; Delhaye, S.; Rommens, C.; Woldt, E.; Eeckhoute, J.; Staels, B.; Duez, H. Role of the nuclear receptor Rev-erb-alpha in the development of atherosclerosis. Atherosclerosis 2015, 241, e6. [Google Scholar] [CrossRef]
- Shostak, A.; Meyer-Kovac, J.; Oster, H. Circadian regulation of lipid mobilization in white adipose tissues. Diabetes 2013, 62, 2195–2203. [Google Scholar] [CrossRef] [PubMed]
- Bray, M.S.; Young, M.E. Circadian rhythms in the development of obesity: Potential role for the circadian clock within the adipocyte. Obes. Rev. 2007, 8, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Chudy, P.; Kotulicova, D.; Stasko, J.; Kubisz, P. The relationship among TAFI, t-PA, PAI-1 and F1 + 2 in type 2 diabetic patients with normoalbuminuria and microalbuminuria. Blood Coagul. Fibrinolysis 2011, 22, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, J.; Hoshide, S.; Eguchi, K.; Ishikawa, S.; Pickering, T.G.; Shimada, K.; Kario, K. Increased low-grade inflammation and plasminogen-activator inhibitor-1 level in nondippers with sleep apnea syndrome. J. Hypertens. 2008, 26, 1181–1187. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Su, W.; Liu, S.; Zhao, G.; Esser, K.; Schroder, E.A.; Lefta, M.; Stauss, H.M.; Guo, Z.; Gong, M.C. Smooth-muscle BMAL1 participates in blood pressure circadian rhythm regulation. J. Clin. Investig. 2015, 125, 324–336. [Google Scholar] [CrossRef] [PubMed]
- Anea, C.B.; Zhang, M.; Stepp, D.W.; Simkins, G.B.; Reed, G.; Fulton, D.J.; Rudic, R.D. Vascular disease in mice with a dysfunctional circadian clock. Circulation 2009, 119, 1510–1517. [Google Scholar] [CrossRef] [PubMed]
- Somanath, P.R.; Podrez, E.A.; Chen, J.; Ma, Y.; Marchant, K.; Antoch, M.; Byzova, T.V. Deficiency in core circadian protein Bmal1 is associated with a prothrombotic and vascular phenotype. J. Cell. Physiol. 2011, 226, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Anea, C.B.; Zhang, M.; Chen, F.; Ali, M.I.; Hart, C.M.M.; Stepp, D.W.; Kovalenkov, Y.O.; Merloiu, A.-M.; Pati, P.; Fulton, D.; et al. Circadian clock control of Nox4 and reactive oxygen species in the vasculature. PLoS ONE 2013, 8, e78626. [Google Scholar] [CrossRef] [PubMed]
- Anea, C.B.; Cheng, B.; Sharma, S.; Kumar, S.; Caldwell, R.W.; Yao, L.; Ali, M.I.; Merloiu, A.M.; Stepp, D.W.; Black, S.M.; et al. Increased superoxide and endothelial NO synthase uncoupling in blood vessels of Bmal1-Knockout Mice. Circ. Res. 2012, 111, 1157–1165. [Google Scholar] [CrossRef] [PubMed]
- Anea, C.B.; Ali, M.I.; Osmond, J.M.; Sullivan, J.C.; Stepp, D.W.; Merloiu, A.M.; Rudic, R.D. Matrix metalloproteinase 2 and 9 dysfunction underlie vascular stiffness in circadian clock mutant mice. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2535–2543. [Google Scholar] [CrossRef] [PubMed]
- Duan, S.Z.; Usher, M.G.; Mortensen, R.M. Peroxisome proliferator-activated receptor—Mediated effects in the vasculature. Circ. Res. 2008, 102, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Yang, G.; Jia, Z.; Zhang, H.; Aoyagi, T.; Soodvilai, S.; Symons, J.D.; Schnermann, J.B.; Gonzalez, F.J.; Litwin, S.E.; et al. Vascular PPARγ Controls Circadian Variation in Blood Pressure and Heart Rate through Bmal1. Cell Metab. 2008, 8, 482–491. [Google Scholar] [CrossRef] [PubMed]
- Viswambharan, H.; Carvas, J.M.; Antic, V.; Marecic, A.; Jud, C.; Zaugg, C.E.; Ming, X.-F.; Montani, J.-P.; Albrecht, U.; Yang, Z. Mutation of the Circadian Clock Gene Per2 Alters Vascular Endothelial Function. Circulation 2007, 115, 2188–2195. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-Y.; Wen, M.-S.; Wang, H.-W.; Hsieh, I.-C.; Li, Y.; Liu, P.-Y.; Lin, F.-C.; Liao, J.K. Increased Vascular Senescence and Impaired Endothelial Progenitor Cell Function Mediated by Mutation of Circadian Gene Per2. Circulation 2008, 118, 2166–2173. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Tang, X.; Zhu, Z.; Liao, X.; Zhao, R.; Fu, W.; Chen, B.; Jiang, J.; Qian, R.; Guo, D. The rhythmic expression of clock genes attenuated in human plaque-derived vascular smooth muscle cells. Lipids Health Dis. 2014, 13, 14. [Google Scholar] [CrossRef] [PubMed]
- Warren, W.S.; Champney, T.H.; Cassone, V.M. The suprachiasmatic nucleus controls the circadian rhythm of heart rate via the sympathetic nervous system. Physiol. Behav. 1994, 55, 1091–1099. [Google Scholar] [CrossRef]
- Maury, E.; Ramsey, K.M.; Bass, J. Circadian Rhythms and Metabolic Syndrome: From Experimental Genetics to Human Disease. Circ. Res. 2010, 106, 447–462. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.J.; Purvis, T.E.; Hu, K.; Scheer, F.A. Circadian misalignment increases cardiovascular disease risk factors in humans. Proc. Natl. Acad. Sci. USA 2016, 113, E1402–E1411. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Fujisawa, Y.; Nakano, D.; Hitomi, H.; Nishiyama, A. Effect of a selective SGLT2 inhibitor, luseogliflozin, on circadian rhythm of sympathetic nervous function and locomotor activities in metabolic syndrome rats. Clin. Exp. Pharmacol. Physiol. 2017, 44, 522–525. [Google Scholar] [CrossRef] [PubMed]
- Dabrowska, E.; Lewandowski, J.; Jedrusik, P.; Symonides, B.; Wocial, B.; Lapinski, M.; Gaciong, Z. Catecholamine excretion and circadian blood pressure profile in patients with pheochromocytoma. Ann. N. Y. Acad. Sci. 2006, 1073, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Curtis, A.M.; Cheng, Y.; Kapoor, S.; Reilly, D.; Price, T.S.; Fitzgerald, G.A. Circadian variation of blood pressure and the vascular response to asynchronous stress. Proc. Natl. Acad. Sci. USA 2007, 104, 3450–3455. [Google Scholar] [CrossRef] [PubMed]
- Terazono, H.; Mutoh, T.; Yamaguchi, S.; Kobayashi, M.; Akiyama, M.; Udo, R.; Ohdo, S.; Okamura, H.; Shibata, S. Adrenergic regulation of clock gene expression in mouse liver. Proc. Natl. Acad. Sci. USA 2003, 100, 6795–6800. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhang, L.; Zhang, Y.; Zhang, B.; He, Y.; Xie, S.; Li, M.; Miao, X.; Chan, E.Y.Y.; Tang, J.L.; et al. Meta-analysis on night shift work and risk of metabolic syndrome. Obes. Rev. 2014, 15, 709–720. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Nam, J.; Lee, J.K.; Oh, S.S.; Kang, H.T.; Koh, S.B. Association between night work and cardiovascular diseases: Analysis of the 3rd Korean working conditions survey. Ann. Occup. Environ. Med. 2015, 27, 15. [Google Scholar] [CrossRef] [PubMed]
- Gale, J.E.; Cox, H.I.; Qian, J.; Block, G.D.; Colwell, C.S.; Matveyenko, A.V. Disruption of Circadian Rhythms Accelerates Development of Diabetes through Pancreatic Beta-Cell Loss and Dysfunction. J. Biol. Rhythms 2011, 26, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Manning, A.K.; Hivert, M.F.; Scott, R.A.; Grimsby, J.L.; Bouatia Naji, N.; Chen, H.; Rybin, D. A genome-wide approach accounting for body mass index identifies genetic variants influencing fasting glycemic traits and insulin resistance. Nat. Genet. 2012, 44, 659–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leu, H.B.; Chung, C.M.; Lin, S.J.; Chiang, K.M.; Yang, H.C.; Ho, H.Y.; Ting, C.T.; Lin, T.H.; Sheu, S.H.; Tsai, W.C.; et al. Association of circadian genes with diurnal blood pressure changes and non-dipper essential hypertension: A genetic association with young-onset hypertension. Hypertens. Res. 2015, 38, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Valladares, M.; Obregon, A.M.; Chaput, J.-P. Association between genetic variants of the clock gene and obesity and sleep duration. J. Physiol. Biochem. 2015, 71, 855–860. [Google Scholar] [CrossRef] [PubMed]
- Kelly, M.A.; Rees, S.D.; Hydrie, M.Z.I.; Shera, A.S.; Bellary, S.; O’Hare, J.P.; Kumar, S.; Taheri, S.; Basit, A.; Barnett, A.H.; et al. Circadian Gene Variants and Susceptibility to Type 2 Diabetes: A Pilot Study. PLoS ONE 2012, 7, e32670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupuis, J.; Langenberg, C.; Prokopenko, I.; Saxena, R.; Soranzo, N.; Jackson, A.U. New genetic loci implicated in fasting glucose homeostasis and their impact on type 2 diabetes risk. Nat. Genet. 2010, 42, 105–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dashti, H.S.; Smith, C.E.; Lee, Y.-C.; Parnell, L.D.; Lai, C.-Q.; Arnett, D.K.; Ordovás, J.M.; Garaulet, M. CRY1 circadian gene variant interacts with carbohydrate intake for insulin resistance in two independent populations: Mediterranean and North American. Chronobiol. Int. 2014, 31, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Hanratty, R.; Chonchol, M.; Havranek, E.P.; Powers, J.D.; Dickinson, L.M.; Ho, P.M.; Magid, D.J.; Steiner, J.F. Relationship between blood pressure and incident chronic kidney disease in hypertensive patients. Clin. J. Am. Soc. Nephrol. 2011, 6, 2605–2611. [Google Scholar] [CrossRef] [PubMed]
- Gabbai, F.B.; Rahman, M.; Hu, B.; Appel, L.J.; Charleston, J.; Contreras, G.; Faulkner, M.L.; Hiremath, L.; Jamerson, K.A.; Lea, J.P.; et al. Relationship between ambulatory BP and clinical outcomes in patients with hypertensive CKD. Clin. J. Am. Soc. Nephrol. 2012, 7, 1770–1776. [Google Scholar] [CrossRef] [PubMed]
- McMullan, C.J.; Hickson, D.A.; Taylor, H.A.; Forman, J.P. Prospective analysis of the association of ambulatory blood pressure characteristics with incident chronic kidney disease. J. Hypertens. 2015, 33, 1939–1946. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Chen, W.; Yuan, J.; Yang, Y.; Mei, Q.; Huang, L. Altered diurnal variation and localization of clock proteins in the remnant kidney of 5/6 nephrectomy rats. Nephrology 2013, 18, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, M.; Tamura, K.; Wakui, H.; Maeda, A.; Ohsawa, M.; Kanaoka, T.; Azushima, K.; Uneda, K.; Haku, S.; Tsurumi-Ikeya, Y.; et al. Upstream stimulatory factors 1 and 2 mediate the transcription of angiotensin II binding and inhibitory protein. J. Biol. Chem. 2013, 288, 19238–19249. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Yuan, J.; Zeng, X.; Peng, C.; Mei, Q.; Chen, W. Effects of chronotherapy of benazepril on the diurnal profile of RAAS and clock genes in the kidney of 5/6 nephrectomy rats. J. Huazhong Univ. Sci. Technol. Med. Sci. 2013, 33, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Hamamura, K.; Matsunaga, N.; Ikeda, E.; Kondo, H.; Ikeyama, H.; Tokushige, K.; Itcho, K.; Furuichi, Y.; Yoshida, Y.; Matsuda, M.; et al. Alterations of hepatic metabolism in chronic kidney disease via D-box-binding protein aggravate the renal dysfunction. J. Biol. Chem. 2016, 291, 4913–4927. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.D.; Yeh, J.K.; Peng, M.T.; Shie, S.S.; Lin, S.L.; Yang, C.H.; Chen, T.H.; Hung, K.C.; Wang, C.C.; Hsieh, I.C.; et al. Circadian CLOCK mediates activation of transforming growth factor-beta; signaling and renal fibrosis through cyclooxygenase 2. Am. J. Pathol. 2015, 185, 3152–3163. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.-Y.; Chang, F.-C.; Ng, H.-Y.; Kuo, C.-C.; Lee, Y.-T.; Lu, C.-Y.; Lee, C.-T. Disrupted circadian rhythm in rats with nephrectomy-induced chronic kidney disease. Life Sci. 2012, 91, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Goto, N.; Uchida, K.; Morozumi, K.; Ueki, T.; Matsuoka, S.; Katayama, A.; Haba, T.; Tominaga, Y.; Fukuda, M.; Nakao, A.; et al. Circadian blood pressure rhythm is disturbed by nephrectomy. Hypertens. Res. 2005, 28, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Dhaun, N.; Moorhouse, R.; MacIntyre, I.M.; Melville, V.; Oosthuyzen, W.; Kimmitt, R.A.; Brown, K.E.; Kennedy, E.D.; Goddard, J.; Webb, D.J. Diurnal variation in blood pressure and arterial stiffness in chronic kidney disease: The Role of Endothelin-1. Hypertension 2014, 64, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.; Welch, A.K.; Barilovits, S.J.; All, S.; Cheng, K.-Y.; Wingo, C.S.; Cain, B.D.; Gumz, M.L. Tissue-specific and time-dependent regulation of the endothelin axis by the circadian clock protein Per1. Life Sci. 2014, 118, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Berthold, H.; Münter, K.; Just, A.; Kirchheim, H.R.; Ehmke, H. Stimulation of the renin-angiotensin system by endothelin subtype A receptor blockade in conscious dogs. Hypertens. (Dallas Tex. 1979) 1999, 33, 1420–1424. [Google Scholar] [CrossRef]
- Santos, P.C.; Krieger, J.E.; Pereira, A.C. Renin-angiotensin system, hypertension, and chronic kidney disease: Pharmacogenetic implications. J. Pharmacol. Sci. 2012, 120, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Kobori, H.; Nangaku, M.; Navar, L.G.; Nishiyama, A. The intrarenal renin-angiotensin system: From physiology to the pathobiology of hypertension and kidney disease. Pharmacol. Rev. 2007, 59, 251–287. [Google Scholar] [CrossRef] [PubMed]
- Isobe, S.; Ohashi, N.; Fujikura, T.; Tsuji, T.; Sakao, Y.; Yasuda, H.; Kato, A.; Miyajima, H.; Fujigaki, Y. Disturbed circadian rhythm of the intrarenal renin-angiotensin system: Relevant to nocturnal hypertension and renal damage. Clin. Exp. Nephrol. 2015, 19, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Nishijima, Y.; Kobori, H.; Kaifu, K.; Mizushige, T.; Hara, T.; Nishiyama, A.; Kohno, M. Circadian rhythm of plasma and urinary angiotensinogen in healthy volunteers and in patients with chronic kidney disease. J. Renin-Angiotensin-Aldosterone Syst. 2014, 15, 505–508. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, K.; Kumar, V.; Koike, N.; Kim, T.-K.; Chong, J.; Buhr, E.D.; Whiteley, A.R.; Low, S.S.; Omura, C.; Fenner, D.; et al. Usf1, a suppressor of the circadian Clock mutant, reveals the nature of the DNA-binding of the CLOCK:BMAL1 complex in mice. elife 2013, 2, e00426. [Google Scholar] [CrossRef] [PubMed]
- Burnier, M.; Coltamai, L.; Maillard, M.; Bochud, M. Renal Sodium Handling and Nighttime Blood Pressure. Semin. Nephrol. 2007, 27, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Floris, M.; Pani, A.; Rosner, M.H. Sodium and volume disorders in advanced chronic kidney disease. Adv. Chronic Kidney Dis. 2016, 23, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Tokonami, N.; Mordasini, D.; Pradervand, S.; Centeno, G.; Jouffe, C.; Maillard, M.; Bonny, O.; Gachon, F.; Gomez, R.A.; Sequeira-Lopez, M.L.S.; et al. Local renal circadian clocks control fluid-electrolyte homeostasis and BP. J. Am. Soc. Nephrol. 2014, 25, 1430–1439. [Google Scholar] [CrossRef] [PubMed]
- Nikolaeva, S.; Pradervand, S.; Centeno, G.; Zavadova, V.; Tokonami, N.; Maillard, M.; Bonny, O.; Firsov, D. The circadian clock modulates renal sodium handling. J. Am. Soc. Nephrol. 2012, 23, 1019–1026. [Google Scholar] [CrossRef] [PubMed]
- Mercier Zuber, A.; Centeno, G.; Pradervand, S.; Nikolaeva, S.; Maquelin, L.; Onard Cardinaux, L.; Bonny, O.; Firsov, D. Molecular clock is involved in predictive circadian adjustment of renal function. Proc. Natl. Acad. Sci. USA 2009, 106, 16523–16528. [Google Scholar] [CrossRef] [PubMed]
- Krid, H.; Dorison, A.; Salhi, A.; Cheval, L.; Crambert, G.; Feraille, E. Expression profile of nuclear receptors along male mouse nephron segments reveals a link between ERRb and thick ascending limb function. PLoS ONE 2012, 7, e34223. [Google Scholar] [CrossRef] [PubMed]
- Saifur Rohman, M.; Emoto, N.; Nonaka, H.; Okura, R.; Nishimura, M.; Yagita, K.; Van Der Horst, G.T.J.; Matsuo, M.; Okamura, H.; Yokoyama, M. Circadian clock genes directly regulate expression of the Na+/H+ exchanger NHE3 in the kidney. Kidney Int. 2005, 67, 1410–1419. [Google Scholar] [CrossRef] [PubMed]
- Shull, G.E.; Schultheis, P.J.; Clarke, L.L.; Meneton, P.; Miller, M.L.; Soleimani, M.; Gawenis, L.R.; Riddle, T.M.; Duffy, J.J.; Doetschman, T.; et al. Renal and intestinal absorptive defects in mice lacking the NHE3 Na+/H+ exchanger. Nat. Genet. 1998, 19, 282–285. [Google Scholar] [CrossRef] [PubMed]
- Guyton, A. Blood pressure control-special role of the kidneys and body fluids. Science 1991, 252, 1813–1816. [Google Scholar] [CrossRef] [PubMed]
- Solocinski, K.; Richards, J.; All, S.; Cheng, K.-Y.; Khundmiri, S.J.; Gumz, M.L. Transcriptional regulation of NHE3 and SGLT1 by the circadian clock protein Per1 in proximal tubule cells. Am. J. Physiol. Ren. Physiol. 2015, 309, F933–F942. [Google Scholar] [CrossRef] [PubMed]
- Gumz, M.L.; Cheng, K.Y.; Lynch, I.J.; Stow, L.R.; Greenlee, M.M.; Cain, B.D.; Wingo, C.S. Regulation of αENaC expression by the circadian clock protein Period 1 in mpkCCDc14 cells. Biochim. Biophys. Acta Gene Regul. Mech. 2010, 1799, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Gumz, M.L.; Stow, L.R.; Lynch, I.J.; Creenlee, M.M.; Rudin, A.; Cain, B.D.; Weaver, D.R.; Wingo, C.S. The circadian clock protein Period1 regulates expression of the renal epithelial sodium channel in mice. J. Clin. Investig. 2009, 119, 2423–2434. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.; Cheng, K.-Y.; All, S.; Skopis, G.; Jeffers, L.; Jeanette Lynch, I.; Wingo, C.S.; Gumz, M.L. A role for the circadian clock protein Per1 in the regulation of aldosterone levels and renal Na+ retention. Am. J. Physiol. Ren. Physiol. 2013, 305, F1697–F1704. [Google Scholar] [CrossRef] [PubMed]
- Stow, L.R.; Richards, J.; Cheng, K.Y.; Lynch, I.J.; Jeffers, L.A.; Greenlee, M.M.; Cain, B.D.; Wingo, C.S.; Gumz, M.L. The circadian protein period 1 contributes to blood pressure control and coordinately regulates renal sodium transport genes. Hypertension 2012, 59, 1151–1156. [Google Scholar] [CrossRef] [PubMed]
- Kohagura, K.; Ohya, Y. Renal tubulointerstitial damage and salt-sensitive hypertension in chronic kidney disease: Is the tubulointerstitium relevant beyond the glomerulus? Hypertens. Res. 2015, 38, 102–103. [Google Scholar] [CrossRef] [PubMed]
- Hussein, A.M.; Malek, H.A.; Saad, M.-A. Renoprotective effects of aliskiren on adenine-induced tubulointerstitial nephropathy: Possible underlying mechanisms. Can. J. Physiol. Pharmacol. 2016, 94, 829–837. [Google Scholar] [CrossRef] [PubMed]
- Doi, M.; Takahashi, Y.; Komatsu, R.; Yamazaki, F.; Yamada, H.; Haraguchi, S.; Emoto, N.; Okuno, Y.; Tsujimoto, G.; Kanematsu, A.; et al. Salt-sensitive hypertension in circadian clock-deficient Cry-null mice involves dysregulated adrenal Hsd3b6. Nat. Med. 2010, 16, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.; Greenlee, M.I.M.; Jeffers, L.A.; Cheng, K.Y.; Guo, L.; Eaton, D.C.; Gumz, M.L. Inhibition of αENaC expression and ENaC activity following blockade of the circadian clock-regulatory kinases CK1δ/ε. Am. J. Physiol. Ren. Physiol. 2012, 303, F918–F927. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Zhang, H.; Yang, J.; Zhang, J.; Li, K.; Huo, B.; Dai, H.; Zhang, W.; Yang, J.; Tan, W.; et al. Nocturnal and circadian rhythm of blood pressure is associated with renal structure damage and function in patients with IgAN. Arch. Med. Res. 2016, 47, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Gast, H.; Gordic, S.; Petrzilka, S.; Lopez, M.; Müller, A.; Gietl, A.; Hock, C.; Birchler, T.; Fontana, A. Transforming growth factor-beta inhibits the expression of clock genes. Ann. N. Y. Acad. Sci. 2012, 1261, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Koch, B.C.P.; van der Putten, K.; Van Someren, E.J.W.; Wielders, J.P.M.; ter Wee, P.M.; Nagtegaal, J.E.; Gaillard, C.A. Impairment of endogenous melatonin rhythm is related to the degree of chronic kidney disease (CREAM study). Nephrol. Dial. Transplant. 2010, 25, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Rodriguez, A.; Abreu-Gonzalez, P.; Sanchez-Sanchez, J.J.; Kaski, J.C.; Reiter, R.J. Melatonin and circadian biology in human cardiovascular disease. J. Pineal Res. 2010, 49, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Rodriguez, A.; Abreu-Gonzalez, P.; Reiter, R.J. Melatonin and cardiovascular disease: Myth or reality? Rev. Española Cardiol. 2012, 65, 215–218. [Google Scholar] [CrossRef] [PubMed]
- Jonas, M.; Garfinkel, D.; Zisapel, N.; Laudon, M.; Grossman, E. Impaired nocturnal melatonin secretion in non-dipper hypertensive patients. Blood Press. 2003, 12, 19–24. [Google Scholar] [PubMed]
- Zeman, M.; Dulkova, K.; Bada, V.; Herichova, I. Plasma melatonin concentrations in hypertensive patients with the dipping and non-dipping blood pressure profile. Life Sci. 2005, 76, 1795–1803. [Google Scholar] [CrossRef] [PubMed]
- Takeda, N.; Maemura, K. Circadian clock and cardiovascular disease. J. Cardiol. 2011, 57, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Takeda, N.; Maemura, K. Chronobiology of acute myocardial infarction molecular biology. Biol. Rhythm Res. 2007, 38, 233–245. [Google Scholar] [CrossRef]
- Poirel, V.J.; Boggio, V.; Dardente, H.; Pevet, P.; Masson-Pevet, M.; Gauer, F. Contrary to other non-photic cues, acute melatonin injection does not induce immediate changes of clock gene mRNA expression in the rat suprachiasmatic nuclei. Neuroscience 2003, 120, 745–755. [Google Scholar] [CrossRef]
- Agez, L.; Laurent, V.; Pévet, P.; Masson-Pévet, M.; Gauer, F. Melatonin affects nuclear orphan receptors mRNA in the rat suprachiasmatic nuclei. Neuroscience 2007, 144, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Ingle, K.A.; Kain, V.; Goel, M.; Prabhu, S.D.; Young, M.E.; Halade, G.V. Cardiomyocyte-specific Bmal1 deletion in mice triggers diastolic dysfunction, extracellular matrix response, and impaired resolution of inflammation. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H1827–H1836. [Google Scholar] [CrossRef] [PubMed]
- Schroder, E.A.; Lefta, M.; Zhang, X.; Bartos, D.C.; Feng, H.-Z.; Zhao, Y.; Patwardhan, A.; Jin, J.-P.; Esser, K.A.; Delisle, B.P. The cardiomyocyte molecular clock, regulation of Scn5a, and arrhythmia susceptibility. Am. J. Physiol. Cell Physiol. 2013, 304, C954–C965. [Google Scholar] [CrossRef] [PubMed]
- Schroder, E.A.; Burgess, D.E.; Zhang, X.; Lefta, M.; Smith, J.L.; Patwardhan, A.; Bartos, D.C.; Elayi, C.S.; Esser, K.A.; Delisle, B.P. The cardiomyocyte molecular clock regulates the circadian expression of Kcnh2 and contributes to ventricular repolarization. Heart Rhythm 2015, 12, 1306–1314. [Google Scholar] [CrossRef] [PubMed]
- Durgan, D.J.; Tsai, J.-Y.; Grenett, M.H.; Pat, B.M.; Ratcliffe, W.F.; Villegas-Montoya, C.; Garvey, M.E.; Nagendran, J.; Dyck, J.R.B.; Bray, M.S.; et al. Evidence Suggesting that the Cardiomyocyte Circadian Clock Modulates Responsiveness of the Heart to Hypertrophic Stimuli in Mice. Chronobiol. Int. 2011, 28, 187–203. [Google Scholar] [CrossRef] [PubMed]
- Lefta, M.; Campbell, K.S.; Feng, H.-Z.; Jin, J.-P.; Esser, K.A. Development of dilated cardiomyopathy in Bmal1-deficient mice. Am. J. Physiol. Heart Circ. Physiol. 2012, 303, H475–H485. [Google Scholar] [CrossRef] [PubMed]
- Young, M.E.; Brewer, R.A.; Peliciari-Garcia, R.A.; Collins, H.E.; He, L.; Birky, T.L.; Peden, B.W.; Thompson, E.G.; Ammons, B.-J.; Bray, M.S.; et al. Cardiomyocyte-specific BMAL1 plays critical roles in metabolism, signaling, and maintenance of contractile function of the heart. J. Biol. Rhythm 2014, 29, 257–276. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhu, D.; Yuan, J.; Han, Z.; Wang, Y.; Qian, Z.; Hou, X.; Wu, T.; Zou, J. CLOCK-BMAL1 regulate the cardiac L-type calcium channel subunit CACNA1C through PI3K-Akt signaling pathway. Can. J. Physiol. Pharmacol. 2016, 94, 1023–1032. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Maillard, M.; Schibler, U.; Burnier, M.; Gachon, F. Cardiac hypertrophy, low blood pressure, and low aldosterone levels in mice devoid of the three circadian PAR bZip transcription factors DBP, HLF, and TEF. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 299, R1013–R1019. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Hitomi, H.; Nishiyama, A. Cardioprotective effects of SGLT2 inhibitors are possibly associated with normalization of the circadian rhythm of blood pressure. Hypertens. Res. 2017, 40, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Kittikulsuth, W.; Fujisawa, Y.; Sufiun, A.; Rafiq, K.; Hitomi, H.; Nakano, D.; Sohara, E.; Uchida, S.; Nishiyama, A. Effects of diuretics on sodium-dependent glucose cotransporter 2 inhibitor-induced changes in blood pressure in obese rats suffering from the metabolic syndrome. J. Hypertens. 2016, 34, 893–906. [Google Scholar] [CrossRef] [PubMed]
- Takeshige, Y.; Fujisawa, Y.; Rahman, A.; Kittikulsuth, W.; Nakano, D.; Mori, H.; Masaki, T.; Ohmori, K.; Kohno, M.; Ogata, H.; et al. A sodium-glucose co-transporter 2 inhibitor empagliflozin prevents abnormality of circadian rhythm of blood pressure in salt-treated obese rats. Hypertens. Res. 2016, 39, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Yanagi, M.; Tamura, K.; Fujikawa, T.; Wakui, H.; Kanaoka, T.; Ohsawa, M.; Azushima, K.; Maeda, A.; Kobori, H.; Umemura, S. The angiotensin II type 1 receptor blocker olmesartan preferentially improves nocturnal hypertension and proteinuria in chronic kidney disease. Hypertens. Res. 2013, 36, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Fuwa, D.; Fukuda, M.; Ogiyama, Y.; Sato, R.; Mizuno, M.; Miura, T.; Abe-Dohmae, S.; Michikawa, M.; Kobori, H.; Ohte, N. Addition of hydrochlorothiazide to angiotensin receptor blocker therapy can achieve a lower sodium balance with no acceleration of intrarenal renin angiotensin system in patients with chronic kidney disease. J. Renin-Angiotensin-Aldosterone Syst. 2016, 17, 505–508. [Google Scholar] [CrossRef] [PubMed]
- Fagard, R.H.; Thijs, L.; Staessen, J.A.; Clement, D.L.; De Buyzere, M.L.; De Bacquer, D.A. Night–day blood pressure ratio and dipping pattern as predictors of death and cardiovascular events in hypertension. J. Hum. Hypertens. 2009, 23, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Hermida, R.C.; Ayala, D.E.; Mojon, A.; Fernandez, J.R. Influence of Time of Day of Blood Pressure-Lowering Treatment on Cardiovascular Risk in Hypertensive Patients With Type 2 Diabetes. Diabetes Care 2011, 34, 1270–1276. [Google Scholar] [CrossRef] [PubMed]
- Hermida, R.C.; Ayala, D.E.; Mojon, A.; Fernandez, J.R. Bedtime Dosing of Antihypertensive Medications Reduces Cardiovascular Risk in CKD. J. Am. Soc. Nephrol. 2011, 22, 2313–2321. [Google Scholar] [CrossRef] [PubMed]
- Roush, G.C.; Fapohunda, J.; Kostis, J.B. Evening Dosing of Antihypertensive Therapy to Reduce Cardiovascular Events: A Third Type of Evidence Based on a Systematic Review and Meta-Analysis of Randomized Trials. J. Clin. Hypertens. 2014, 16, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Bonten, T.N.; Saris, A.; van Oostrom, M.J.; Snoep, J.D.; Rosendaal, F.R.; Zwaginga, J.; Eikenboom, J.; van der Meer, P.F.; van der Bom, J.G. Effect of aspirin intake at bedtime versus on awakening on circadian rhythm of platelet reactivity. Thromb. Haemost. 2014, 112, 1209–1218. [Google Scholar] [CrossRef] [PubMed]
- Bonten, T.N.; Snoep, J.D.; Assendelft, W.J.J.; Zwaginga, J.J.; Eikenboom, J.; Huisman, M.V.; Rosendaal, F.R.; van der Bom, J.G. Time-dependent effects of aspirin on blood pressure and morning platelet reactivity. Hypertension 2015, 65, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Muller, J.E.; Stone, P.H.; Turi, Z.G.; Rutherford, J.D.; Czeisler, C.A.; Parker, C.; Poole, W.K.; Passamani, E.; Roberts, R.; Robertson, T.; et al. Circadian variation in the frequency of onset of acute myocardial infarction. N. Engl. J. Med. 1985, 313, 1315–1322. [Google Scholar] [CrossRef] [PubMed]
- Schloss, M.J.; Horckmans, M.; Nitz, K.; Duchene, J.; Drechsler, M.; Bidzhekov, K.; Scheiermann, C.; Weber, C.; Soehnlein, O.; Steffens, S. The time-of-day of myocardial infarction onset affects healing through oscillations in cardiac neutrophil recruitment. EMBO Mol. Med. 2016, 8, 937–948. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Sun, Y.; Xu, T.; Sun, L.; Liu, L.; Sun, W.; Feng, X.; Ma, J.; Wang, L.; Yao, L. Effect of nocturnal hemodialysis versus conventional hemodialysis on end-stage renal disease: A meta-analysis and systematic review. PLoS ONE 2017, 12, e0169203. [Google Scholar] [CrossRef] [PubMed]
- Tsimakouridze, E.V.; Alibhai, F.J.; Martino, T.A. Therapeutic applications of circadian rhythms for the cardiovascular system. Front. Pharmacol. 2015, 6, 77. [Google Scholar] [CrossRef] [PubMed]
- Floras, J.S. Sleep apnea and cardiovascular risk. J. Cardiol. 2014, 63, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Marin, J.M.; Carrizo, S.J.; Vicente, E.; Agusti, A.G. Long-term cardiovascular outcomes in men with obstructive sleep apnoea-hypopnoea with or without treatment with continuous positive airway pressure: An observational study. Lancet 2005, 365, 1046–1053. [Google Scholar] [CrossRef]
- Sharma, S.K.; Agrawal, S.; Damodaran, D.; Sreenivas, V.; Kadhiravan, T.; Lakshmy, R.; Jagia, P.; Kumar, A. CPAP for the metabolic syndrome in patients with obstructive sleep apnea. N. Engl. J. Med. 2011, 365, 2277–2286. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X. Melatonin and cardiac pathophysiology. Heart Metab. 2009, 44, 31–34. [Google Scholar]
- Reiter, R.J.; Tan, D.-X.; Korkmaz, A. The circadian melatonin rhythm and its modulation: Possible impact on hypertension. J. Hypertens. 2009, 27, S17–S20. [Google Scholar] [CrossRef] [PubMed]
- Scheer, F.A.; Van Montfrans, G.A.; van Someren, E.J.W.; Mairuhu, G.; Buijs, R.M. Daily nighttime melatonin reduces blood pressure in male patients with essential hypertension. Hypertension 2004, 43, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.M.; Chen, H.R.; Hsiao, G.; Sheu, J.R.; Wang, J.J.; Yen, M.H. Protective effects of melatonin on myocardial ischemia/reperfusion injury in vivo. J. Pineal Res. 2002, 33, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Sahna, E.; Olmez, E.; Acet, A. Effects of physiological and pharmacological concentrations of melatonin on ischemia-reperfusion arrhythmias in rats: Can the incidence of sudden cardiac death be reduced. J. Pineal Res. 2002, 32, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Hirota, T.; Lee, J.W.; Lewis, W.G.; Zhang, E.E.; Breton, G.; Liu, X.; Garcia, M.; Peters, E.C.; Etchegaray, J.-P.; Traver, D.; et al. High-Throughput Chemical Screen Identifies a Novel Potent Modulator of Cellular Circadian Rhythms and Reveals CKIα as a Clock Regulatory Kinase. PLoS Biol. 2010, 8, e1000559. [Google Scholar] [CrossRef] [PubMed]
- Isojima, Y.; Nakajima, M.; Ukai, H.; Fujishima, H.; Yamada, R.G.; Masumoto, K.-H.; Kiuchi, R.; Ishida, M.; Ukai-Tadenuma, M.; Minami, Y.; et al. CKI/-dependent phosphorylation is a temperature-insensitive, period-determining process in the mammalian circadian clock. Proc. Natl. Acad. Sci. USA 2009, 106, 15744–15749. [Google Scholar] [CrossRef] [PubMed]
- Hirota, T.; Lee, J.W.; St. John, P.C.; Sawa, M.; Iwaisako, K.; Noguchi, T.; Pongsawakul, P.Y.; Sonntag, T.; Welsh, D.K.; Brenner, D.A.; et al. Identification of Small Molecule Activators of Cryptochrome. Science 2012, 337, 1094–1097. [Google Scholar] [CrossRef] [PubMed]
- Kojetin, D.J.; Burris, T.P. REV-ERB and ROR nuclear receptors as drug targets. Nat. Rev. Drug Discov. 2014, 13, 197–216. [Google Scholar] [CrossRef] [PubMed]
- Solt, L.A.; Wang, Y.; Banerjee, S.; Hughes, T.; Kojetin, D.J.; Lundasen, T.; Shin, Y.; Liu, J.; Cameron, M.D.; Noel, R.; et al. Regulation of circadian behaviour and metabolism by synthetic REV-ERB agonists. Nature 2012, 485, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Trump, R.P.; Bresciani, S.; Cooper, A.W.J.; Tellam, J.P.; Wojno, J.; Blaikley, J.; Orband-Miller, L.A.; Kashatus, J.A.; Boudjelal, M.; Dawson, H.C.; et al. Optimized chemical probes for REV-ERBα. J. Med. Chem. 2013, 56, 4729–4737. [Google Scholar] [CrossRef] [PubMed]
- Kojetin, D.; Wang, Y.; Kamenecka, T.M.; Burris, T.P. Identification of SR8278, a Synthetic Antagonist of the Nuclear Heme Receptor REV-ERB. ACS Chem. Biol. 2011, 6, 131–134. [Google Scholar] [CrossRef] [PubMed]
- Vieira, E.; Marroquí, L.; Batista, T.M.; Caballero-Garrido, E.; Carneiro, E.M.; Boschero, A.C.; Nadal, A.; Quesada, I. The Clock Gene Rev-erb α Regulates Pancreatic β-Cell Function: Modulation by Leptin and High-Fat Diet. Endocrinology 2012, 153, 592–601. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Kumar, N.; Nuhant, P.; Cameron, M.D.; Istrate, M.A.; Roush, W.R.; Griffin, P.R.; Burris, T.P. Identification of SR1078, a Synthetic Agonist for the Orphan Nuclear Receptors RORα and RORγ. ACS Chem. Biol. 2010, 5, 1029–1034. [Google Scholar] [CrossRef] [PubMed]
- Kharitonenkov, A.; Wroblewski, V.J.; Koester, A.; Chen, Y.F.; Clutinger, C.K.; Tigno, X.T.; Hansen, B.C.; Shanafelt, A.B.; Etgen, G.J. The metabolic state of diabetic monkeys is regulated by fibroblast growth factor-21. Endocrinology 2007, 148, 774–781. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Kojetin, D.J.; Solt, L.A.; Kumar, K.G.; Nuhant, P.; Duckett, D.R.; Cameron, M.D.; Butler, A.A.; Roush, W.R.; Griffin, P.R.; et al. Identification of SR3335 (ML-176): A Synthetic RORα Selective Inverse Agonist. ACS Chem. Biol. 2011, 6, 218–222. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Experimental Model | Molecular Mechanism | Altered Physiological Functions | References |
|---|---|---|---|
| db/db mice (type 2 diabetic model) | Attenuated circadian oscillation of Bmal1 and Dbp | Hypertension, disrupted BP circadian rhythm | Su et al. 2008 [17] |
| db/db mice | Altered clock gene expression both in central (Per1) and peripheral clock (Per1/2, Cry1/2), as well as their target genes (Dbp, Pparγ, Rev-erbα) | Disrupted diurnal contraction of vasculature | Su et al. 2012 [18] |
| High-fat-diet-induced obesity in mice | Altered circadian oscillation of clock (Per1-3; Cry1/2; Bmal1) and clock-controlled (Dbp, E4bp4, Pdk4, Pepck, Nhe3) genes | Hyperglycemia, hypercholesterolemia, hyperinsulinemia | Hsieh et al. 2010 [19] |
| db/db mice | Altered rhythmic expression of Per2 and Bmal1, increased Pai-1 mRNA expression | Attenuated circadian behavior | Kudo et al. 2004 [20] |
| Streptozotocin (STZ)-induced diabetes in Clock-mutant mice | Circadian augmentation of Pai-1 is diminished | Clock is involved in the diabetes-induced circadian augmentation of Pai-1 | Oishi et al. 2005 [21] |
| Dim light at night (alteration of circadian timing system) in mice | Attenuated circadian oscillation of Per1 and Per2 in hypothalamus, and Rev-Erb in liver and adipose tissue | Onset and progression of metabolic abnormalities | Fonken et al. 2011 [22] |
| Human subjects with type 2 diabetes (T2D) | BMAL1 single nucleotide polymorphisms (SNPs) (2 BMAL1 haplotypes) | SNP is associated with hypertension and T2D | Woon et al. 2007 [23] |
| Human subjects with T2D | CLOCK-rs4580704 (C > G) SNP | Cardiovascular (CV) disorders | Corella et al. 2016 [24] |
| Human subjects with rotating shiftwork | Interaction (epistatic effect) of serotonin transporter and CLOCK gene variation | Metabolic abnormalities | Sookoian et al. 2010 [25] |
| STZ-induced diabetes in mice | Altered circadian oscillation of core clock and their target genes | Contractile dysfunction of the heart | Young et al. 2002 [26] |
| Experimental Model | Molecular Mechanism | Altered Physiological Functions | References |
|---|---|---|---|
| 5/6 nephrectomy in rats | Altered localization and diurnal variation of BMAL1, DBP, and PER2 in the kidney | Altered diurnal rhythm of renal function | Huang et al. 2013 [72] |
| Unilateral ureteral obstruction (UUO) in mice | Agtrap ⬇, Usf 1 ⬆, Usf2 ⬇ | Renal dysfunction | Matsuda et al. 2013 [73] |
| 5/6 nephrectomy in rats | Altered diurnal expression of clock genes (Per2, Dbp, and Bmal1), plasma renin activity, angiotensin II, and aldosterone | Severe kidney injury and altered diurnal rhythm of BP | Huang et al. 2013 [74] |
| 5/6 nephrectomy in mice | Dbp ⬇, Cyp3a11 ⬇ and Cyp26a1 ⬇, Tgf-β1 ⬆ | Altered hepatic metabolism aggravates renal dysfunction | Hamamura et al. 2016 [75] |
| UUO in Clock-KO mice | Cyclooxygenase 2 ⬆, collagen synthesis ⬆, oxidative stress ⬆, Tgf-β ⬆ | Renal dysfunction | Chen et al. 2015 [76] |
| 5/6 nephrectomy in rats | Altered circadian oscillation of Per1 and 2 in the hypothalamus | Sleep disturbance associated with CKD | Hsu et al. 2012 [77] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, A.; Hasan, A.U.; Nishiyama, A.; Kobori, H. Altered Circadian Timing System-Mediated Non-Dipping Pattern of Blood Pressure and Associated Cardiovascular Disorders in Metabolic and Kidney Diseases. Int. J. Mol. Sci. 2018, 19, 400. https://doi.org/10.3390/ijms19020400
Rahman A, Hasan AU, Nishiyama A, Kobori H. Altered Circadian Timing System-Mediated Non-Dipping Pattern of Blood Pressure and Associated Cardiovascular Disorders in Metabolic and Kidney Diseases. International Journal of Molecular Sciences. 2018; 19(2):400. https://doi.org/10.3390/ijms19020400
Chicago/Turabian StyleRahman, Asadur, Arif Ul Hasan, Akira Nishiyama, and Hiroyuki Kobori. 2018. "Altered Circadian Timing System-Mediated Non-Dipping Pattern of Blood Pressure and Associated Cardiovascular Disorders in Metabolic and Kidney Diseases" International Journal of Molecular Sciences 19, no. 2: 400. https://doi.org/10.3390/ijms19020400
APA StyleRahman, A., Hasan, A. U., Nishiyama, A., & Kobori, H. (2018). Altered Circadian Timing System-Mediated Non-Dipping Pattern of Blood Pressure and Associated Cardiovascular Disorders in Metabolic and Kidney Diseases. International Journal of Molecular Sciences, 19(2), 400. https://doi.org/10.3390/ijms19020400