The Roles of Mitochondrion in Intergenomic Gene Transfer in Plants: A Source and a Pool


Abstract
:1. Introduction
2. Results and Discussion
2.1. The Role of Mitochondrion as a Gene Source: Intergenomic Gene Transfer from Mitochondrion
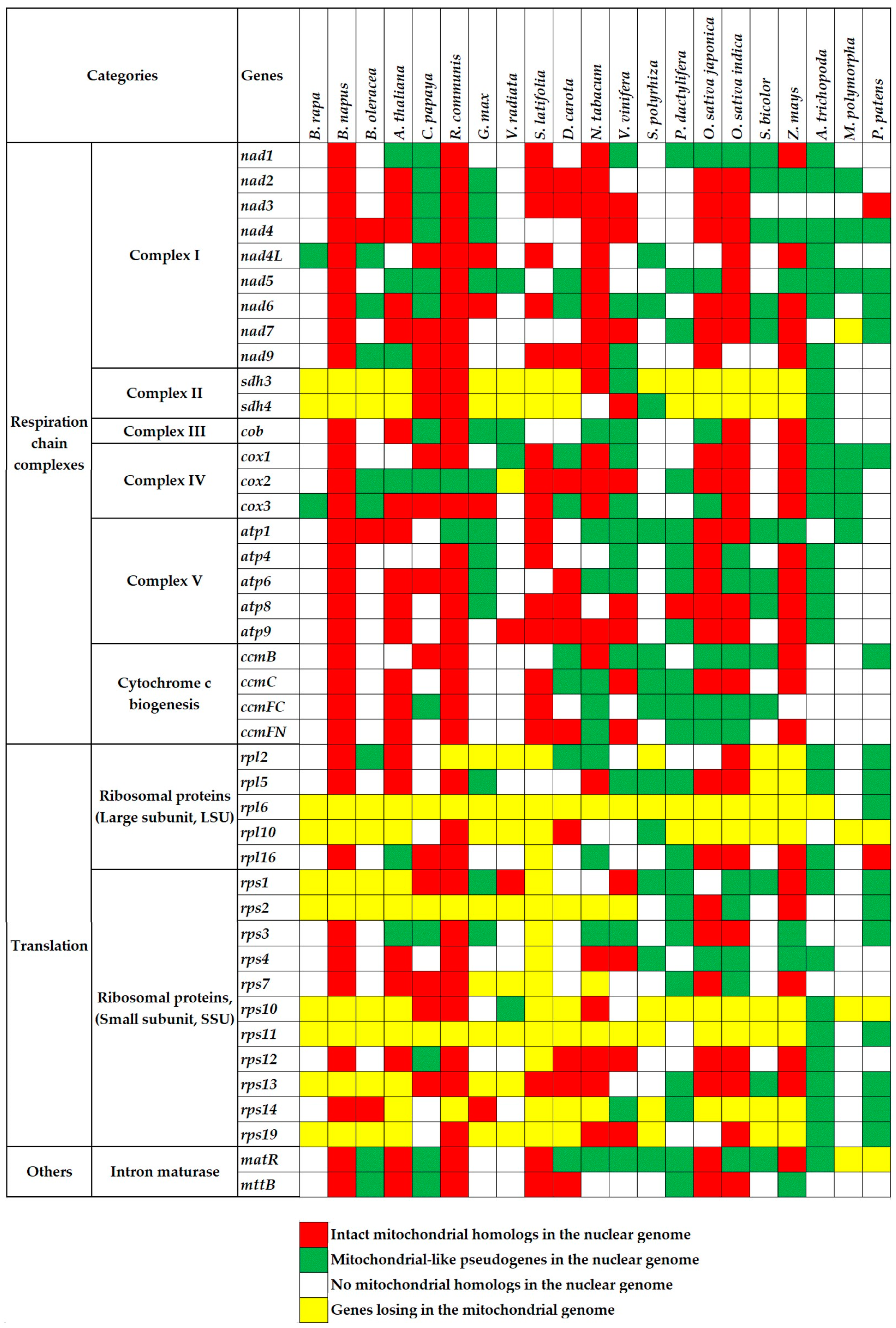
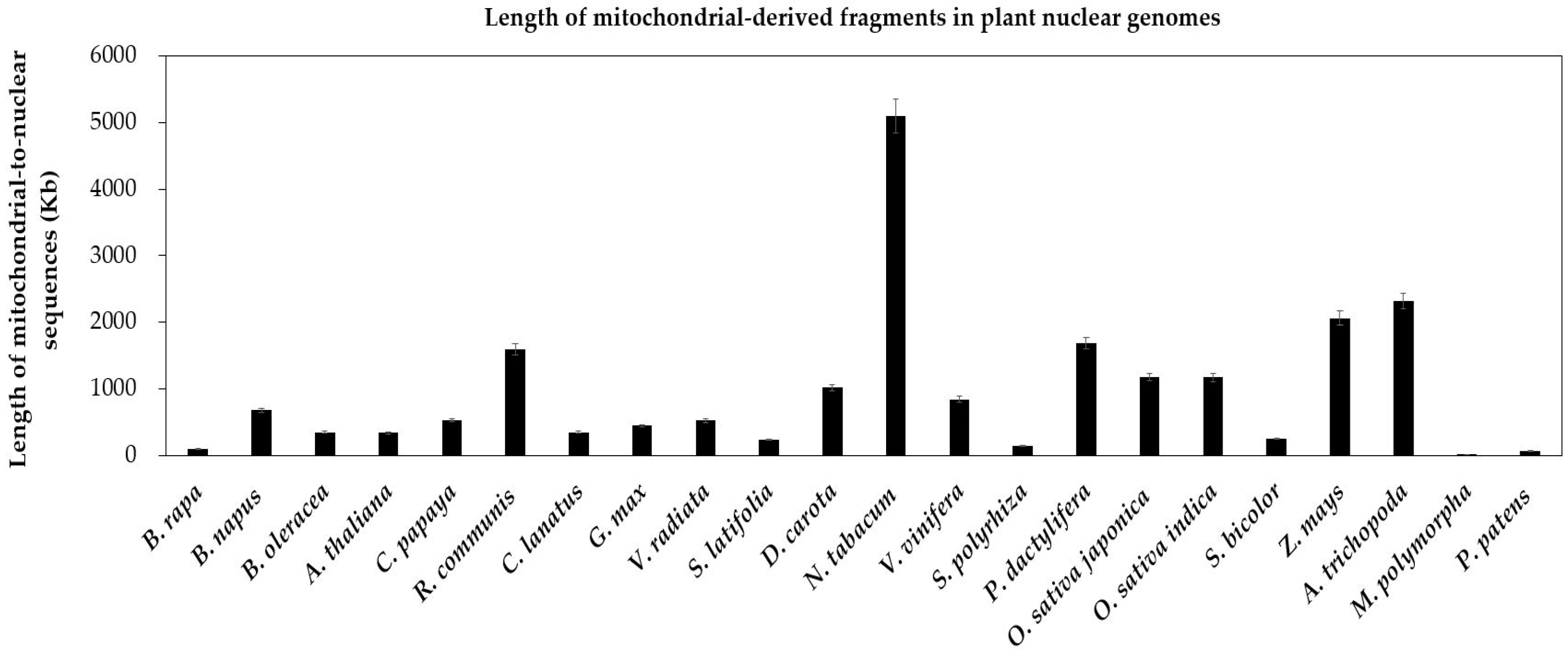
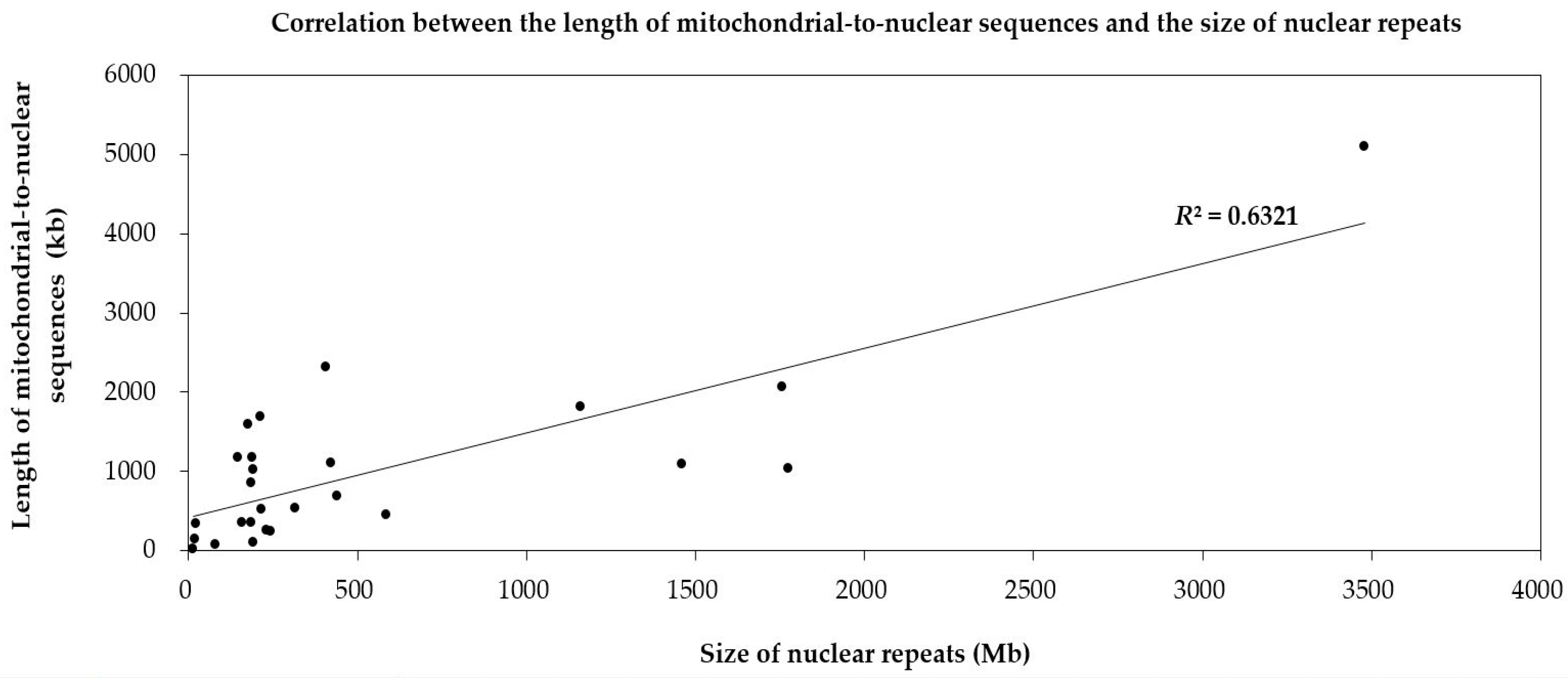
2.1.1. Intergenomic Gene Transfer from Mitochondrion to Nucleus
2.1.2. Intergenomic Gene Transfer from Mitochondrion to Chloroplast
2.2. The Role of Mitochondrion as a Gene Pool: Intergenomic Gene Transfer into Mitochondrion
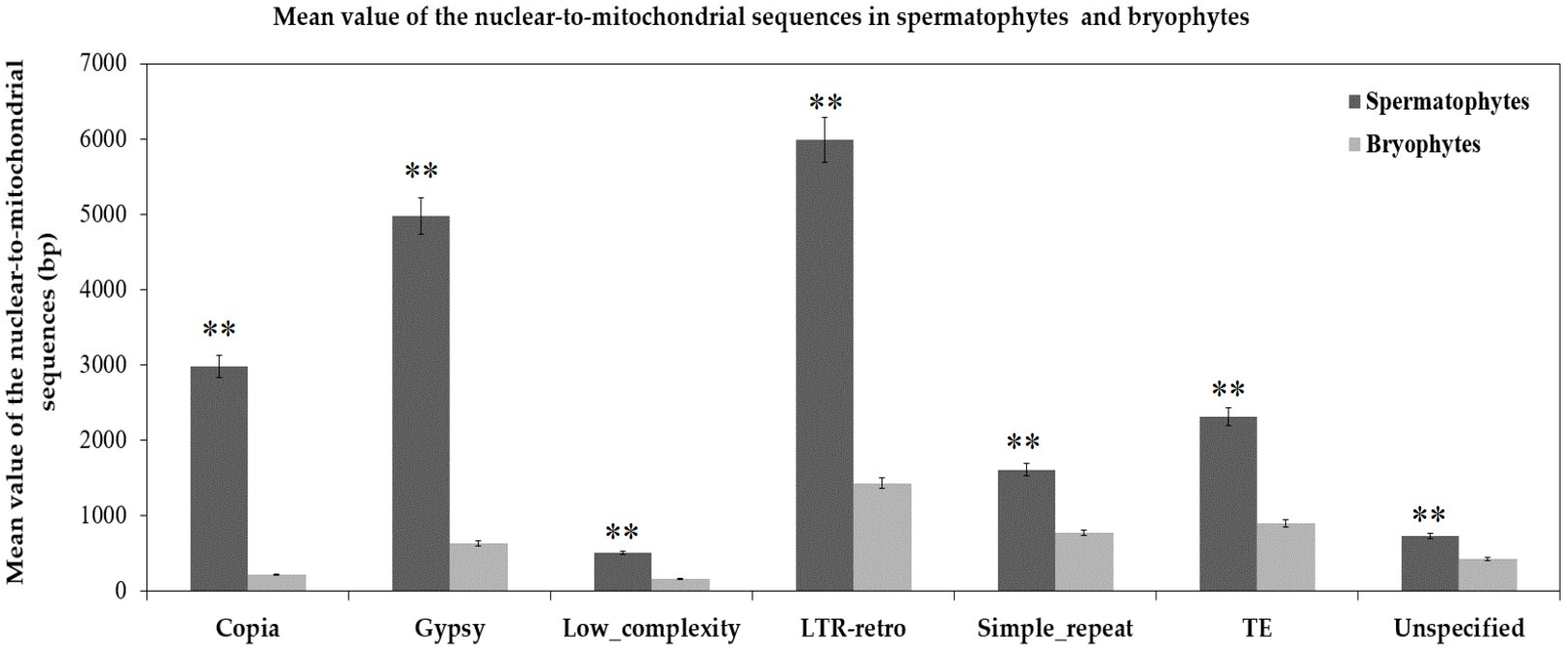
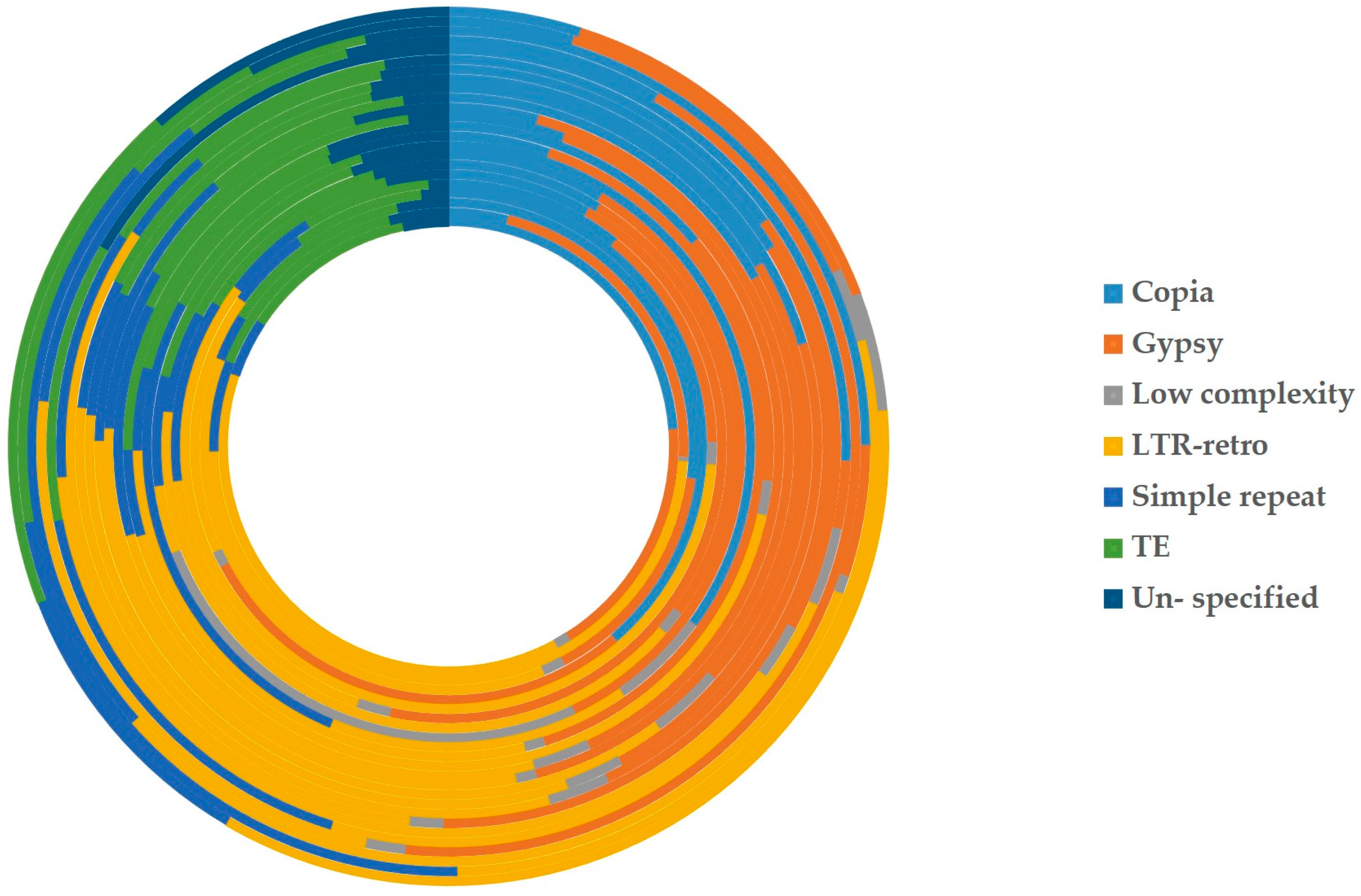
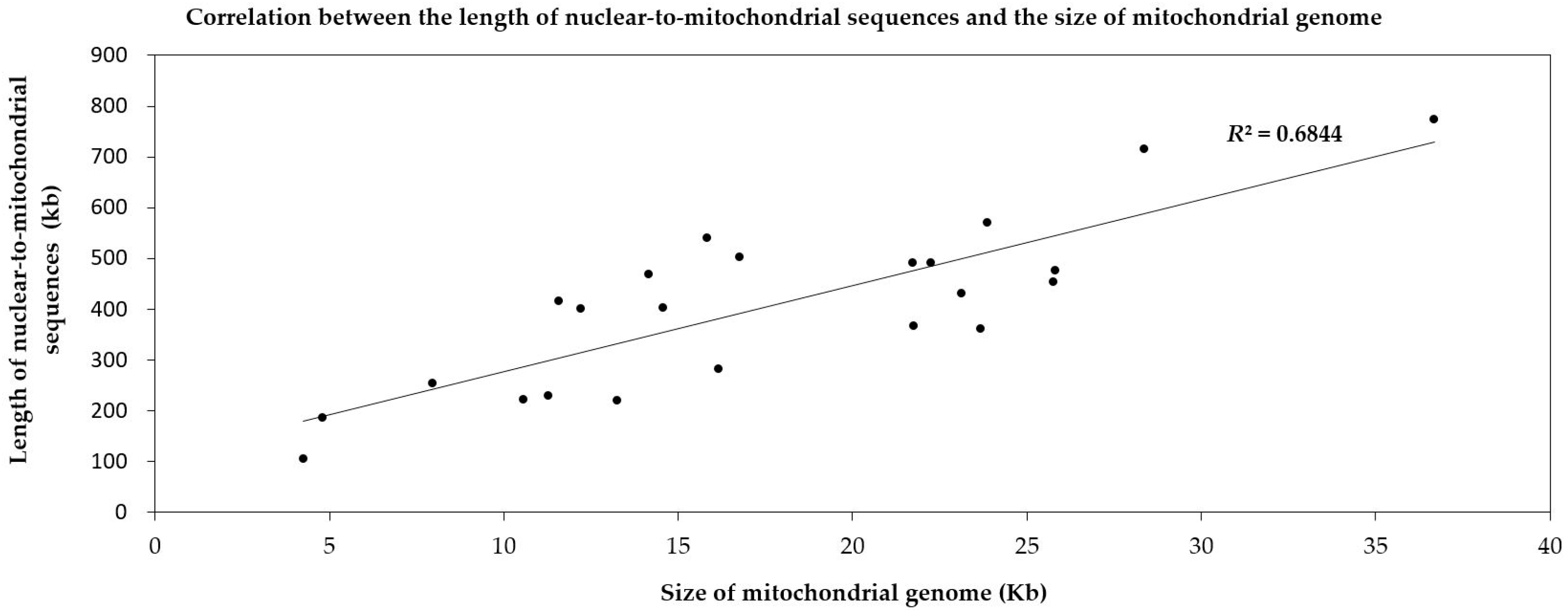
2.2.1. Intergenomic Gene Transfer from Nucleus to Mitochondrion
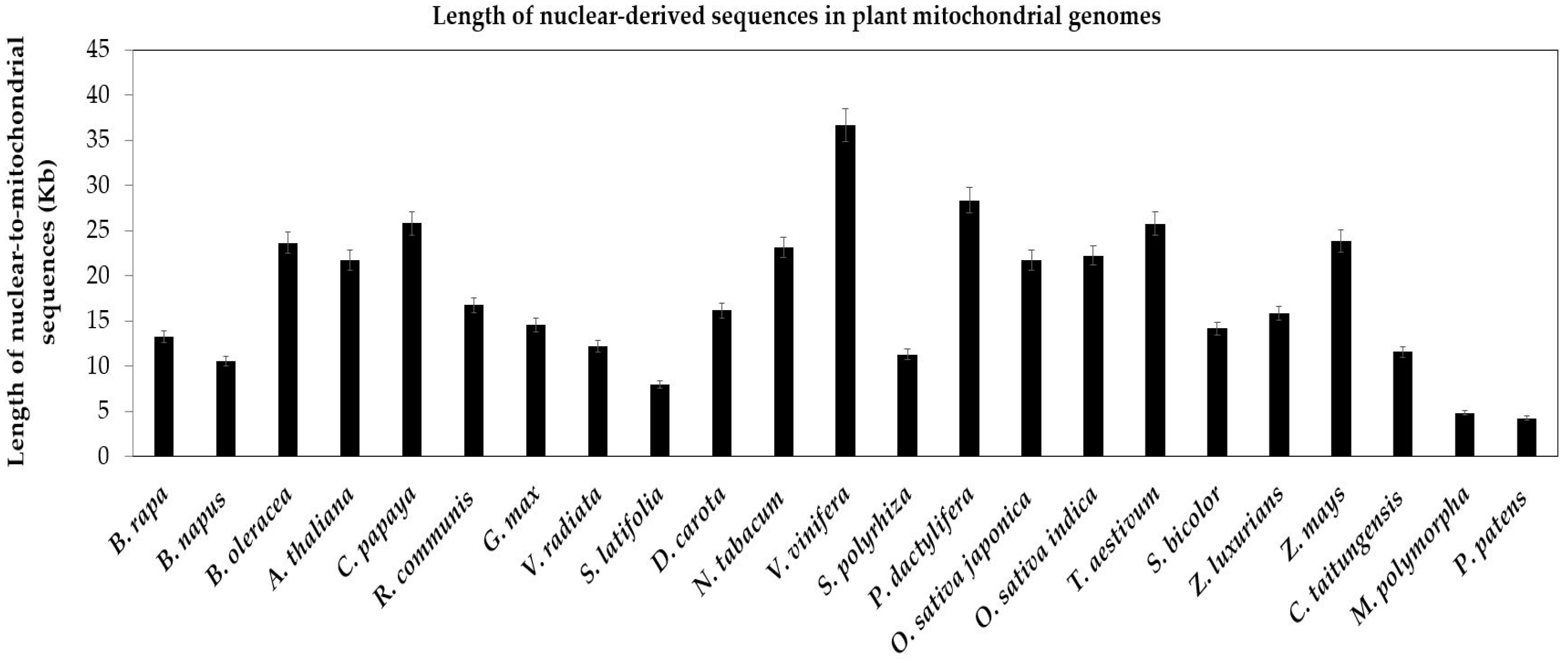
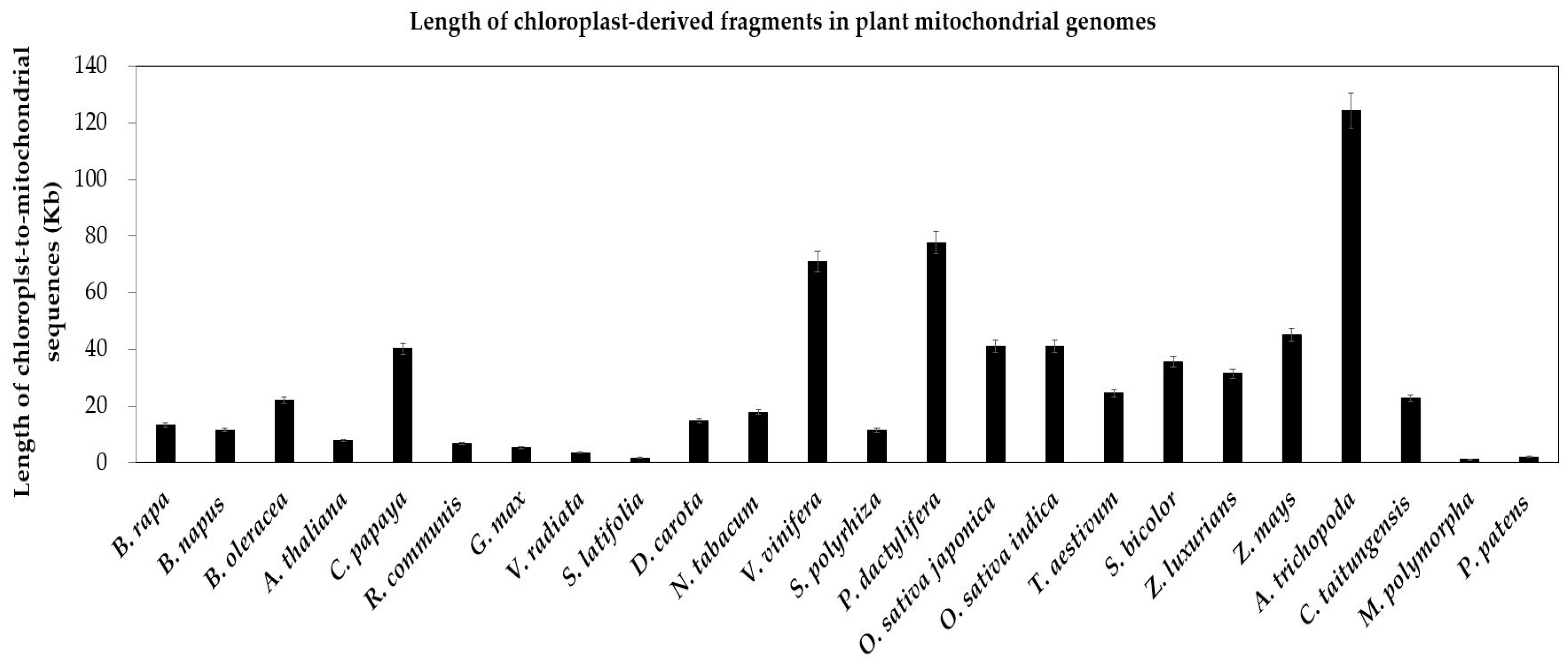
2.2.2. Intergenomic Gene Transfer from Chloroplast to Mitochondrion
3. Materials and Methods
3.1. Availability of Chloroplast, Mitochondrial and Nuclear Genomes
3.2. Detection of Total Intergenomic-Transfer DNA Sequences
3.3. Identification of Intergenomic-Transfer Homologies
3.4. Detection of the Repeats in Mitochondrial Genomes
3.5. NHEJ Analysis
3.6. Phylogenetic Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gray, M.W. Mosaic nature of the mitochondrial proteome: Implications for the origin and evolution of mitochondria. Proc. Natl. Acad. Sci. USA 2015, 112, 10133–10138. [Google Scholar] [CrossRef] [PubMed]
- Bock, R. Witnessing genome evolution: Experimental reconstruction of endosymbiotic and horizontal gene transfer. Annu. Rev. Genet. 2017, 51, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Stern, D.B.; Lonsdale, D.M. Mitochondrial and chloroplast genomes of maize have a 12-kilobase DNA-sequence in common. Nature 1982, 299, 698–702. [Google Scholar] [CrossRef] [PubMed]
- Knoop, V.; Unseld, M.; Marienfeld, J.; Brandt, P.; Sunkel, S.; Ullrich, H.; Brennicke, A. Copia-, gypsy- and line-like retrotransposon fragments in the mitochondrial genome of Arabidopsis thaliana. Genetics 1996, 142, 579–585. [Google Scholar] [PubMed]
- Wang, D.; Rousseau-Gueutin, M.; Timmis, J.N. Plastid sequences contribute to some plant mitochondrial genes. Mol. Biol. Evol. 2012, 29, 1707–1711. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.Z.; Cao, D.D.; Li, S.S.; Su, A.G.; Geng, J.N.; Grover, C.E.; Hu, S.N.; Hua, J.P. The complete mitochondrial genome of Gossypium hirsutum and evolutionary analysis of higher plant mitochondrial genomes. PLoS ONE 2013, 8, e69476. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.Y.; Chen, Z.W.; Grover, C.E.; Wang, Y.M.; Li, S.S.; Liu, G.Z.; Ma, Z.Y.; Wendel, J.F.; Hua, J.P. Rapid evolutionary divergence of Gossypium barbadense and G. hirsutum mitochondrial genomes. BMC Genom. 2015, 16, 770. [Google Scholar] [CrossRef] [PubMed]
- Notsu, Y.; Masood, S.; Nishikawa, T.; Kubo, N.; Akiduki, G.; Nakazono, M.; Hirai, A.; Kadowaki, K. The complete sequence of the rice (Oryza sativa L.) mitochondrial genome: Frequent DNA sequence acquisition and loss during the evolution of flowering plants. Mol. Genet. Genom. 2002, 268, 434–445. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wu, Y.W.; Shih, A.C.C.; Wu, C.S.; Wang, Y.N.; Chaw, S.M. Transfer of chloroplast genomic DNA to mitochondrial genome occurred at least 300 mya. Mol. Biol. Evol. 2007, 24, 2040–2048. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Moreno, L.; Gonzalez, V.M.; Benjak, A.; Marti, M.C.; Puigdomenech, P.; Aranda, M.A.; Garcia-Mas, J. Determination of the melon chloroplast and mitochondrial genome sequences reveals that the largest reported mitochondrial genome in plants contains a significant amount of DNA having a nuclear origin. BMC Genom. 2011, 12, 424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alverson, A.J.; Rice, D.W.; Dickinson, S.; Barry, K.; Palmer, J.D. Origins and recombination of the bacterial-sized multichromosomal mitochondrial genome of cucumber. Plant Cell 2011, 23, 2499–2513. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.W.; Nie, H.S.; Grover, C.E.; Wang, Y.M.; Li, P.; Wang, M.Y.; Pei, H.L.; Zhao, Y.P.; Li, S.S.; Wendel, J.F.; et al. Entire nucleotide sequences of Gossypium raimondii and G. arboreum mitochondrial genomes revealed a-genome species as cytoplasmic donor of the allotetraploid species. Plant Biol. 2017, 19, 484–493. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, A.; Small, I.; Cosset, A.; Weil, J.H.; Marechal-Drouard, L. Editing and import: Strategies for providing plant mitochondria with a complete set of functional transfer rnas. Biochimie 1996, 78, 518–529. [Google Scholar] [CrossRef]
- Clifton, S.W.; Minx, P.; Fauron, C.M.R.; Gibson, M.; Allen, J.O.; Sun, H.; Thompson, M.; Barbazuk, W.B.; Kanuganti, S.; Tayloe, C.; et al. Sequence and comparative analysis of the maize NB mitochondrial genome. Plant Physiol. 2004, 136, 3486–3503. [Google Scholar] [CrossRef] [PubMed]
- Sloan, D.B.; Alverson, A.J.; Storchova, H.; Palmer, J.D.; Taylor, D.R. Extensive loss of translational genes in the structurally dynamic mitochondrial genome of the angiosperm Silene latifolia. BMC Evol. Biol. 2010, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Iorizzo, M.; Senalik, D.; Szklarczyk, M.; Grzebelus, D.; Spooner, D.; Simon, P. De novo assembly of the carrot mitochondrial genome using next generation sequencing of whole genomic DNA provides first evidence of DNA transfer into an angiosperm plastid genome. BMC Plant Biol. 2012, 12, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Straub, S.C.K.; Cronn, R.C.; Edwards, C.; Fishbein, M.; Liston, A. Horizontal transfer of DNA from the mitochondrial to the plastid genome and its subsequent evolution in milkweeds (Apocynaceae). Genome Biol. Evol. 2013, 5, 1872–1885. [Google Scholar] [CrossRef] [PubMed]
- Iorizzo, M.; Grzebelus, D.; Senalik, D.; Szklarczyk, M.; Spooner, D.; Simon, P. Against the traffic: The first evidence for mitochondrial DNA transfer into the plastid genome. Mob. Genet. Elem. 2012, 2, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.F.; Erez, A.; Nagamani, S.C.S.; Dhar, S.U.; Kolodziejska, K.E.; Dharmadhikari, A.V.; Cooper, M.L.; Wiszniewska, J.; Zhang, F.; Withers, M.A.; et al. Chromosome catastrophes involve replication mechanisms generating complex genomic rearrangements. Cell 2011, 146, 888–902. [Google Scholar] [CrossRef] [PubMed]
- Hastings, P.J.; Lupski, J.R.; Rosenberg, S.M.; Ira, G. Mechanisms of change in gene copy number. Nat. Rev. Genet. 2009, 10, 551–564. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.; Stoebe, B.; Goremykin, V.; Hansmann, S.; Hasegawa, M.; Kowallik, K.V. Gene transfer to the nucleus and the evolution of chloroplasts. Nature 1998, 393, 162–165. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, J.L.; Schmidt, G.W. Pervasive migration of organellar DNA to the nucleus in plants. J. Mol. Evol. 1995, 41, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Bergthorsson, U.; Adams, K.L.; Thomason, B.; Palmer, J.D. Widespread horizontal transfer of mitochondrial genes in flowering plants. Nature 2003, 424, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Ku, C.; Nelson-Sathi, S.; Roettger, M.; Sousa, F.L.; Lockhart, P.J.; Bryant, D.; Hazkani-Covo, E.; McInerney, J.O.; Landan, G.; Martin, W.F. Endosymbiotic origin and differential loss of eukaryotic genes. Nature 2015, 524, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.W.; Nie, H.S.; Wang, Y.M.; Pei, H.L.; Li, S.S.; Zhang, L.D.; Hua, J.P. Rapid evolutionary divergence of diploid and allotetraploid gossypium mitochondrial genomes. BMC Genom. 2017, 18, 876. [Google Scholar] [CrossRef] [PubMed]
- Bock, R.; Timmis, J.N. Reconstructing evolution: Gene transfer from plastids to the nucleus. BioEssays 2008, 30, 556–566. [Google Scholar] [CrossRef] [PubMed]
- Bonnefoy, N.; Remacle, C.; Fox, T.D. Genetic transformation of saccharomyces cerevisiae and chlamydomonas reinhardtii mitochondria. Methods Cell Biol. 2007, 80, 525–548. [Google Scholar] [PubMed]
- Covello, P.; Gray, M.W. Silent mitochondrial and active nuclear genes for subunit 2 of cytochrome c oxidase (cox2) in soybean: Evidence for rna-mediated gene transfer. EMBO J. 1992, 11, 3815–3820. [Google Scholar] [PubMed]
- Nugent, J.M.; Palmer, J.D. RNA-mediated transfer of the gene coxII from the mitochondrion to the nucleus during flowering plant evolution. Cell 1991, 66, 473–481. [Google Scholar] [CrossRef]
- Wang, D.; Timmis, J.N. Cytoplasmic organelle DNA preferentially inserts into open chromatin. Genome Biol. Evol. 2013, 5, 1060–1064. [Google Scholar] [CrossRef] [PubMed]
- Kudla, J.; Albertazzi, F.; Blazević, D.; Hermann, M.; Bock, R. Loss of the mitochondrial cox2 intron 1 in a family of monocotyledonous plants and utilization of mitochondrial intron sequences for the construction of a nuclear intron. Mol. Genet. Genom. 2002, 267, 223–230. [Google Scholar]
- Kadowaki, K.-I.; Kubo, N.; Ozawa, K.; Hirai, A. Targeting presequence acquisition after mitochondrial gene transfer to the nucleus occurs by duplication of existing target signals. EMBO J. 1997, 15, 6652–6661. [Google Scholar]
- Adams, K.L.; Qiu, Y.L.; Stoutemyer, M.; Palmer, J.D. Punctuated evolution of mitochondrial gene content: High and variable rates of mitochondrial gene loss and transfer to the nucleus during angiosperm evolution. Proc. Natl. Acad. Sci. USA 2002, 99, 9905–9912. [Google Scholar] [CrossRef] [PubMed]
- Adams, K.L.; Palmer, J.D. Evolution of mitochondrial gene content: Gene loss and transfer to the nucleus. Mol. Phylogenet. Evol. 2003, 29, 380–395. [Google Scholar] [CrossRef]
- Wang, D.; Qu, Z.P.; Adelson, D.L.; Zhu, J.K.; Timmis, J.N. Transcription of nuclear organellar DNA in a model plant system. Genome Biol. Evol. 2014, 6, 1327–1334. [Google Scholar] [CrossRef] [PubMed]
- Goremykin, V.V.; Salamini, F.; Velasco, R.; Viola, R. Mitochondrial DNA of Vitis vinifera and the issue of rampant horizontal gene transfer. Mol. Biol. Evol. 2009, 26, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.R. Mitochondrion-to-plastid DNA transfer: It happens. New Phytol. 2014, 202, 736–738. [Google Scholar] [CrossRef] [PubMed]
- Downie, S.; Jansen, R. A comparative analysis of whole plastid genomes from the apiales: Expansion and contraction of the inverted repeat, mitochondrial to plastid transfer of DNA and identification of highly divergent noncoding regions. Syst. Bot. 2015, 40, 336–351. [Google Scholar] [CrossRef]
- Spooner, D.M.; Ruess, H.; Iorizzo, M.; Senalik, D.; Simon, P. Entire plastid phylogeny of the carrot genus (Daucus, Apiaceae): Concordance with nuclear data and mitochondrial and nuclear DNA insertions to the plastid. Am. J. Bot. 2017, 104, 296–312. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.F.; Zhang, Y.X.; Guo, Z.H.; Li, D.Z. Evidence for horizontal transfer of mitochondrial DNA to the plastid genome in a bamboo genus. Sci. Rep. 2015, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wysocki, W.P.; Clark, L.G.; Attigala, L.; Ruiz-Sanchez, E.; Duvall, M.R. Evolution of the bamboos (Bambusoideae; Poaceae): A full plastome phylogenomic analysis. BMC Evol. Biol. 2015, 15, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.R. Extending the limited transfer window hypothesis to inter-organelle DNA migration. Genome Biol. Evol. 2011, 3, 743–748. [Google Scholar] [CrossRef] [PubMed]
- Cerutti, H.; Jagendorf, A. Movement of DNA across the chloroplast envelope: Implications for the transfer of promiscuous DNA. Photosynth. Res. 1995, 46, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Lloyd, A.H.; Timmis, J.N. Environmental stress increases the entry of cytoplasmic organellar DNA into the nucleus in plants. Proc. Natl. Acad. Sci. USA 2012, 109, 2444–2448. [Google Scholar] [CrossRef] [PubMed]
- Kurland, C.G.; Andersson, S.G.E. Origin and evolution of the mitochondrial proteome. Microbiol. Mol. Biol. Rev. 2000, 64, 786–820. [Google Scholar] [CrossRef] [PubMed]
- Kitazaki, K.; Kubo, T. Cost of having the largest mitochondrial genome: Evolutionary mechanism of plant mitochondrial genome. J. Bot. 2010, 2010, 620137. [Google Scholar] [CrossRef]
- Ong, H.C.; Palmer, J.D. Pervasive survival of expressed mitochondrial rps14 pseudogenes in grasses and their relatives for 80 million years following three functional transfers to the nucleus. BMC Evol. Biol. 2006, 6, 55. [Google Scholar] [CrossRef] [PubMed]
- Pamilo, P.; Viljakainen, L.; Vihavainen, A. Exceptionally high density of numts in the honeybee genome. Mol. Biol. Evol. 2007, 24, 1340–1346. [Google Scholar] [CrossRef] [PubMed]
- Timmis, J.N.; Ayliffe, M.A.; Huang, C.Y.; Martin, W. Endosymbiotic gene transfer: Organelle genomes forge eukaryotic chromosomes. Nat. Rev. Genet. 2004, 5, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, H.; Wang, J.; Sun, R.; Wu, J.; Liu, S.; Bai, Y.; Mun, J.H.; Bancroft, I.; Cheng, F.; et al. The genome of the mesopolyploid crop species Brassica rapa. Nat. Genet. 2011, 43, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, D.; Wang, X.; Ji, C.; Cheng, F. The genome sequence of allopolyploid Brassica juncea and analysis of differential homoeolog gene expression influencing selection. Nat. Genet. 2016, 48, 1225–1232. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Liu, Y.; Yang, X.; Tong, C.; Edwards, D.; Parkin, I.A.; Zhao, M.; Ma, J.; Yu, J.; Huang, S.; et al. The Brassica oleracea genome reveals the asymmetrical evolution of polyploid genomes. Nat. Commun. 2014, 5, 3930. [Google Scholar] [CrossRef] [PubMed]
- Pucker, B.; Holtgrawe, D.; Rosleff Sorensen, T.; Stracke, R.; Viehover, P.; Weisshaar, B. A de novo genome sequence assembly of the Arabidopsis thaliana accession niederzenz-1 displays presence/absence variation and strong synteny. PLoS ONE 2016, 11, e0164321. [Google Scholar] [CrossRef] [PubMed]
- Ming, R.; Hou, S.; Feng, Y.; Yu, Q.; Dionne-Laporte, A.; Saw, J.H.; Senin, P.; Wang, W.; Ly, B.V.; Lewis, K.L.; et al. The draft genome of the transgenic tropical fruit tree papaya (Carica papaya Linnaeus). Nature 2008, 452, 991–996. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.P.; Crabtree, J.; Zhao, Q.; Lorenzi, H.; Orvis, J.; Puiu, D.; Melake-Berhan, A.; Jones, K.M.; Redman, J.; Chen, G.; et al. Draft genome sequence of the oilseed species Ricinus communis. Nat. Biotechnol. 2010, 28, 951–956. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Zhang, J.; Sun, H.; Salse, J.; Lucas, W.J.; Zhang, H.; Zheng, Y.; Mao, L.; Ren, Y.; Wang, Z.; et al. The draft genome of watermelon (Citrullus lanatus) and resequencing of 20 diverse accessions. Nat. Genet. 2013, 45, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.; Thelen, J.J.; Cheng, J.; et al. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.J.; Kim, S.K.; Kim, M.Y.; Lestari, P.; Kim, K.H.; Ha, B.K.; Jun, T.H.; Hwang, W.J.; Lee, T.; Lee, J.; et al. Genome sequence of mungbean and insights into evolution within vigna species. Nat. Commun. 2014, 5, 5443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cegan, R.; Vyskot, B.; Kejnovsky, E.; Kubat, Z.; Blavet, H.; Safar, J.; Dolezel, J.; Blavet, N.; Hobza, R. Genomic diversity in two related plant species with and without sex chromosomes—Silene latifolia and S. vulgaris. PLoS ONE 2012, 7, e31898. [Google Scholar] [CrossRef] [PubMed]
- Iorizzo, M.; Ellison, S.; Senalik, D. A high-quality carrot genome assembly provides new insights into carotenoid accumulation and asterid genome evolution. Nat. Genet. 2016, 48, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Sierro, N.; Battey, J.N.; Ouadi, S.; Bakaher, N.; Bovet, L.; Willig, A.; Goepfert, S.; Peitsch, M.C.; Ivanov, N.V. The tobacco genome sequence and its comparison with those of tomato and potato. Nat. Commun. 2014, 5, 3833. [Google Scholar] [CrossRef] [PubMed]
- Jaillon, O.; Aury, J.M.; Noel, B.; Policriti, A.; Clepet, C.; Casagrande, A.; Choisne, N.; Aubourg, S.; Vitulo, N.; Jubin, C.; et al. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 2007, 449, 463–467. [Google Scholar] [PubMed]
- Wang, W.; Haberer, G.; Gundlach, H.; Glasser, C.; Nussbaumer, T.; Luo, M.C.; Lomsadze, A.; Borodovsky, M.; Kerstetter, R.A.; Shanklin, J.; et al. The Spirodela polyrhiza genome reveals insights into its neotenous reduction fast growth and aquatic lifestyle. Nat. Commun. 2014, 5, 3311. [Google Scholar] [CrossRef] [PubMed]
- Al-Mssallem, I.S.; Hu, S.; Zhang, X.; Lin, Q.; Liu, W.; Tan, J.; Yu, X.; Liu, J.; Pan, L.; Zhang, T.; et al. Genome sequence of the date palm Phoenix dactylifera L. Nat. Commun. 2013, 4, 2274. [Google Scholar] [CrossRef] [PubMed]
- Goff, S.A.; Ricke, D.; Lan, T.H.; Presting, G.; Wang, R.; Dunn, M.; Glazebrook, J.; Sessions, A.; Oeller, P.; Varma, H.; et al. A draft sequence of the rice genome (Oryza sativa L. ssp. japonica). Science 2002, 296, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, L.L.; Xing, F.; Kudrna, D.A.; Yao, W.; Copetti, D.; Mu, T.; Li, W.; Song, J.M.; Xie, W. Extensive sequence divergence between the reference genomes of two elite indica rice varieties zhenshan 97 and minghui 63. Proc. Natl. Acad. Sci. USA 2016, 113, E5163–E5171. [Google Scholar] [CrossRef] [PubMed]
- Paterson, A.H.; Bowers, J.E.; Bruggmann, R.; Dubchak, I.; Grimwood, J.; Gundlach, H.; Haberer, G.; Hellsten, U.; Mitros, T.; Poliakov, A.; et al. The Sorghum bicolor genome and the diversification of grasses. Nature 2009, 457, 551–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnable, P.S.; Ware, D.; Fulton, R.S.; Stein, J.C.; Wei, F.; Pasternak, S.; Liang, C.; Zhang, J.; Fulton, L.; Graves, T.A.; et al. The B73 maize genome: Complexity, diversity and dynamics. Science 2009, 326, 1112–1115. [Google Scholar] [CrossRef] [PubMed]
- Project, A.G. The Amborella genome and the evolution of flowering plants. Science 2013, 342, 1241089. [Google Scholar] [CrossRef] [PubMed]
- Izuno, A.; Hatakeyama, M.; Nishiyama, T.; Tamaki, I.; Shimizu-Inatsugi, R.; Sasaki, R.; Shimizu, K.K.; Isagi, Y. Genome sequencing of Metrosideros polymorpha (Myrtaceae), a dominant species in various habitats in the hawaiian islands with remarkable phenotypic variations. J. Plant Res. 2016, 129, 727–736. [Google Scholar] [CrossRef] [PubMed]
- Rensing, S.A.; Lang, D.; Zimmer, A.D.; Terry, A.; Salamov, A.; Shapiro, H.; Nishiyama, T.; Perroud, P.F.; Lindquist, E.A.; Kamisugi, Y.; et al. The Physcomitrella genome reveals evolutionary insights into the conquest of land by plants. Science 2008, 319, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Qiu, F.; Ungerer, M.C. Genomic abundance and transcriptional activity of diverse gypsy and copia long terminal repeat retrotransposons in three wild sunflower species. BMC Plant Biol. 2018, 18, 6. [Google Scholar] [CrossRef] [PubMed]
- Goremykin, V.V.; Lockhart, P.J.; Viola, R.; Velasco, R. The mitochondrial genome of Malus domestica and the import-driven hypothesis of mitochondrial genome expansion in seed plants. Plant J. 2012, 71, 615–626. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ruhlman, T.A.; Sabir, J.; Blazier, J.C.; Jansen, R.K. Coordinated rates of evolution between interacting plastid and nuclear genes in Geraniaceae. Plant Cell 2015, 27, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, C.; Kobayashi, Y.; Aoki, S.; Sugita, C.; Sugita, M. Complete chloroplast DNA sequence of the moss Physcomitrella patens: Evidence for the loss and relocation of rpoa from the chloroplast to the nucleus. Nucleic Acids Res. 2003, 31, 5324–5331. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Hazkani-Covo, E.; Covo, S. Numt-mediated double-strand break repair mitigates deletions during primate genome evolution. PLoS Genet. 2008, 4, e1000237. [Google Scholar] [CrossRef] [PubMed]
- Ju, Y.S.; Tubio, J.M.C.; Mifsud, W.; Fu, B.Y.; Davies, H.R.; Ramakrishna, M.; Li, Y.L.; Yates, L.; Gundem, G.; Tarpey, P.S.; et al. Frequent somatic transfer of mitochondrial DNA into the nuclear genome of human cancer cells. Genome Res. 2015, 25, 814–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. Mega5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.W.; Feng, K.; Grover, C.E.; Li, P.B.; Liu, F.; Wang, Y.M.; Xu, Q.; Shang, M.Z.; Zhou, Z.L.; Cai, X.Y.; et al. Chloroplast DNA structural variation, phylogeny and age of divergence among diploid cotton species. PLoS ONE 2016, 11, e0157183. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.W.; Grover, C.E.; Li, P.B.; Wang, Y.M.; Nie, H.S.; Zhao, Y.P.; Wang, M.Y.; Liu, F.; Zhou, Z.L.; Wang, X.X.; et al. Molecular evolution of the plastid genome during diversification of the cotton genus. Mol. Phylogenet. Evol. 2017, 112, 268–276. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Repeat Sizes (Mb) | Genome Sizes (Mb) | Repeat/Genome (%) | References | |
|---|---|---|---|---|---|
| Spermatophytes | Eudicots | ||||
| B. rapa | 191.63 | 284.13 | 67.44 | [50] | |
| B. napus | 441.77 | 930.51 | 47.48 | [51] | |
| B. oleracea | 185.43 | 539.91 | 34.34 | [52] | |
| A. thaliana 1 | 23.58 | 119.67 | 19.70 | [53] | |
| C. papaya | 316.53 | 369.78 | 85.60 | [54] | |
| R. communis | 176.00 | 350.62 | 50.20 | [55] | |
| C. lanatus | 159.80 | 321.05 | 49.77 | [56] | |
| G. max | 587.10 | 978.97 | 59.97 | [57] | |
| V. radiata | 216.17 | 548.08 | 39.44 | [58] | |
| S. latifolia | 244.82 | 665.28 | 36.80 | [59] | |
| D. carota | 193.70 | 473.00 | 40.95 | [60] | |
| N. tabacum | 3479.49 | 4500.00 | 77.32 | [61] | |
| V. vinifera | 185.35 | 487.00 | 38.06 | [62] | |
| Monocots | |||||
| S. polyrhiza 1 | 19.43 | 132.01 | 14.72 | [63] | |
| P. dactylifera | 214.34 | 558.02 | 38.41 | [64] | |
| O. sativa japonica | 188.00 | 374.42 | 50.21 | [65] | |
| O. sativa indica | 148.14 | 374.25 | 39.58 | [66] | |
| S. bicolor | 231.28 | 739.15 | 31.29 | [67] | |
| Z. mays | 1757.48 | 2067.62 | 85.00 | [68] | |
| Basal Angiosperms | |||||
| A. trichopoda | 407.43 | 706.50 | 57.67 | [69] | |
| Bryophytes | M. polymorpha 1 | 12.48 | 304.37 | 4.10 | [70] |
| P. patens 1 | 79.37 | 477.95 | 16.61 | [71] | |
| Species | Mitochondrial Genome | |||
|---|---|---|---|---|
| Repeat Size (Kb) | Repeat Number (>1 kb) | Repeat Number (>100 bp) | ||
| Spermatophytes | Eudicots | |||
| B. rapa | 3.80 | 1 | 9 | |
| B. napus | 4.62 | 1 | 17 | |
| B. oleracea | 152.00 | 2 | 24 | |
| A. thaliana | 15.63 | 2 | 25 | |
| C. papaya | 13.43 | 1 | 13 | |
| R. communis | 5.80 | 6 | 6 | |
| G. max | 60.67 | 13 | 68 | |
| V. radiata | 1.02 | 0 | 6 | |
| S. latifolia | 23.27 | 15 | 17 | |
| D. carota | 71.09 | 4 | 19 | |
| N. tabacum | 42.07 | 3 | 22 | |
| V. vinifera | 5.77 | 0 | 26 | |
| Monocots | ||||
| S. polyrhiza | 1.58 | 0 | 5 | |
| P. dactylifera | 3.03 | 1 | 12 | |
| O. sativa japonica | 141.19 | 12 | 39 | |
| O. sativa indica | 141.76 | 11 | 27 | |
| S. bicolor | 58.56 | 5 | 18 | |
| Z. mays | 51.94 | 4 | 19 | |
| Basal Angiosperms | ||||
| A. trichopoda | 266.14 | 1 | 1811 | |
| Gymnosperms | ||||
| C. taitungensis | 62.65 | 2 | 5070 | |
| Bryophytes | M. polymorpha | 2.08 | 0 | 13 |
| P. patens | 0 | 0 | 0 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, N.; Wang, Y.; Hua, J. The Roles of Mitochondrion in Intergenomic Gene Transfer in Plants: A Source and a Pool. Int. J. Mol. Sci. 2018, 19, 547. https://doi.org/10.3390/ijms19020547
Zhao N, Wang Y, Hua J. The Roles of Mitochondrion in Intergenomic Gene Transfer in Plants: A Source and a Pool. International Journal of Molecular Sciences. 2018; 19(2):547. https://doi.org/10.3390/ijms19020547
Chicago/Turabian StyleZhao, Nan, Yumei Wang, and Jinping Hua. 2018. "The Roles of Mitochondrion in Intergenomic Gene Transfer in Plants: A Source and a Pool" International Journal of Molecular Sciences 19, no. 2: 547. https://doi.org/10.3390/ijms19020547
APA StyleZhao, N., Wang, Y., & Hua, J. (2018). The Roles of Mitochondrion in Intergenomic Gene Transfer in Plants: A Source and a Pool. International Journal of Molecular Sciences, 19(2), 547. https://doi.org/10.3390/ijms19020547