Bax Inhibitor-1 Acts as an Anti-Influenza Factor by Inhibiting ROS Mediated Cell Death and Augmenting Heme-Oxygenase 1 Expression in Influenza Virus Infected Cells


 ,
,  , ,
, , 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
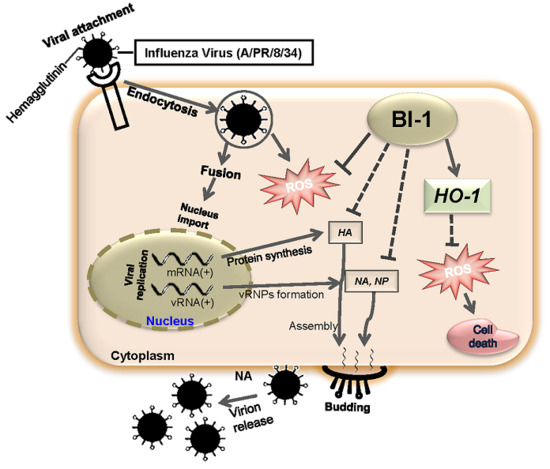
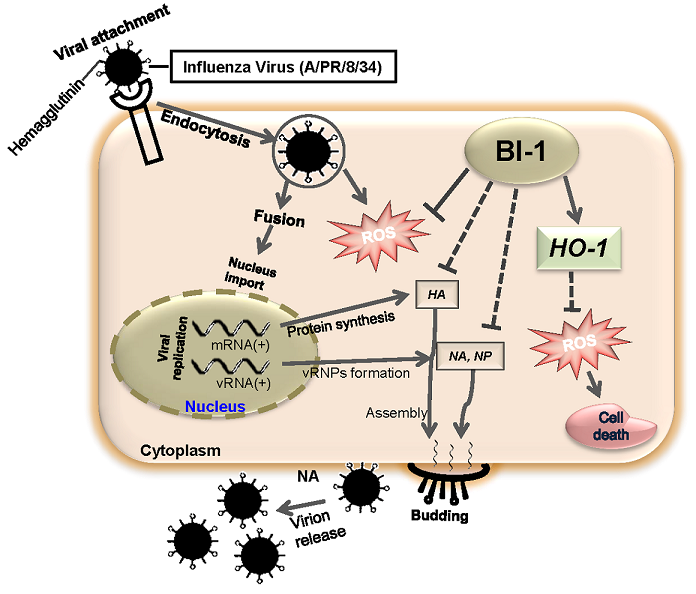
2. Results and Discussion
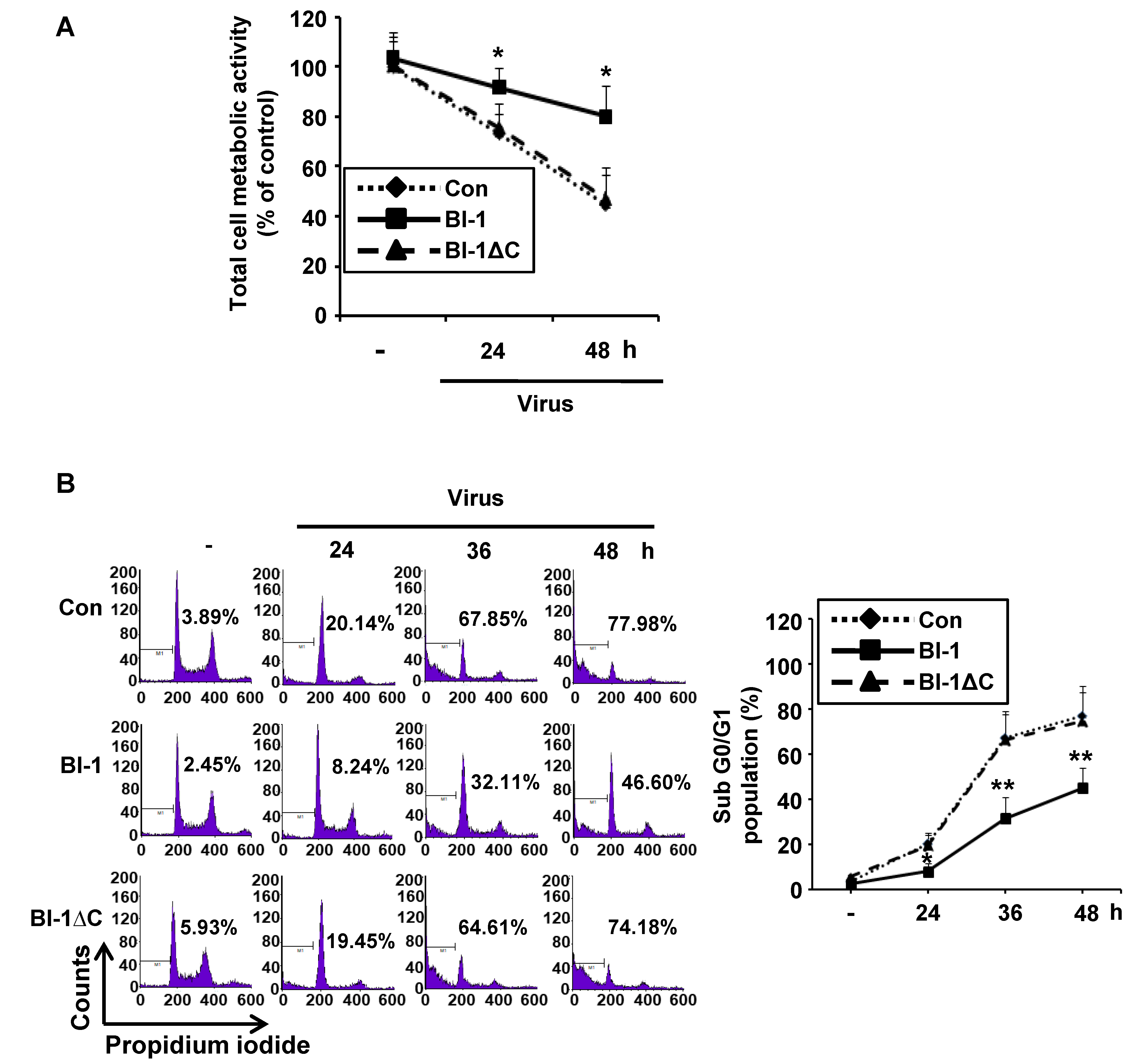
2.1. Overexpression of Bax Inhibitor-1 (BI-1) Relieves Virus Induced Cell Death in Madin-Darby Canine Kidney (MDCK) Cells
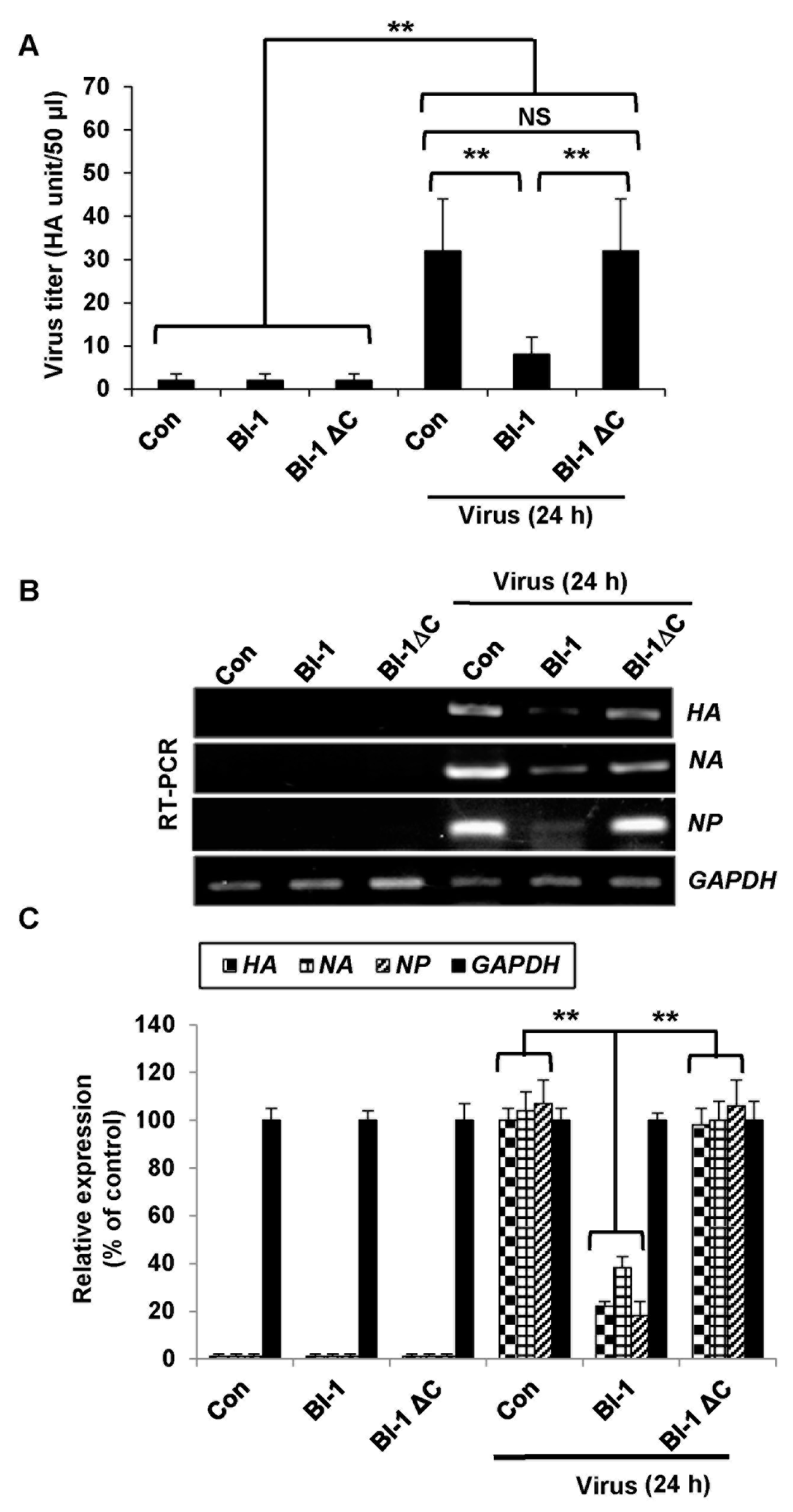
2.2. Overexpression of BI-1 Inhibits Viral Replication and Viral Gene Expression in MDCK Cells
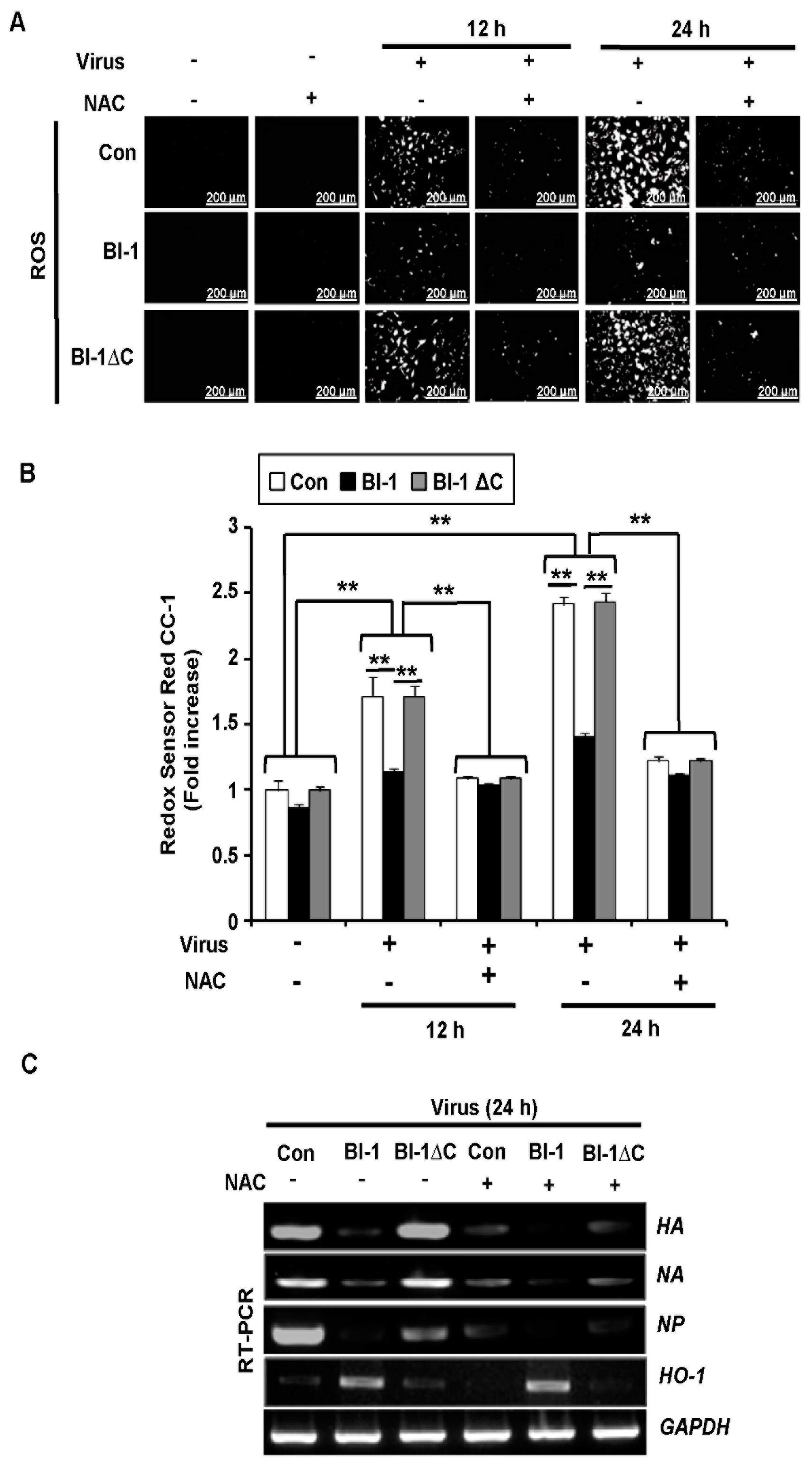
2.3. BI-1 Overexpression Suppresses ROS-Mediated Influenza Virus Infection in MDCK Cells
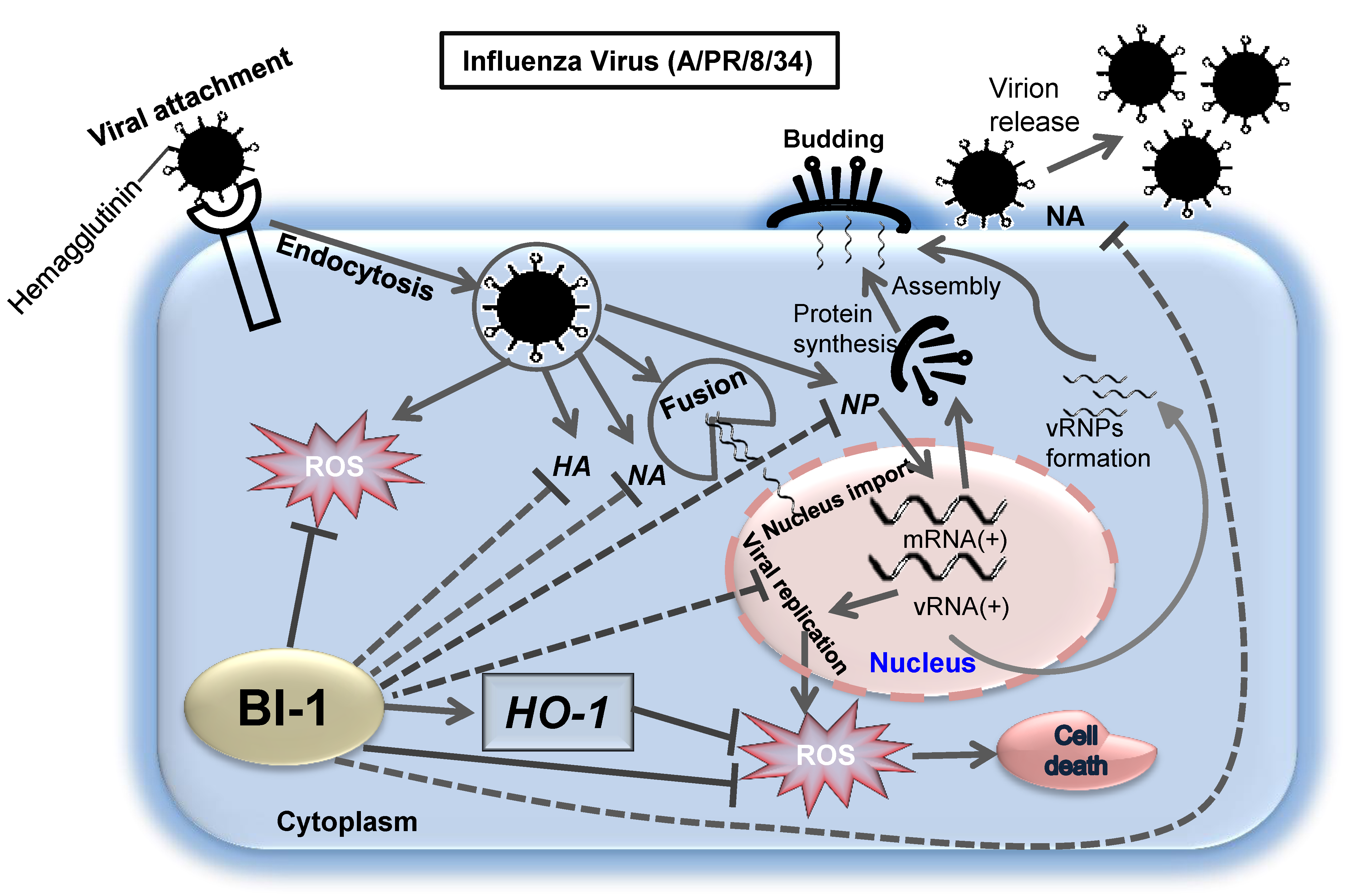
2.4. BI-1 May Independently Act as an Antiviral Protein through the Suppression of ROS Production by Activating the Expression of HO-1 with Additional Effect of the ROS Scavenging Agent NAC
3. Materials and Methods
3.1. Cells, Virus, Reagents, and Lentiviral Infection
3.2. Cell Viability and Cell Cycle Analysis
3.3. Virus Yield Reduction Assay
3.4. Intracellular ROS Detection
3.5. RT-PCR
3.6. Western Blot Analysis
3.7. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| BI-1 | Bax inhibitor-1 |
| ERK | Extracellular Signal–regulated kinase |
| STAT | Signal transducer and activator of transcription |
| NAC | N-acetyl cysteine |
| ROS | Reactive oxygen species |
| HO-1 | Heme oxygenase-1 |
| HA | Hemagglutinin |
| NA | Neuraminidase |
| M2 | Matrix-2 |
| MDCK | Madin Darby Canine Kidney |
| cPPT | Central polypurine tract |
| TCID50 | Tissue culture infective dose 50 |
| PBS | Phosphate buffered saline |
| IgG | Immunoglobulin G |
| ECL | Enhanced chemiluminescence |
| FBS | Fetal bovine serum |
| GFP | Green fluorescent protein |
| WPRE | Woodchuck hepatitis virus post-transcriptional regulatory element |
References
- Palese, P. Influenza: Old and new threats. Nat. Med. 2004, 10, S82–S87. [Google Scholar] [CrossRef] [PubMed]
- Neumann, G.; Fujii, K.; Kino, Y.; Kawaoka, Y. An improved reverse genetics system for influenza A virus generation and its implications for vaccine production. Proc. Natl. Acad. Sci. USA 2005, 102, 16825–16829. [Google Scholar] [CrossRef] [PubMed]
- Hayden, F.G. Respiratory viral threats. Curr. Opin. Infect. Dis. 2006, 19, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Meijer, A.; Rebelo-de-Andrade, H.; Correia, V.; Besselaar, T.; Drager-Dayal, R.; Fry, A.; Gregory, V.; Gubareva, L.; Kageyama, T.; Lackenby, A.; et al. Global update on the susceptibility of human influenza viruses to neuraminidase inhibitors, 2012–2013. Antivir. Res. 2014, 110, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Hurt, A.C.; Besselaar, T.G.; Daniels, R.S.; Ermetal, B.; Fry, A.; Gubareva, L.; Huang, W.; Lackenby, A.; Lee, R.T.; Lo, J.; et al. Global update on the susceptibility of human influenza viruses to neuraminidase inhibitors, 2014–2015. Antivir. Res. 2016, 132, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Bouvier, N.M.; Lowen, A.C.; Palese, P. Oseltamivir-resistant influenza A viruses are transmitted efficiently among guinea pigs by direct contact but not by aerosol. J. Virol. 2008, 82, 10052–10058. [Google Scholar] [CrossRef] [PubMed]
- Söderholm, S.; Kainov, D.E.; Öhman, T.; Denisova, O.V.; Schepens, B.; Kulesskiy, E.; Imanishi, S.Y.; Corthals, G.; Hintsanen, P.; Aittokallio, T. Phosphoproteomics to characterize host response during influenza A virus infection of human macrophages. Mol. Cell. Proteom. 2016, 15, 3203–3219. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Jia, W.; Zhao, A. Anti-influenza agents from plants and traditional Chinese medicine. Phytother. Res. 2006, 20, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J.; Song, J.H.; Park, K.S.; Kwon, D.H. Inhibitory effects of quercetin 3-rhamnoside on influenza A virus replication. Eur. J. Pharm. Sci. 2009, 37, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Vlahos, R.; Stambas, J.; Selemidis, S. Suppressing production of reactive oxygen species (ROS) for influenza A virus therapy. Trends Pharmacol. Sci. 2012, 33, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.; Lowther, S.; Stambas, J. Inhibition of reactive oxygen species production ameliorates inflammation induced by influenza A viruses via upregulation of SOCS1 and SOCS3. J. Virol. 2015, 89, 2672–2683. [Google Scholar] [CrossRef] [PubMed]
- Gangehei, L.; Ali, M.; Zhang, W.; Chen, Z.; Wakame, K.; Haidari, M. Oligonol a low molecular weight polyphenol of lychee fruit extract inhibits proliferation of influenza virus by blocking reactive oxygen species-dependent ERK phosphorylation. Phytomedicine 2010, 17, 1047–1056. [Google Scholar] [CrossRef] [PubMed]
- Uchide, N.; Ohyama, K. Antiviral function of pyrrolidine dithiocarbamate against influenza virus: The inhibition of viral gene replication and transcription. J. Antimicrob. Chemother. 2003, 52, 8–10. [Google Scholar] [CrossRef] [PubMed]
- Baek, D.; Nam, J.; Koo, Y.D.; Kim, D.H.; Lee, J.; Jeong, J.C.; Kwak, S.-S.; Chung, W.S.; Lim, C.O.; Bahk, J.D. Bax-induced cell death of Arabidopsisis meditated through reactive oxygen-dependent and-independent processes. Plant Mol. Biol. 2004, 56, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, P.M.; Ryu, S.-Y.; Bombrun, A.; Antonsson, B.; Kinnally, K.W. MAC inhibitors suppress mitochondrial apoptosis. Biochem. J. 2009, 423, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Bombrun, A.; Gerber, P.; Casi, G.; Terradillos, O.; Antonsson, B.; Halazy, S. 3,6-dibromocarbazole piperazine derivatives of 2-propanol as first inhibitors of cytochrome c release via Bax channel modulation. J. Med. Chem. 2003, 46, 4365–4368. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Brahmbhatt, H.; Mergenthaler, P.; Zhang, Z.; Sang, J.; Daude, M.; Ehlert, F.G.; Diederich, W.E.; Wong, E.; Zhu, W. A small-molecule inhibitor of Bax and Bak oligomerization prevents genotoxic cell death and promotes neuroprotection. Cell Chem. Biol. 2017, 24, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, P.M.; Teijido, O.; Mirzalieva, O.; Dejean, L.M.; Pavlov, E.V.; Antonsson, B.; Kinnally, K.W. MAC inhibitors antagonize the pro-apoptotic effects of tBid and disassemble Bax/Bak oligomers. J. Bioenerg. Biomembr. 2017, 49, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, E.R.; Jeon, K.; Choi, H.Y.; Lim, H.; Kim, S.J.; Chae, H.J.; Park, S.H.; Kim, S.; Seo, Y.R.; et al. Role of BI-1 (TEGT)-mediated ERK1/2 activation in mitochondria-mediated apoptosis and splenomegaly in BI-1 transgenic mice. Biochim. Biophys. Acta Mol. Cell Res. 1823, 1823, 876–888. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, H.; Nirasawa, S.; Kiba, A.; Urasaki, N.; Saitoh, H.; Ito, M.; Kawai-Yamada, M.; Uchimiya, H.; Terauchi, R. Overexpression of Bax inhibitor suppresses the fungal elicitor-induced cell death in rice (Oryza sativa L.) cells. Plant J. 2003, 33, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Li, Y.; Li, H.; Zhang, Y.; Xu, J.; Ren, L.; Fu, S.; Zhou, Y. Bax inhibitor-1 is overexpressed in non-small cell lung cancer and promotes its progression and metastasis. Int. J. Clin. Exp. Pathol. 2015, 8, 1411. [Google Scholar] [PubMed]
- Lee, G.-H.; Kim, H.-K.; Chae, S.-W.; Kim, D.-S.; Ha, K.-C.; Cuddy, M.; Kress, C.; Reed, J.C.; Kim, H.-R.; Chae, H.-J. Bax inhibitor-1 regulates endoplasmic reticulum stress-associated reactive oxygen species and heme oxygenase-1 expression. J. Biol. Chem. 2007, 282, 21618–21628. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.R.; Lee, G.H.; Lee, H.Y.; Kim, D.S.; Chung, M.J.; Lee, Y.C.; Kim, H.R.; Chae, H.J. BAX inhibitor-1-associated V-ATPase glycosylation enhances collagen degradation in pulmonary fibrosis. Cell Death Dis. 2014, 5, e1113. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Yadav, R.K.; Jeong, G.S.; Kim, H.R.; Chae, H.J. The characteristics of Bax inhibitor-1 and its related diseases. Curr. Mol. Med. 2014, 14, 603–615. [Google Scholar] [CrossRef] [PubMed]
- Robinson, K.S.; Clements, A.; Williams, A.C.; Berger, C.N.; Frankel, G. Bax inhibitor 1 in apoptosis and disease. Oncogene 2011, 30, 2391–2400. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Zhang, A.; Zhang, C.; Ni, H.; Gao, J.; Wang, C.; Zhao, Q.; Wang, X.; Wang, X.; Ma, C. Heme oxygenase-1 acts as an antiviral factor for porcine reproductive and respiratory syndrome virus infection and over-expression inhibits virus replication in vitro. Antivir. Res. 2014, 110, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, J.A.; González, P.A.; Kalergis, A.M. Modulation of antiviral immunity by heme oxygenase-1. Am. J. Pathol. 2017, 187, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Kawai-Yamada, M.; Ohori, Y.; Uchimiya, H. Dissection of Arabidopsis Bax inhibitor-1 suppressing Bax-, hydrogen peroxide-, and salicylic acid-induced cell death. Plant Cell 2004, 16, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Jeon, K.; Lim, H.; Kim, J.H.; Han, D.; Lee, E.R.; Yang, G.M.; Song, M.K.; Cho, S.G. Bax inhibitor-1 enhances survival and neuronal differentiation of embryonic stem cells via differential regulation of mitogen-activated protein kinases activities. Biochim. Biophys. Acta Mol. Cell Res. 2012, 1823, 2190–2200. [Google Scholar] [CrossRef] [PubMed]
- Sawai, R.; Kuroda, K.; Shibata, T.; Gomyou, R.; Osawa, K.; Shimizu, K. Anti-influenza virus activity of Chaenomeles sinensis. J. Ethnopharmacol. 2008, 118, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.K.; Choi, H.Y.; Hwang, J.S.; Dayem, A.A.; Kim, J.H.; Kim, Y.B.; Poo, H.; Cho, S.G. Antiviral activity of 3,4′-dihydroxyflavone on influenza a virus. J. Microbiol. 2014, 52, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Dayem, A.A.; Choi, H.Y.; Kim, Y.B.; Cho, S.G. Antiviral Effect of Methylated Flavonol Isorhamnetin against Influenza. PLoS ONE 2015, 10, e0121610. [Google Scholar] [CrossRef] [PubMed]
- Geiler, J.; Michaelis, M.; Naczk, P.; Leutz, A.; Langer, K.; Doerr, H.-W.; Cinatl, J. N-acetyl-l-cysteine (NAC) inhibits virus replication and expression of pro-inflammatory molecules in A549 cells infected with highly pathogenic H5N1 influenza A virus. Biochem. Pharmacol. 2010, 79, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-H.; Song, M.; Kang, G.-H.; Lee, E.-R.; Choi, H.-Y.; Lee, C.; Kim, J.-H.; Kim, Y.; Koo, B.-N.; Cho, S.-G. Combined treatment of 3-hydroxyflavone and imatinib mesylate increases apoptotic cell death of imatinib mesylate-resistant leukemia cells. Leuk. Res. 2012, 36, 1157–1164. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-R.; Lee, G.-H.; Cho, E.Y.; Chae, S.-W.; Ahn, T.; Chae, H.-J. Bax inhibitor 1 regulates ER-stress-induced ROS accumulation through the regulation of cytochrome P450 2E1. J. Cell Sci. 2009, 122, 1126–1133. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-C.; Kappes, M.A.; Chen, M.-C.; Lin, C.-C.; Wang, T.T. Distinct susceptibility and applicability of MDCK derivatives for influenza virus research. PLoS ONE 2017, 12, e0172299. [Google Scholar] [CrossRef] [PubMed]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hossain, M.K.; Saha, S.K.; Abdal Dayem, A.; Kim, J.-H.; Kim, K.; Yang, G.-M.; Choi, H.Y.; Cho, S.-G. Bax Inhibitor-1 Acts as an Anti-Influenza Factor by Inhibiting ROS Mediated Cell Death and Augmenting Heme-Oxygenase 1 Expression in Influenza Virus Infected Cells. Int. J. Mol. Sci. 2018, 19, 712. https://doi.org/10.3390/ijms19030712
Hossain MK, Saha SK, Abdal Dayem A, Kim J-H, Kim K, Yang G-M, Choi HY, Cho S-G. Bax Inhibitor-1 Acts as an Anti-Influenza Factor by Inhibiting ROS Mediated Cell Death and Augmenting Heme-Oxygenase 1 Expression in Influenza Virus Infected Cells. International Journal of Molecular Sciences. 2018; 19(3):712. https://doi.org/10.3390/ijms19030712
Chicago/Turabian StyleHossain, Mohammed Kawser, Subbroto Kumar Saha, Ahmed Abdal Dayem, Jung-Hyun Kim, Kyeongseok Kim, Gwang-Mo Yang, Hye Yeon Choi, and Ssang-Goo Cho. 2018. "Bax Inhibitor-1 Acts as an Anti-Influenza Factor by Inhibiting ROS Mediated Cell Death and Augmenting Heme-Oxygenase 1 Expression in Influenza Virus Infected Cells" International Journal of Molecular Sciences 19, no. 3: 712. https://doi.org/10.3390/ijms19030712
APA StyleHossain, M. K., Saha, S. K., Abdal Dayem, A., Kim, J. -H., Kim, K., Yang, G. -M., Choi, H. Y., & Cho, S. -G. (2018). Bax Inhibitor-1 Acts as an Anti-Influenza Factor by Inhibiting ROS Mediated Cell Death and Augmenting Heme-Oxygenase 1 Expression in Influenza Virus Infected Cells. International Journal of Molecular Sciences, 19(3), 712. https://doi.org/10.3390/ijms19030712