Gold Nanoparticle-Induced Cell Death and Potential Applications in Nanomedicine



Abstract
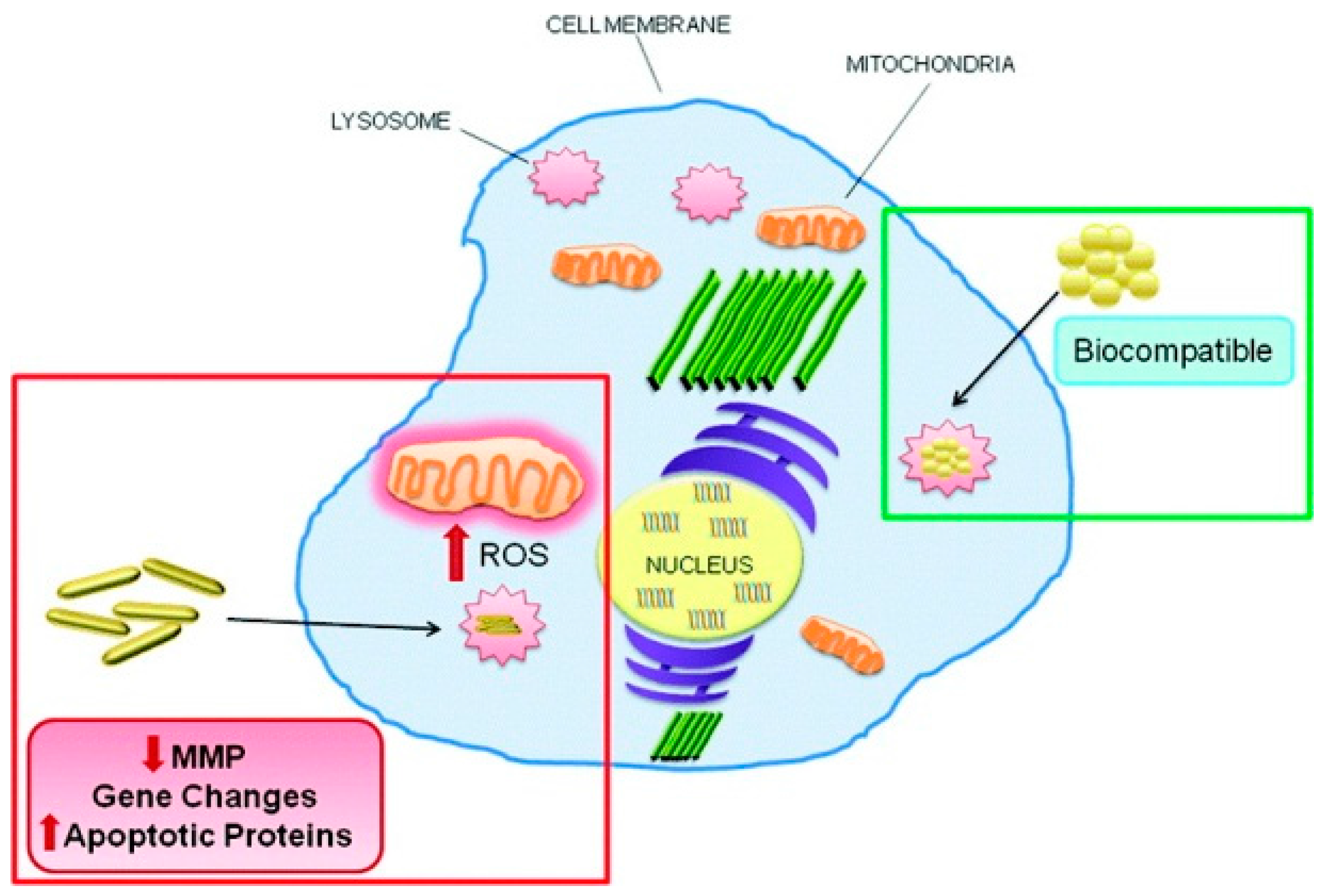
:1. Introduction
2. Determination of Cell Proliferation by GNPs
3. GNP-Induced Apoptosis
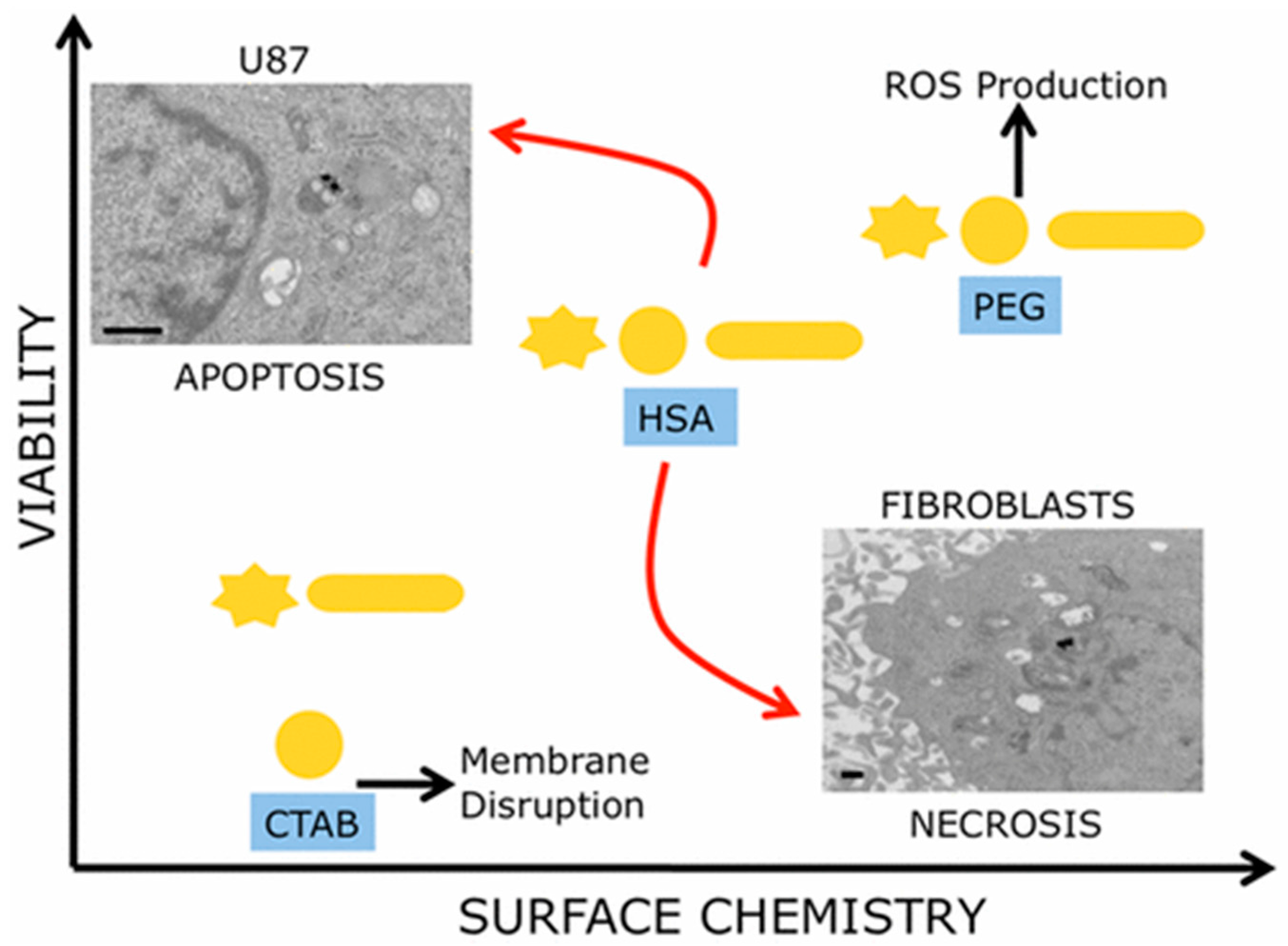
4. GNP-Induced Necrosis
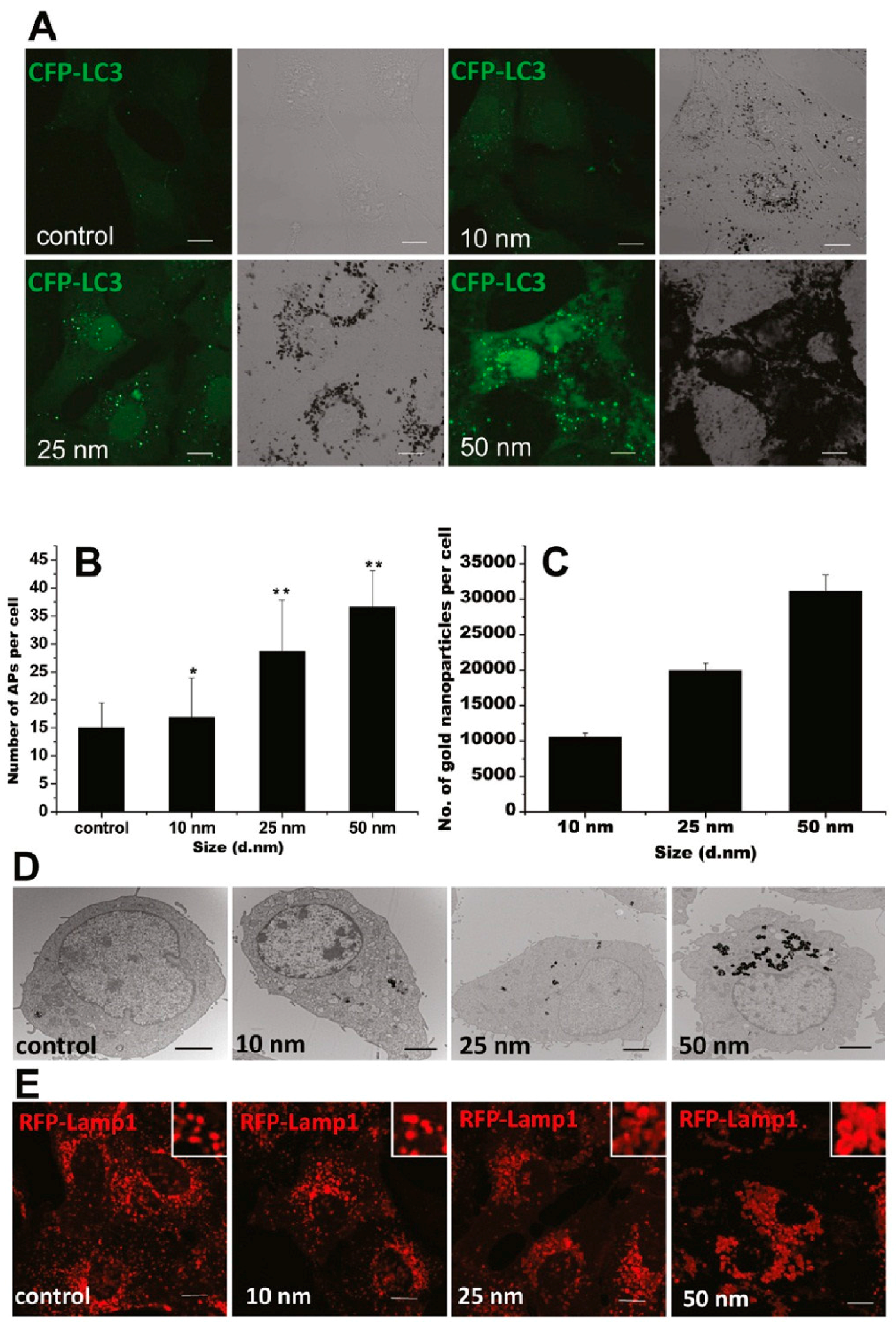
5. GNP-Induced Autophagy
6. Conclusions and Perspectives for Future Work
Acknowledgments
Conflicts of Interest
References
- Peter, M.E. Apoptosis meets necrosis. Nature 2011, 471, 310–312. [Google Scholar] [CrossRef] [PubMed]
- Levine, B.; Yuan, J. Autophagy in cell death: An innocent convict? J. Clin. Investig. 2005, 115, 2679–2688. [Google Scholar] [CrossRef] [PubMed]
- Vandenabeele, P.; Galluzzi, L.; Vanden Berghe, T.; Kroemer, G. Molecular mechanisms of necroptosis: An ordered cellular explosion. Nat. Rev. Mol. Cell. Biol. 2010, 11, 700–714. [Google Scholar] [CrossRef] [PubMed]
- Formigli, L.; Papucci, L.; Tani, A.; Schiavone, N.; Tempestini, A.; Orlandini, G.E.; Capaccioli, S.; Orlandini, S.Z. Aponecrosis: Morphological and biochemical exploration of a syncretic process of cell death sharing apoptosis and necrosis. J. Cell. Physiol. 2000, 182, 41–49. [Google Scholar] [CrossRef]
- Bergsbaken, T.; Fink, S.L.; Cookson, B.T. Pyroptosis: Host cell death and inflammation. Nat. Rev. Microbiol. 2009, 7, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Kerr, J.F.R.; Wyllie, A.H.; Currie, A.R. Apoptosis: A basic biological phenomenon with wide-ranging implications in tissue kinetics. Br. J. Cancer 1972, 26, 239–257. [Google Scholar] [CrossRef] [PubMed]
- Wyllie, A.H.; Kerr, J.F.R.; Currie, A.R. Cell death: The significance of apoptosis. In International Review of Cytology; Bourne, G.H., Danielli, J.F., Jeon, K.W., Eds.; Academic Press: New York NY, USA, 1980; Volume 68, pp. 251–306. [Google Scholar]
- Edinger, A.L.; Thompson, C.B. Death by design: Apoptosis, necrosis and autophagy. Curr. Opin. Cell Biol. 2004, 16, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Susan, E. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar]
- Thompson, C.B. Apoptosis in the pathogenesis and treatment of disease. Science 1995, 267, 1456–1462. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef]
- Yuan, J.; Yankner, B.A. Apoptosis in the nervous system. Nature 2000, 407, 802–809. [Google Scholar] [CrossRef] [PubMed]
- Green, D.R.; Evan, G.I. A matter of life and death. Cancer Cell 2002, 1, 19–30. [Google Scholar] [CrossRef]
- Vaux, D.L.; Flavell, R.A. Apoptosis genes and autoimmunity. Curr. Opin. Immunol. 2000, 12, 719–724. [Google Scholar] [CrossRef]
- Vellai, T. Autophagy genes and aging. Cell Death Differ. 2009, 16, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Ost, A.; Svensson, K.; Ruishalme, I.; Brännmark, C.; Franck, N.; Krook, H.; Sandström, P.; Kjolhede, P.; Strålfors, P. Attenuated mTOR signaling and enhanced autophagy in adipocytes from obese patients with type 2 diabetes. Mol. Med. 2010, 16, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.; Sluimer, J.; Wang, Y.; Subramanian, M.; Brown, K.; Pattison, J.S.; Robbins, J.; Martinez, J.; Tabas, I. Macrophage autophagy plays a protective role in advanced atherosclerosis. Cell Metab. 2012, 15, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, N.; Levine, B.; Cuervo, A.M.; Klionsky, D.J. Autophagy fights disease through cellular self-digestion. Nature 2008, 451, 1069–1075. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.E.; Zuckerman, J.E.; Choi, C.H.J.; Seligson, D.; Tolcher, A.; Alabi, C.A.; Yen, Y.; Heidel, J.D.; Ribas, A. Evidence of RNAi in humans from systemically administered siRNA via targeted nanoparticles. Nature 2010, 464, 1067–1070. [Google Scholar] [CrossRef] [PubMed]
- Mura, S.; Nicolas, J.; Couvreur, P. Stimuli-responsive nanocarriers for drug delivery. Nat. Mater. 2013, 12, 991–1003. [Google Scholar] [CrossRef] [PubMed]
- Boisselier, E.; Astruc, D. Gold nanoparticles in nanomedicine: Preparations, imaging, diagnostics, therapies and toxicity. Chem. Soc. Rev. 2009, 38, 1759–1782. [Google Scholar] [CrossRef] [PubMed]
- Dreaden, E.C.; Alkilany, A.M.; Huang, X.; Murphy, C.J.; El-Sayed, M.A. The golden age: Gold nanoparticles for biomedicine. Chem. Soc. Rev. 2012, 41, 2740–2779. [Google Scholar] [CrossRef] [PubMed]
- Duncan, R. Polymer conjugates as anticancer nanomedicines. Nat. Rev. Cancer 2006, 6, 688–701. [Google Scholar] [CrossRef] [PubMed]
- Giljohann, D.A.; Seferos, D.S.; Daniel, W.L.; Massich, M.D.; Patel, P.C.; Mirkin, C.A. Gold nanoparticles for biology and medicine. Angew. Chem. Int. Ed. 2010, 49, 3280–3294. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, I.H.; Huang, X.H.; El-Sayed, M.A. Surface plasmon resonance scattering and absorption of anti-EGFR antibody conjugated gold nanoparticles in cancer diagnostics: Applications in oral cancer. Nano Lett. 2005, 5, 829–834. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, P.; Han, G.; De, M.; Kim, C.K.; Rotello, V.M. Gold nanoparticles in delivery applications. Adv. Drug Deliv. Rev. 2008, 60, 1307–1315. [Google Scholar] [CrossRef] [PubMed]
- Chithrani, D.B.; Jelveh, S.; Jalali, F.; van Prooijen, M.; Allen, C.; Bristow, R.G.; Hill, R.P.; Jaffray, D.A. Gold nanoparticles as radiation sensitizers in cancer therapy. Radiat. Res. 2010, 173, 719–728. [Google Scholar] [CrossRef] [PubMed]
- Hainfeld, J.F.; Dilmanian, F.A.; Slatkin, D.N.; Smilowitz, H.M. Radiotherapy enhancement with gold nanoparticles. J. Pharm. Pharmacol. 2008, 60, 977–985. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Jain, P.K.; El-Sayed, I.H.; El-Sayed, M.A. Plasmonic photothermal therapy (PPTT) using gold nanoparticles. Lasers Med. Sci. 2008, 23, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.H.; El-Sayed, I.H.; Qian, W.; El-Sayed, M.A. Cancer cell imaging and photothermal therapy in the near-infrared region by using gold nanorods. J. Am. Chem. Soc. 2006, 128, 2115–2120. [Google Scholar] [CrossRef] [PubMed]
- Chithrani, B.D.; Ghazani, A.A.; Chan, W.C.W. Determining the size and shape dependence of gold nanoparticle uptake into mammalian cells. Nano Lett. 2006, 6, 662–668. [Google Scholar] [CrossRef] [PubMed]
- Walkey, C.D.; Olsen, J.B.; Guo, H.; Emili, A.; Chan, W.C.W. Nanoparticle size and surface chemistry determine serum protein adsorption and macrophage uptake. J. Am. Chem. Soc. 2012, 134, 2139–2147. [Google Scholar] [CrossRef] [PubMed]
- Su, G.; Zhou, H.; Mu, Q.; Zhang, Y.; Li, L.; Jiao, P.; Jiang, G.; Yan, B. Effective surface charge density determines the electrostatic attraction between nanoparticles and cells. J. Phys. Chem. C 2012, 116, 4993–4998. [Google Scholar] [CrossRef]
- Li, S.; Zhai, S.; Liu, Y.; Zhou, H.; Wu, J.; Jiao, Q.; Zhang, B.; Zhu, H.; Yan, B. Experimental modulation and computational model of nano-hydrophobicity. Biomaterials 2015, 52, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Yen, H.-J.; Hsu, S.-h.; Tsai, C.-L. Cytotoxicity and immunological response of gold and silver nanoparticles of different sizes. Small 2009, 5, 1553–1561. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.-W.; Liaw, J.-W.; Kao, Y.-C.; Huang, M.-Y.; Lee, C.-Y.; Rau, L.-R.; Huang, C.-Y.; Wei, K.-C.; Ye, T.-C. Internalized gold nanoparticles do not affect the osteogenesis and apoptosis of MG63 osteoblast-like cells: A quantitative, in vitro study. PLoS ONE 2013, 8, e76545. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Li, Z.Q.; Wang, Y.; Liu, H.W. Gold nanoparticles promote proliferation of human periodontal ligament stem cells and have limited effects on cells differentiation. J. Nanomater. 2016, 2016, 1431836. [Google Scholar] [CrossRef]
- Grabinski, C.; Schaeublin, N.; Wijaya, A.; D’Couto, H.; Baxamusa, S.H.; Hamad-Schifferli, K.; Hussain, S.M. Effect of gold nanorod surface chemistry on cellular response. ACS Nano 2011, 5, 2870–2879. [Google Scholar] [CrossRef] [PubMed]
- Abo-Zeid, M.A.M.; Liehr, T.; Glei, M.; Gamal-Eldeen, A.M.; Zawrah, M.; Ali, M. Detection of cyto- and genotoxicity of rod-shaped gold nanoparticles in human blood lymphocytes using comet-FISH. Cytologia 2015, 80, 173–181. [Google Scholar] [CrossRef]
- Bhattacharya, R.; Patra, C.R.; Verma, R.; Kumar, S.; Greipp, P.R.; Mukherjee, P. Gold nanoparticles inhibit the proliferation of multiple myeloma cells. Adv. Mater. 2010, 19, 711–716. [Google Scholar] [CrossRef]
- Pernodet, N.; Fang, X.H.; Sun, Y.; Bakhtina, A.; Ramakrishnan, A.; Sokolov, J.; Ulman, A.; Rafailovich, M. Adverse effects of citrate/gold nanoparticles on human dermal fibroblasts. Small 2006, 2, 766–773. [Google Scholar] [CrossRef] [PubMed]
- Mironava, T.; Hadjiargyrou, M.; Simon, M.; Rafailovich, M.H. Gold nanoparticles cellular toxicity and recovery: Adipose derived stromal cells. Nanotoxicology 2014, 8, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Uboldi, C.; Bonacchi, D.; Lorenzi, G.; Hermanns, M.I.; Pohl, C.; Baldi, G.; Unger, R.E.; Kirkpatrick, C.J. Gold nanoparticles induce cytotoxicity in the alveolar type-II cell lines A549 and NCIH441. Part. Fibre Toxicol. 2009, 6, 18. [Google Scholar] [CrossRef] [PubMed]
- Freese, C.; Uboldi, C.; Gibson, M.I.; Unger, R.E.; Weksler, B.B.; Romero, I.A.; Couraud, P.O.; Kirkpatrick, C.J. Uptake and cytotoxicity of citrate-coated gold nanospheres: Comparative studies on human endothelial and epithelial cells. Part. Fibre Toxicol. 2012, 9, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karthikeyan, B.; Kalishwaralal, K.; Sheikpranbabu, S.; Deepak, V.; Haribalaganesh, R.; Gurunathan, S. Gold nanoparticles downregulate VEGF-and IL-1β-induced cell proliferation through Src kinase in retinal pigment epithelial cells. Exp. Eye Res. 2010, 91, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, R.; Mukherjee, P.; Xiong, Z.; Atala, A.; Soker, S.; Mukhopadhyay, D. Gold nanoparticles inhibit VEGF165-induced proliferation of HUVEC cells. Nano Lett. 2004, 4, 2479–2481. [Google Scholar] [CrossRef]
- Hengartner, M.O. The biochemistry of apoptosis. Nature 2000, 407, 770–776. [Google Scholar] [CrossRef] [PubMed]
- Szegezdi, E.; Logue, S.E.; Gorman, A.M.; Samali, A. Mediators of endoplasmic reticulum stress-induced apoptosis. EMBO Rep. 2006, 7, 880–885. [Google Scholar] [CrossRef] [PubMed]
- Kressel, M.; Groscurth, P. Distinction of apoptotic and necrotic cell death by in situ labelling of fragmented DNA. Cell Tissue Res. 1994, 278, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Schaeublin, N.M.; Braydich-Stolle, L.K.; Maurer, E.I.; Park, K.; MacCuspie, R.I.; Afrooz, A.R.M.N.; Vaia, R.A.; Saleh, N.B.; Hussain, S.M. Does shape matter? Bioeffects of gold nanomaterials in a human skin cell model. Langmuir 2012, 28, 3248–3258. [Google Scholar] [CrossRef] [PubMed]
- Cho, W.-S.; Kim, S.; Han, B.S.; Son, W.C.; Jeong, J. Comparison of gene expression profiles in mice liver following intravenous injection of 4 and 100 nm-sized PEG-coated gold nanoparticles. Toxicol. Lett. 2009, 191, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Quan, Y.-Y.; Wang, X.-P.; Chen, T.-S. Gold nanoparticles of diameter 13 nm induce apoptosis in rabbit articular chondrocytes. Nanoscale Res. Lett. 2016, 11, 249. [Google Scholar] [CrossRef] [PubMed]
- Di Bucchianico, S.; Fabbrizi, M.R.; Cirillo, S.; Uboldi, C.; Gilliland, D.; Valsami-Jones, E.; Migliore, L. Aneuploidogenic effects and DNA oxidation induced in vitro by differently sized gold nanoparticles. Int. J. Nanomed. 2014, 9, 2191–2204. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Clift, M.J.D.; Casey, A.; del Pino, P.; Pelaz, B.; Conde, J.; Byrne, H.J.; Rothen-Rutishauser, B.; Estrada, G.; de la Fuente, J.M.; Stoeger, T. Investigating the role of shape on the biological impact of gold nanoparticles in vitro. Nanomedicine 2015, 10, 2643–2657. [Google Scholar] [CrossRef] [PubMed]
- Schaeublin, N.M.; Braydich-Stolle, L.K.; Schrand, A.M.; Miller, J.M.; Hutchison, J.; Schlager, J.J.; Hussain, S.M. Surface charge of gold nanoparticles mediates mechanism of toxicity. Nanoscale 2011, 3, 410–420. [Google Scholar] [CrossRef] [PubMed]
- Durocher, I.; Noël, C.; Lavastre, V.; Girard, D. Evaluation of the in vitro and in vivo proinflammatory activities of gold(+) and gold(−) nanoparticles. Inflamm. Res. 2017, 66, 981–992. [Google Scholar] [CrossRef] [PubMed]
- Muthukumarasamyvel, T.; Rajendran, G.; Santhana Panneer, D.; Kasthuri, J.; Kathiravan, K.; Rajendiran, N. Role of surface hydrophobicity of dicationic amphiphile-stabilized gold nanoparticles on A549 lung cancer cells. ACS Omega 2017, 2, 3527–3538. [Google Scholar] [CrossRef]
- Zhang, C.; Zhou, Z.; Zhi, X.; Ma, Y.; Wang, K.; Wang, Y.; Zhang, Y.; Fu, H.; Jin, W.; Pan, F.; et al. Insights into the distinguishing stress-induced cytotoxicity of chiral gold nanoclusters and the relationship with GSTP1. Theranostics 2015, 5, 134–149. [Google Scholar] [CrossRef] [PubMed]
- De Araújo, R.F.; de Araújo, A.A.; Pessoa, J.B.; Freire Neto, F.P.; da Silva, G.R.; Leitão Oliveira, A.L.C.S.; de Carvalho, T.G.; Silva, H.F.O.; Eugênio, M.; Sant’Anna, C.; et al. Anti-inflammatory, analgesic and anti-tumor properties of gold nanoparticles. Pharmacol. Rep. 2017, 69, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Pavlovich, E.; Volkova, N.; Yakymchuk, E.; Perepelitsyna, O.; Sydorenko, M.; Goltsev, A. In vitro study of influence of Au nanoparticles on HT29 and SPEV cell lines. Nanoscale Res. Lett. 2017, 12, 494. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Xu, K.; Ji, L.; Tang, B. Effect of gold nanoparticles on glutathione depletion-induced hydrogen peroxide generation and apoptosis in HL7702 cells. Toxicol. Lett. 2011, 205, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, D.; Xavier, P.L.; Chaudhari, K.; John, R.; Dasgupta, A.K.; Pradeep, T.; Chakrabarti, G. Unprecedented inhibition of tubulin polymerization directed by gold nanoparticles inducing cell cycle arrest and apoptosis. Nanoscale 2013, 5, 4476–4489. [Google Scholar] [CrossRef] [PubMed]
- Teles, M.; Soares, A.M.V.M.; Tort, L.; Guimarães, L.; Oliveira, M. Linking cortisol response with gene expression in fish exposed to gold nanoparticles. Sci. Total Environ. 2017, 584, 1004–1011. [Google Scholar] [CrossRef] [PubMed]
- Ramalingam, V.; Revathidevi, S.; Shanmuganayagam, T.S.; Muthulakshmi, L.; Rajaram, R. Gold nanoparticle induces mitochondria-mediated apoptosis and cell cycle arrest in nonsmall cell lung cancer cells. Gold Bull. 2017, 50, 177–189. [Google Scholar] [CrossRef]
- Dong, L.; Li, M.; Zhang, S.; Li, J.; Shen, G.; Tu, Y.; Zhu, J.; Tao, J. Cytotoxicity of BSA-stabilized gold nanoclusters: In vitro and in vivo study. Small 2015, 11, 2571–2581. [Google Scholar] [CrossRef] [PubMed]
- Imperatore, R.; Carotenuto, G.; Di Grazia, M.A.; Ferrandino, I.; Palomba, L.; Mariotti, R.; Vitale, E.; De Nicola, S.; Longo, A.; et al. Imidazole-stabilized gold nanoparticles induce neuronal apoptosis: An in vitro and in vivo study. J. Biomed. Mater. Res. A 2015, 103, 1436–1446. [Google Scholar] [CrossRef] [PubMed]
- Cho, W.-S.; Cho, M.; Jeong, J.; Choi, M.; Cho, H.-Y.; Han, B.S.; Kim, S.H.; Kim, H.O.; Lim, Y.T.; Chung, B.H.; et al. Acute toxicity and pharmacokinetics of 13 nm-sized PEG-coated gold nanoparticles. Toxicol. Appl. Pharm. 2009, 236, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Ahamed, M.; Akhtar, M.J.; Khan, M.A.M.; Alhadlaq, H.A.; Alrokayan, S.A. Cytotoxic response of platinum-coated gold nanorods in human breast cancer cells at very low exposure levels. Environ. Toxicol. 2016, 31, 1344–1356. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Li, Q.; Ni, W.; Zhang, N.; Zheng, X.; Wang, Y.; Shao, D.; Tai, G. Cytotoxicity of various types of gold-mesoporous silica nanoparticles in human breast cancer cells. Int. J. Nanomed. 2015, 10, 6075–6087. [Google Scholar]
- Wang, Y.; Wan, J.; Miron, R.J.; Zhao, Y.; Zhang, Y. Antibacterial properties and mechanisms of gold-silver nanocages. Nanoscale 2016, 8, 11143–11152. [Google Scholar] [CrossRef] [PubMed]
- Chuang, S.-M.; Lee, Y.-H.; Liang, R.-Y.; Roam, G.-D.; Zeng, Z.-M.; Tu, H.-F.; Wang, S.-K.; Chueh, P.J. Extensive evaluations of the cytotoxic effects of gold nanoparticles. BBA Gen. Subj. 2013, 1830, 4960–4973. [Google Scholar] [CrossRef] [PubMed]
- Chueh, P.J.; Liang, R.-Y.; Lee, Y.-H.; Zeng, Z.-M.; Chuang, S.-M. Differential cytotoxic effects of gold nanoparticles in different mammalian cell lines. J. Hazard. Mater. 2014, 264, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhu, X.; Gong, J.; Sun, Y.; Chen, D.; Wang, J.; Wang, Y.; Guo, M.; Li, W. Lysosome-mitochondria-mediated apoptosis specifically evoked in cancer cells induced by gold nanorods. Nanomedicine 2016, 11, 1993–2006. [Google Scholar] [CrossRef] [PubMed]
- Noël, C.; Simard, J.-C.; Girard, D. Gold nanoparticles induce apoptosis, endoplasmic reticulum stress events and cleavage of cytoskeletal proteins in human neutrophils. Toxicol. In Vitro 2016, 31, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Neuss, S.; Leifert, A.; Fischler, M.; Wen, F.; Simon, U.; Schmid, G.; Brandau, W.; Jahnen-Dechent, W. Size-dependent cytotoxicity of gold nanoparticles. Small 2007, 3, 1941–1949. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.-H.; Li, W.-T.; Hung, W.-I.; Chen, C.-P.; Yeh, J.-M. Cytotoxicity and differentiation effects of gold nanoparticles to human bone marrow mesenchymal stem cells. Biomed. Eng. Appl. Basis Commun. 2011, 23, 141–152. [Google Scholar] [CrossRef]
- Abdelhalim, M.A.K.; Jarrar, B.M. Gold nanoparticles induced cloudy swelling to hydropic degeneration, cytoplasmic hyaline vacuolation, polymorphism, binucleation, karyopyknosis, karyolysis, karyorrhexis and necrosis in the liver. Lipids Health Dis. 2011, 10, 166. [Google Scholar] [CrossRef] [PubMed]
- Saleh, H.M.; Soliman, O.A.; Elshazly, M.O.; Raafat, A.; Gohar, A.K.; Salaheldin, T.A. A cute hematologic, hepatologic, and nephrologic changes after intraperitoneal injections of 18 nm gold nanoparticles in hamsters. Int. J. Nanomed. 2016, 11, 2505–2513. [Google Scholar] [CrossRef] [PubMed]
- Bhamidipati, M.; Fabris, L. Multiparametric assessment of gold nanoparticle cytotoxicity in cancerous and healthy cells: The role of size, shape, and surface chemistry. Bioconjug. Chem. 2017, 28, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Kim, Y.J.; Lee, J.W.; Lee, Y.; Lee, S.; Lim, Y.-B.; Chung, H.W. Cytotoxicity and genotoxicity induced by photothermal effects of colloidal gold nanorods. J. Nanosci. Nanotechnol. 2013, 13, 4437–4445. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.L.; Chen, D.; Wang, Y.; Zhang, L.; Dong, W.; Dai, J.X.; Jin, C.; Dong, X.; Sun, Y.; Zhao, H.; et al. Lysosome-dependent necrosis specifically evoked in cancer cells by gold nanorods. Nanomedicine 2017, 12, 1575–1589. [Google Scholar] [CrossRef] [PubMed]
- Kondath, S.; Raghavan, B.S.; Anantanarayanan, R.; Rajaram, R. Synthesis and characterisation of morin reduced gold nanoparticles and its cytotoxicity in MCF-7 cells. Chem.-Biol. Interact. 2014, 224, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Ramalingam, V.; Revathidevi, S.; Shanmuganayagam, T.S.; Muthulakshmi, L.; Rajaram, R. Biogenic gold nanoparticles induce cell cycle arrest through oxidative stress and sensitize mitochondrial membranes in A549 lung cancer cells. RSC Adv. 2016, 6, 20598–20608. [Google Scholar] [CrossRef]
- Nirmala, J.G.; Akila, S.; Narendhirakannan, R.T.; Chatterjee, S. Vitis vinifera peel polyphenols stabilized gold nanoparticles induce cytotoxicity and apoptotic cell death in A431 skin cancer cell lines. Adv. Powder Technol. 2017, 28, 1170–1184. [Google Scholar] [CrossRef]
- El-Kassas, H.Y.; El-Sheekh, M.M. Cytotoxic activity of biosynthesized gold nanoparticles with an extract of the red seaweed corallina officinalis on the MCF-7 human breast cancer cell line. Asian Pac. J. Cancer Prev. 2014, 15, 4311–4317. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhao, Y.; Zhang, X. Knockdown of glutamate cysteine ligase catalytic subunit by siRNA causes the gold nanoparticles-induced cytotoxicity in lung cancer cells. PLoS ONE 2015, 10, e0118870. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Gu, X.H.; Zhang, K.; Ding, Y.; Wei, X.B.; Zhang, X.M.; Zhao, Y.X. Gold nanoparticles trigger apoptosis and necrosis in lung cancer cells with low intracellular glutathione. J. Nanopart. Res. 2013, 15, 1745. [Google Scholar] [CrossRef]
- Glick, D.; Barth, S.; Macleod, K.F. Autophagy: Cellular and molecular mechanisms. J. Pathol. 2010, 221, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Menzies, F.M.; Fleming, A.; Caricasole, A.; Bento, C.F.; Andrews, S.P.; Ashkenazi, A.; Füllgrabe, J.; Jackson, A.; Jimenez Sanchez, M.; Karabiyik, C.; et al. Autophagy and neurodegeneration: Pathogenic mechanisms and therapeutic opportunities. Neuron 2017, 93, 1015–1034. [Google Scholar] [CrossRef] [PubMed]
- Amaravadi, R.; Kimmelman, A.C.; White, E. Recent insights into the function of autophagy in cancer. Genes Dev. 2016, 30, 1913–1930. [Google Scholar] [CrossRef] [PubMed]
- Deretic, V.; Saitoh, T.; Akira, S. Autophagy in infection, inflammation and immunity. Nat. Rev. Immunol. 2013, 13, 722–737. [Google Scholar] [CrossRef] [PubMed]
- Rubinsztein, D.C.; Mariño, G.; Kroemer, G. Autophagy and aging. Cell 2011, 146, 682–695. [Google Scholar] [CrossRef] [PubMed]
- Levine, B.; Kroemer, G. Autophagy in the pathogenesis of disease. Cell 2008, 132, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Karantza-Wadsworth, V.; Patel, S.; Kravchuk, O.; Chen, G.; Mathew, R.; Jin, S.; White, E. Autophagy mitigates metabolic stress and genome damage in mammary tumorigenesis. Genes Dev. 2007, 21, 1621–1635. [Google Scholar] [CrossRef] [PubMed]
- Degenhardt, K.; Mathew, R.; Beaudoin, B.; Bray, K.; Anderson, D.; Chen, G.; Mukherjee, C.; Shi, Y.; Gélinas, C.; Fan, Y.; et al. Autophagy promotes tumor cell survival and restricts necrosis, inflammation, and tumorigenesis. Cancer Cell 2006, 10, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Yang, M.; Kang, R.; Wang, Z.; Zhao, Y.; Yu, Y.; Xie, M.; Yin, X.; Livesey, K.M.; Loze, M.T.; et al. DAMP-mediated autophagy contributes to drug resistance. Autophagy 2011, 7, 112–114. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Wei, M.; Li, S.; Le, W. Nanomaterial-modulated autophagy: Underlying mechanisms and functional consequences. Nanomedicine 2016, 11, 1417–1430. [Google Scholar] [CrossRef] [PubMed]
- Lucocq, J.M.; Hacker, C. Cutting a fine figure: On the use of thin sections in electron microscopy to quantify autophagy. Autophagy 2013, 9, 1443–1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Duan, J.; Yu, Y.; Li, Y.; Liu, X.; Zhou, X.; Ho, K.-f.; Tian, L.; Sun, Z. Silica nanoparticles induce autophagy and autophagic cell death in HepG2 cells triggered by reactive oxygen species. J. Hazard. Mater. 2014, 270, 176–186. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Hartono, D.; Ong, C.-N.; Bay, B.-H.; Yung, L.-Y.L. Autophagy and oxidative stress associated with gold nanoparticles. Biomaterials 2010, 31, 5996–6003. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhang, Y.; Zhang, C.; Cui, X.; Zhai, S.; Liu, Y.; Li, C.; Zhu, H.; Qu, G.; Jiang, G.; et al. Tuning cell autophagy by diversifying carbon nanotube surface chemistry. ACS Nano 2014, 8, 2087–2099. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Zhou, H.; Gao, J. Nanoparticles modulate autophagic effect in a dispersity-dependent manner. Sci. Rep. (UK) 2015, 5, 14361. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Wu, Y.; Jin, S.; Tian, Y.; Zhang, X.; Zhao, Y.; Yu, L.; Liang, X.-J. Gold nanoparticles induce autophagosome accumulation through size-dependent nanoparticle uptake and lysosome impairment. ACS Nano 2011, 5, 8629–8639. [Google Scholar] [CrossRef] [PubMed]
- Ding, F.; Li, Y.; Liu, J.; Liu, L.; Yu, W.; Wang, Z.; Ni, H.; Liu, B.; Chen, P. Overendocytosis of gold nanoparticles increases autophagy and apoptosis in hypoxic human renal proximal tubular cells. Int. J. Nanomed. 2014, 9, 4317–4330. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Kong, N.; Zhang, Y.; Yang, W.; Yan, F. Size-dependent effects of gold nanoparticles on osteogenic differentiation of human periodontal ligament progenitor cells. Theranostics 2017, 7, 1214–1224. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Ji, Y.; Wu, X.; Xu, H. Trafficking of gold nanorods in breast cancer cells: Uptake, lysosome maturation, and elimination. ACS Appl. Mater. Interfaces 2013, 5, 9856–9865. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Wang, J.-H.; Liu, T.; Xie, Z.; Yu, X.-F.; Li, W. Surface chemistry but not aspect ratio mediates the biological toxicity of gold nanorods in vitro and in vivo. Sci. Rep. (UK) 2015, 5, 11398. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, C.; Cao, W.; Ma, B.; Ma, X.; Jin, S.; Zhang, J.; Wang, P.C.; Li, F.; Liang, X.-J. Anchoring effects of surface chemistry on gold nanorods: Modulating autophagy. J. Mater. Chem. B 2015, 3, 3324–3330. [Google Scholar] [CrossRef] [PubMed]
- Ma, N.; Liu, P.; He, N.; Gu, N.; Wu, F.-G.; Chen, Z. Action of gold nanospikes-based nanoradiosensitizers: Cellular internalization, radiotherapy, and autophagy. ACS Appl. Mater. Interfaces 2017, 9, 31526–31542. [Google Scholar] [CrossRef] [PubMed]
- Bhowmik, T.; Gomes, A. NKCT1 (purified Naja kaouthia protein toxin) conjugated gold nanoparticles induced Akt/mTOR inactivation mediated autophagic and caspase 3 activated apoptotic cell death in leukemic cell. Toxicon 2016, 121, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Joshi, P.; Chakraborti, S.; Ramirez-Vick, J.E.; Ansari, Z.A.; Shanker, V.; Chakrabarti, P.; Singh, S.P. The anticancer activity of chloroquine-gold nanoparticles against MCF-7 breast cancer cells. Colloid Surf. B 2012, 95, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Ke, S.; Zhou, T.; Yang, P.; Wang, Y.; Zhang, P.; Chen, K.; Ren, L.; Ye, S. Gold nanoparticles enhance TRAIL sensitivity through Drpl-mediated apoptotic and autophagic mitochondrial fission in NSCLC cells. Int. J. Nanomed. 2017, 12, 2531–2551. [Google Scholar] [CrossRef] [PubMed]
- Haynes, B.; Zhang, Y.; Liu, F.; Li, J.; Petit, S.; Kothayer, H.; Bao, X.; Westwell, A.D.; Mao, G.; Shekhar, M.P.V. Gold nanoparticle conjugated Rad6 inhibitor induces cell death in triple negative breast cancer cells by inducing mitochondrial dysfunction and PARP-1 hyperactivation: Synthesis and characterization. Nanomedicine 2016, 12, 745–757. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Wu, J.; Wang, F.; Liu, B.; Huang, C.; Wei, Y. Deconvoluting the role of reactive oxygen species and autophagy in human diseases. Free Radic. Biol. Med. 2013, 65, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Scherz-Shouval, R.; Shvets, E.; Fass, E.; Shorer, H.; Gil, L.; Elazar, Z. Reactive oxygen species are essential for autophagy and specifically regulate the activity of Atg4. EMBO J. 2007, 26, 1749–1760. [Google Scholar] [CrossRef] [PubMed]
- Syntichaki, P.; Tavernarakis, N. The biochemistry of neuronal necrosis: Rogue biology? Nat. Rev. Neurosci. 2003, 4, 672–684. [Google Scholar] [CrossRef] [PubMed]
- Jiao, P.; Zhou, H.; Otto, M.; Mu, Q.; Li, L.; Su, G.; Zhang, Y.; Butch, E.R.; Snyder, S.E.; Jiang, G.; et al. Leading neuroblastoma cells to die by multiple premeditated attacks from a multifunctionalized nanoconstruct. J. Am. Chem. Soc. 2011, 133, 13918–13921. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Jiao, P.; Yang, L.; Li, X.; Yan, B. Enhancing cell recognition by scrutinizing cell surfaces with a nanoparticle array. J. Am. Chem. Soc. 2011, 133, 680–682. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Properties | Cell Type | Cell Death | Ref. | |||||
|---|---|---|---|---|---|---|---|---|
| Size | Surface | Normal | Cancer | Apoptosis | Necrosis | Autophagy | ||
| GNPs | 1 nm | BSA | HepG-2 | + | + | N.A. | [65] | |
| 1.5 nm | TMAT (positive charge), MES (negative charge) | HaCaT | + | − | N.A. | [55] | ||
| 2.3 nm | l/d-glutathione | MGC-803 | +(d-glutathione > l-glutathione) | − | N.A. | [58] | ||
| 2.7 nm | 2-mercapto-1-methylimidazole | SH-SY5Y | + | N.A. | N.A. | [66] | ||
| 1–10 nm | sodium polyacrylate (negative charge) | freshly isolated human neutrophils | + | N.A. | N.A. | [56] | ||
| 7.4 nm | PVP | HT-29 | + | − | N.A. | [59] | ||
| 13 nm | citrate | Primary rabbit articular chondrocytes | + | − | N.A. | [52] | ||
| 13 nm | PEG | Liver of BALB/c mice | + | N.A. | N.A. | [67] | ||
| 20–25 nm | DCaLC, DCaC, DcaDC Hydrophobicity: DCaLC > DCaDC > DCaC | A549 | +(DCaLC > DCaDC > DCaC) | N.A. | N.A. | [57] | ||
| 29 nm | NaBH4 | A549 | + | N.A. | N.A. | [64] | ||
| 40 nm | citrate | head kidney of Sparus aurata | + | N.A. | N.A. | [63] | ||
| 40 nm | PVP | head kidney of Sparus aurata | + | N.A. | N.A. | [63] | ||
| 10–40 nm | N.A. | Vero cells | + | N.A. | N.A. | [72] | ||
| 4 nm, 100 nm | PEG | liver tissue of BALB/c mice | +(4 nm ≈ 100 nm) | N.A. | N.A. | [51] | ||
| 8, 15, 40 nm | citrate | SPEV, HL7702 | HT-29, A549 | + | + | N.A. | [60,61,62] | |
| 5 nm, 15 nm | citrate | human peripheral blood lymphocytes (PBL), Raw264.7 | +(15 nm > 5 nm) | + | N.A. | [53] | ||
| 20 nm, 70 nm | N.A. | Human polymorpho-nuclear neutrophil cells | + | N.A. | N.A. | [74] | ||
| GNRs | 16.7 nm × 43.8 nm | PEG | HaCaT | + | N.A. | N.A. | [50] | |
| 10 nm × 39 nm, 10 nm × 41 nm | N.A. | AGS | + | N.A. | N.A. | [71] | ||
| 50–60 nm × 20–30 nm | CTAB | MCF-7, N87 | + | + | N.A. | [73] | ||
| Hexagonal GNPs | 150 nm × 9 nm | HS–C2H4–CONH–PEG–O–C3H6–COOH | Calu-3 | + | N.A. | N.A. | [54] | |
| Properties | Cell Type | Cell Death | Ref. | |||||
|---|---|---|---|---|---|---|---|---|
| Size | Surface | Normal | Cancer | Apoptosis | Necrosis | Autophagy | ||
| GNPs | 1–2 nm | triphenylphosphine monosulfonate (TPPMS); tris-sulfonated triphenylphosphine (TPPTS) | L929; J774 A1 | HeLa; SK-Mel-28 | + | + | N.A. | [75] |
| 10 nm | N.A. | Rat hepatocyte | N.A. | + | N.A. | [77] | ||
| 14.6 nm | polyphenols | MCF-7 | N.A. | + | N.A. | [85] | ||
| 18 nm | citrate | Hamsters hepatocyte | N.A. | N.A. | + | N.A. | [78] | |
| 18.1 nm | citrate | 2M-125C | N.A. | + | + | N.A. | [76] | |
| 20 nm | morin | PBMCs, HBL-100 cells | − | − | N.A. | [82] | ||
| 20 nm | morin | MCF-7 | + | + | N.A. | [82] | ||
| 25.1 nm | CTAB | human dermal fibroblast (hfb) cells | − | + | N.A. | [79] | ||
| 25.1 nm | CTAB | U87 | + | − | N.A. | [79] | ||
| 25.1 nm | HSA | human dermal fibroblast (hfb) cells | U87 | − | − | N.A. | [79] | |
| 37 nm | N.A. | A549 | + | − | N.A. | [83] | ||
| 50 nm | polyphenols | A431 | + | + | N.A. | [84] | ||
| GNRs | 52.7 nm × 23.0 nm | CTAB | human dermal fibroblast (hfb) cells | − | + | N.A. | [79] | |
| 52.7 nm × 23.0 nm | HSA | human dermal fibroblast (hfb) cells | U87 | − | + | N.A. | [79] | |
| 119 × 28 nm | CTAB | normal human blood lymphocytes | − | − | N.A. | [80] | ||
| 119 × 28 nm | CTAB | A549 | − | + | N.A. | [80] | ||
| 20 × 53 nm | CTAB | CHO | − | − | N.A. | [81] | ||
| 20 × 53 nm | CTAB | MCF-7 | + | + | N.A. | [81] | ||
| GNSRs | The tip-to-tip distance: 62.5 ± 11.2 nm | CTAB | human dermal fibroblast (hfb) cells | − | + | N.A. | [79] | |
| The tip-to-tip distance: 62.5 ± 11.2 nm | CTAB | U87 | + | − | N.A. | [79] | ||
| The tip-to-tip distance: 62.5 ± 11.2 nm nm | HSA | human dermal fibroblast (hfb) cells | − | + | N.A. | [79] | ||
| The tip-to-tip distance: 62.5 ± 11.2 nm nm | HSA | U87 | − | − | N.A. | [79] | ||
| Properties | Cell Type | Cell Death | Ref. | |||||
|---|---|---|---|---|---|---|---|---|
| Size | Surface | Normal | Cancer | Apoptosis | Necrosis | Autophagy | ||
| GNPs | 5 nm | citrate | PDLPs | N.A. | N.A. | bloakade of autophagy | [105] | |
| 5 nm | citrate | HK-2 | + (only in hypoxic cells) | N.A. | + (in hypoxic cells > in normoxic cells) | [104] | ||
| 20 nm | citrate | MRC-5 | N.A. | N.A. | + | [100] | ||
| 10, 25, 50 nm | citrate | NRK | N.A. | N.A. | + (blockade of autophagic flux, 50 nm > 25 nm > 10 nm) | [103] | ||
| 13, 45 nm | citrate | PDLPs | N.A. | N.A. | + (45 nm > 13 nm) | [105] | ||
| GNRs | N.A. | CTAB | HCT116, BEL7402 | + | N.A. | + | [107] | |
| N.A. | CTAB | HEK293T, L02, HFF | PC3 | N.A. | N.A. | + | [107] | |
| N.A. | CTAB/PSS, CTAB/PAH, CTAB/PSS/PAH, CTAB/PAH/PSS | HCT116 | − | N.A. | − | [107] | ||
| 55 nm × 14 nm (length × diameter) | CTAB | A549 | N.A. | N.A. | + | [108] | ||
| PSS, PDDAC | A549 | N.A. | N.A. | − | [108] | |||
| average length of 10–40 nm | N.A. | Vero | + | N.A. | N.A. | [72] | ||
| average length of 10–40 nm | N.A. | NIH3T3 | − | N.A. | + | [72] | ||
| average length of 10–40 nm | N.A. | MRC-5 | − | N.A. | N.A. | [72] | ||
| GNSs | 54 ± 9 nm | no coating (GNSs), HS-PEG-NH2 (NH2-GNSs), HS-PEG-FA (FA-GNSs), cysteine-terminated TAT (TAT-GNSs) | KB | + (TAT-GNSs > FA-GNSs > NH2-GNSs > GNSs) | N.A. | + (TAT-GNSs > FA-GNSs > NH2-GNSs > GNSs) | [109] | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, H.; Jia, J.; Jiang, C.; Zhai, S. Gold Nanoparticle-Induced Cell Death and Potential Applications in Nanomedicine. Int. J. Mol. Sci. 2018, 19, 754. https://doi.org/10.3390/ijms19030754
Sun H, Jia J, Jiang C, Zhai S. Gold Nanoparticle-Induced Cell Death and Potential Applications in Nanomedicine. International Journal of Molecular Sciences. 2018; 19(3):754. https://doi.org/10.3390/ijms19030754
Chicago/Turabian StyleSun, Hainan, Jianbo Jia, Cuijuan Jiang, and Shumei Zhai. 2018. "Gold Nanoparticle-Induced Cell Death and Potential Applications in Nanomedicine" International Journal of Molecular Sciences 19, no. 3: 754. https://doi.org/10.3390/ijms19030754
APA StyleSun, H., Jia, J., Jiang, C., & Zhai, S. (2018). Gold Nanoparticle-Induced Cell Death and Potential Applications in Nanomedicine. International Journal of Molecular Sciences, 19(3), 754. https://doi.org/10.3390/ijms19030754