Tolerance to Plant Pathogens: Theory and Experimental Evidence

Abstract
:1. Introduction
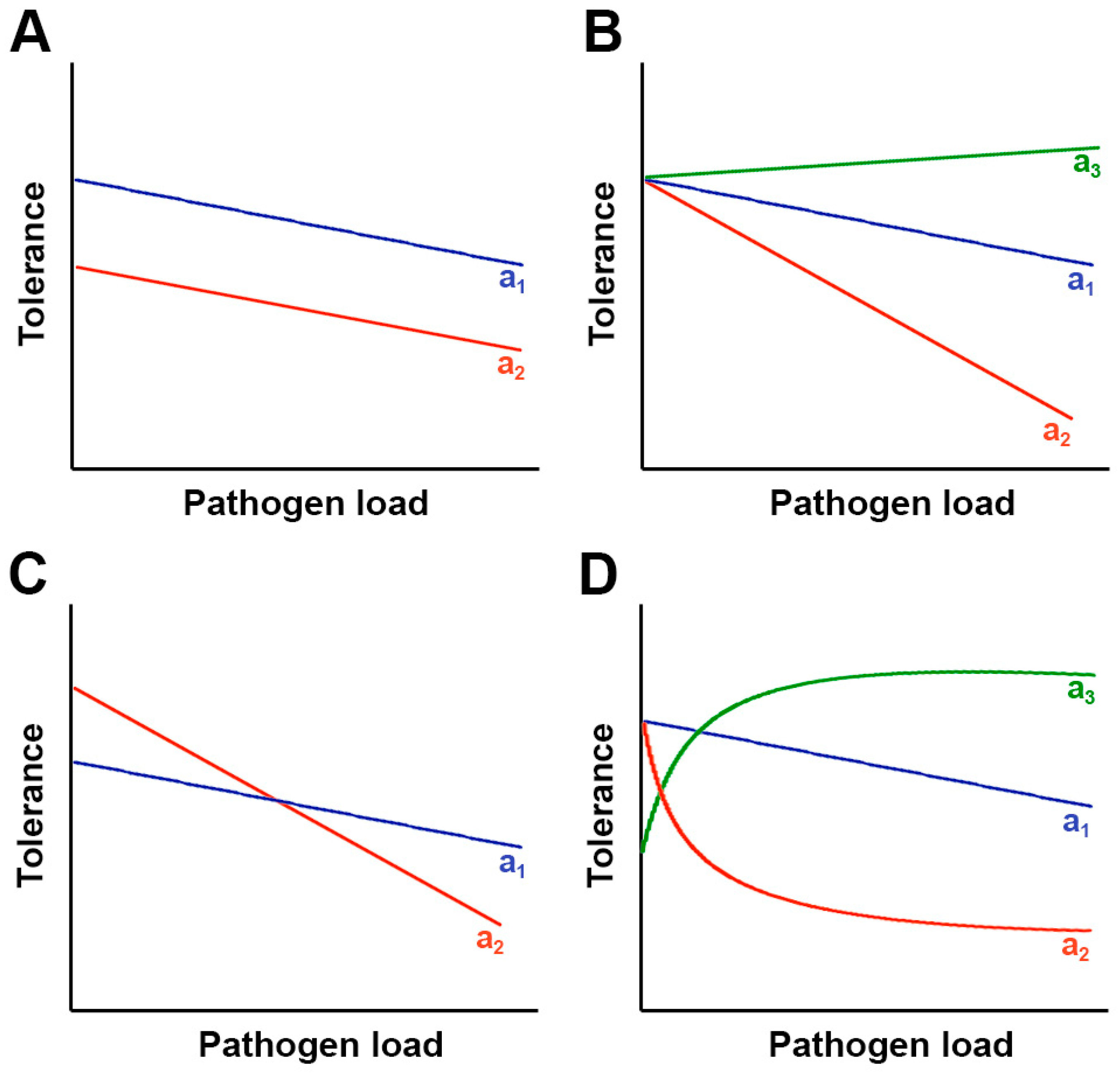
2. Theory on the Evolution of Tolerance to Pathogens
3. Measuring Tolerance to Pathogens
4. Experimental Evidence of Tolerance in Plant-Pathogen Interactions
4.1. Fungi and Oomycetes
4.2. Viruses
4.3. Bacteria
4.4. Parasitic Plants
5. Mechanistic Basis and Genetic Determinants of Plant Tolerance to Pathogens
6. Conclusions and Future Perspective
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Windsor, D.A. Most of the species on Earth are parasites. Int. J. Parasitol. 1998, 28, 1939–1941. [Google Scholar] [CrossRef]
- Read, A.F. The evolution of virulence. Trends Microbiol. 1994, 2, 73–76. [Google Scholar] [CrossRef]
- D’Arcy, C.J.; Eastburn, D.M.; Schumann, G.L. Illustrated glossary of plant pathology. Plant Health. Instr. 2001. [Google Scholar] [CrossRef]
- Anderson, P.K.; Cunningham, A.A.; Patel, N.G.; Morales, F.J.; Epstein, P.R.; Daszak, P. Emerging infectious diseases of plants: Pathogen pollution, climate change and agrotechnology drivers. Trends Ecol. Evol. 2004, 19, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, G.S. Evolutionary ecology of plant diseases in natural ecosystems. Annu. Rev. Phytopathol. 2002, 40, 13–43. [Google Scholar] [CrossRef] [PubMed]
- Pagán, I.; Fraile, A.; García-Arenal, F. Evolution of the interactions of viruses with their plant hosts. In Virus Evolution: Current Research and Future Directions; Weaver, S.C., Denison, M., Roossinck, M., Vignuzzi, M., Eds.; Caister Academic Press: Poole, UK, 2016; pp. 127–154. [Google Scholar]
- Alexander, H.M.; Mauck, K.E.; Whitfield, A.E.; Garrett, K.A.; Malmstrom, C.M. Plant-virus interactions and the agro-ecological interface. Eur. J. Plant Pathol. 2014, 138, 529–547. [Google Scholar] [CrossRef]
- Oerke, E.-C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Vurro, M.; Bonciani, B.; Vannacci, G. Emerging infectious diseases of crop plants in developing countries: Impact on agriculture and socio-economic consequences. Food Secur. 2010, 2, 113–132. [Google Scholar] [CrossRef]
- FAO. Keeping Plant Pests and Diseases at Bay: Experts Focus on Global Measures; FAO, 2015; http://www.fao.org/news/story/en/item/280489/icode/. [Google Scholar]
- Agnew, P.; Koella, J.C.; Michalakis, Y. Host life history responses to parasitism. Microbes Infect. 2000, 2, 891–896. [Google Scholar] [CrossRef]
- Clarke, D.D. Tolerance of parasites and disease in plants and its significance in host-parasite interactions. Adv. Plant Pathol. 1986, 5, 161–198. [Google Scholar]
- Strauss, S.Y.; Agrawal, A.A. The ecology and evolution of plant tolerance to herbivory. Trends Ecol. Evol. 1999, 14, 179–185. [Google Scholar] [CrossRef]
- Little, T.J.; Shuker, D.M.; Colegrave, N.; Day, T.; Graham, A.L. The coevolution of virulence: Tolerance in perspective. PLoS Pathog. 2010, 6, e1001006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Råberg, L. How to live with the enemy: Understanding tolerance to parasites. PLoS Biol. 2014, 12, e1001989. [Google Scholar] [CrossRef] [PubMed]
- Roy, B.A.; Kirchner, J.W. Evolutionary dynamics of pathogen resistance and tolerance. Evolution 2000, 54, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Vale, P.F.; Fenton, A.; Brown, S.P. Limiting damage during infection: Lessons from infection tolerance for novel therapeutics. PLoS Biol. 2014, 12, e1001769. [Google Scholar] [CrossRef] [PubMed]
- Van der Bosch, F.; Akudibalah, G.; Seal, S.; Jeger, M. Host resistance and the evolutionary response of plant viruses. J. Appl. Evol. 2006, 43, 506–516. [Google Scholar] [CrossRef]
- Restif, O.; Koella, J.C. Concurrent evolution of resistance and tolerance to pathogens. Am. Nat. 2004, 164, E90–E102. [Google Scholar] [CrossRef] [PubMed]
- Cobb, N. Contributions to an economic knowledge of Australian rusts (Uredineae). Agric. Gaz. N. S. Wales 1894, 5, 239–250. [Google Scholar]
- Calwell, R.H.; Schaffer, J.F.; Compton, L.E.; Patterson, F.L. Tolerance to cereal leaf rusts. Science 1958, 128, 714–715. [Google Scholar] [CrossRef] [PubMed]
- Kramer, T.; Gildemacher, B.H.; Van der Ster, M.; Parlevliet, J.E. Tolerance of spring barley cultivars to leaf rust, Puccinia hordei. Eyphytica 1980, 29, 209–216. [Google Scholar] [CrossRef]
- Sacristán, S.; García-Arenal, F. The evolution of virulence and pathogenicity in plant pathogen populations. Mol. Plant Pathol. 2008, 9, 369–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doumayrou, J.; Leblaye, S.; Froissart, R.; Michalakis, Y. Reduction of leaf area and symptom severity as proxies of disease-induced plant mortality: The example of the Cauliflower mosaic virus infecting two Brassicaceae hosts. Virus Res. 2013, 176, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Fineblum, W.L.; Rausher, M.D. Tradeoff between resistance and tolerance to herbivore damage in a morning glory. Nature 1995, 377, 517–520. [Google Scholar] [CrossRef]
- Baucom, R.S.; de Roode, J.C. Ecological immunology and tolerance in plants and animals. Funct. Ecol. 2011, 25, 18–28. [Google Scholar] [CrossRef]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef] [PubMed]
- Hudson, P.J.; Dobson, A.P.; Lafferty, K.D. Is a healthy ecosystem one that is rich in parasites? Trends Ecol. Evol. 2006, 21, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Ostfeld, R.S.; Keesing, F. Effects of host diversity on infectious disease. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 157–182. [Google Scholar] [CrossRef]
- Van der Meijden, E.; Wijn, H.; Verkaar, J. Defence and regrowth: Alternative plant strategies in the struggle against herbivores. Oikos 1988, 51, 355–363. [Google Scholar] [CrossRef]
- Herms, D.A.; Mattson, W.J. The dilemma of plants: To grow or defend. Q. Rev. Biol. 1992, 67, 283–335. [Google Scholar] [CrossRef]
- Belsky, A.J.; Carson, W.P.; Jensen, C.L.; Gordon, A.F. Overcompensation by plants: Herbivore optimization or red herring? Evol. Ecol. 1993, 7, 109–121. [Google Scholar] [CrossRef]
- Rosenthal, J.P.; Kotanen, P.M. Terrestrial plant tolerance to herbivory. Trends Ecol. Evol. 1994, 9, 145–148. [Google Scholar] [CrossRef]
- Mauricio, R.; Rausher, M.D.; Burdick, D.S. Variation in the defence strategies of plants: Are resistance and tolerance mutually exclusive? Ecology 1997, 78, 1301–1311. [Google Scholar]
- Boots, M.; Bowers, R.G. Three mechanisms of host resistance to microparasites—Avoidance, recovery and tolerance—Show different evolutionary dynamics. J. Theor. Biol. 1999, 201, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Fornoni, J.; Núñez-Farfán, J.; Valverde, P.L.; Rausher, M.D. Evolution of mixed strategies of plant defence allocation against natural enemies. Evolution 2004, 58, 1685–1695. [Google Scholar] [CrossRef] [PubMed]
- Restif, O.; Koella, J.C. Shared control of epidemiological traits in a coevolutionary model of host-parasite interactions. Am. Nat. 2003, 161, 827–836. [Google Scholar] [CrossRef] [PubMed]
- Lafferty, K.D.; Kuris, A.M. Parasitic castration: The evolution and ecology of body snatchers. Trends Parasitol. 2009, 25, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Best, A.; White, A.; Boots, M. Resistance is futile but tolerance explains why parasites do not castrate their hosts. Evolution 2010, 64, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Antonovics, J.; Thrall, P.H. The cost of resistance and the maintenance of genetic polymorphism in host-parasite systems. Proc. R. Soc. Lond. B Biol. Sci. 1994, 257, 105–110. [Google Scholar] [CrossRef]
- Bowers, R.G.; Boots, M.; Begon, M. Life-history trade-offs and the evolution of parasite resistance: Competition between host strains. Proc. R. Soc. Lond. B Biol. Sci. 1994, 257, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Boots, M.; Haraguchi, Y. The evolution of costly resistance in host-parasite systems. Am. Nat. 1999, 153, 359–370. [Google Scholar]
- Miller, M.R.; White, A.; Boots, M. The evolution of host resistance: Tolerance and control as distinct strategies. J. Theor. Biol. 2005, 236, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Best, A.; White, A.; Boots, M. Maintenance of host variation in tolerance to pathogens and parasites. Proc. Natl. Acad. Sci. USA 2008, 105, 20786–20791. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.M.; May, R. Coevolution of hosts and parasites. Parasitology 1982, 85, 411–426. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.R.; White, A.; Boots, M. The evolution of parasites in response to tolerance in their hosts: The good, the bad, and apparent commensalism. Evolution 2006, 60, 945–956. [Google Scholar] [CrossRef] [PubMed]
- van der Bosch, F.; Jeger, M.; Gilligan, C.A. Disease control and its selection for damaging plant virus strains in vegetatively propagated staple food crops; a theoretical assessment. Proc. R. Soc. Lond. B Biol. Sci. 2007, 274, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Kirchner, J.W.; Roy, B.A. Evolutionary implications of host-pathogen specificity: The fitness consequences of host life history traits. Evol. Ecol. 2001, 14, 665–692. [Google Scholar] [CrossRef]
- Detilleux, J.C. Effectiveness analysis of resistance and tolerance to infection. Genet. Sel. Evol. 2011, 43, 9. [Google Scholar] [CrossRef] [PubMed]
- Débarre, F.; Lion, S.; van Baalen, M.; Gandon, S. Evolution of host life-history traits in a spatially structured host-parasite system. Am. Nat. 2012, 179, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Horns, F.; Hood, E. The evolution of disease resistance and tolerance in spatially structured populations. Ecol. Evol. 2012, 2, 1705–1711. [Google Scholar] [CrossRef] [PubMed]
- Cousineau, S.V.; Alizon, S. Parasite evolution in response to sex-based host heterogeneity in resistance and tolerance. J. Evol. Biol. 2014, 27, 2753–2766. [Google Scholar] [CrossRef] [PubMed]
- Råberg, L.; Graham, A.L.; Read, A.F. Decomposing health: Tolerance and resistance to parasites in animals. Philos. Trans. R. Soc. Lond. B 2009, 364, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Schlichting, C.; Pigliucci, M. Phenotypic Plasticity: A Reaction Norm Perspective; Sinauer Associates: Sunderland, MA, USA, 1998. [Google Scholar]
- Simms, E.L. Defining tolerance as a reaction norm. Evol. Ecol. 2000, 14, 563–570. [Google Scholar] [CrossRef]
- Simms, E.; Triplett, J. Costs and benefits of plant responses to disease: Resistance and tolerance. Evolution 1994, 48, 1973–1985. [Google Scholar] [CrossRef] [PubMed]
- Kutzer, M.A.; Armitage, S.A. Maximising fitness in the face of parasites: A review of host tolerance. Zoology 2016, 119, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Carr, D.E.; Murphy, J.F.; Eubanks, M.D. The susceptibility and response of inbred and outbred Mimulus guttatus to infection by Cucumber mosaic virus. Evol. Ecol. 2003, 17, 85–103. [Google Scholar] [CrossRef]
- Hily, J.M.; Poulicard, N.; Mora, M.A.; Pagán, I.; García-Arenal, F. Environment and host genotype determine the outcome of a plant-virus interaction: From antagonism to mutualism. New Phytol. 2016, 209, 812–822. [Google Scholar] [CrossRef] [PubMed]
- Kause, A. Genetic analysis of tolerance to infections using random regressions: A simulation study. Genet. Res. 2011, 93, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Kause, A.; Ødegård, J. The genetic analysis of tolerance to infections: A review. Front. Genet. 2012, 3, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Maynard Smith, J. Evolutionary Genetics; Oxford University Press: London, UK, 1989. [Google Scholar]
- Day, T. On the evolution of virulence and the relationship between various measures of mortality. Proc. R. Soc. Lond. B Biol. Sci. 2002, 269, 1317–1323. [Google Scholar] [CrossRef] [PubMed]
- Rohr, J.R.; Raffel, T.R.; Hall, C.A. Developmental variation in resistance and tolerance in a multi-host–parasite system. Funct. Ecol. 2010, 24, 1110–1121. [Google Scholar] [CrossRef]
- Pagán, I.; Alonso-Blanco, C.; García-Arenal, F. The relationship of within-host multiplication and virulence in a plant-virus system. PLoS ONE 2007, 2, e786. [Google Scholar]
- Pagán, I.; Alonso-Blanco, C.; García-Arenal, F. Host responses in life-history traits and tolerance to virus infection in Arabidopsis thaliana. PLoS Pathog. 2008, 4, e1000124. [Google Scholar]
- Vijayan, V.; López-González, S.; Sánchez, F.; Ponz, F.; Pagán, I. Virulence evolution of a sterilizing plant virus: Tuning multiplication and resource exploitation. Virus Evol. 2017, 3, vex033. [Google Scholar] [CrossRef] [PubMed]
- Alexander, H.M.; Bruns, E.; Schebor, H.; Malmstrom, C.M. Crop-associated virus infection in a native perennial grass: Reduction in plant fitness and dynamic patterns of virus detection. J. Ecol. 2017, 105, 1021–1031. [Google Scholar] [CrossRef]
- Fraile, A.; McLeish, M.J.; Pagán, I.; González-Jara, P.; Piñero, P.; García-Arenal, F. Environmental heterogeneity and the evolution of plant-virus interactions: Viruses in wild pepper populations. Virus Res. 2017, 241, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Goss, E.M.; Bergelson, J. Fitness consequences of infection of Arabidopsis thaliana with its natural bacterial pathogen Pseudomonas viridiflava. Oecologia 2007, 152, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Politowski, K.; Browning, J.A. Tolerance and resistance to plant disease: An epidemiological study. Phytopathology 1978, 68, 1177–1185. [Google Scholar] [CrossRef]
- Roberts, J.J.; Hendricks, L.T.; Patterson, F.L. Tolerance to leaf rust in susceptible wheat cultivars. Phytopathology 1984, 74, 349–351. [Google Scholar] [CrossRef]
- Newton, A.C. Exploitation of diversity within crops—The key to disease tolerance? Front. Plant Sci. 2016, 7, 665. [Google Scholar] [CrossRef] [PubMed]
- Ben-Kalio, V.D.; Clarke, D.D. Studies on tolerance in wild plants: Effects of Erysiphe fischeri on the growth and development of Senecio vulgaris. Physiol. Plant Pathol. 1979, 14, 203–211. [Google Scholar] [CrossRef]
- Roy, B.A.; Kirchner, J.W.; Christian, C.E.; Rose, L.E. High disease incidence and apparent disease tolerance in a North American Great Basin plant community. Evol. Ecol. 2000, 14, 421–438. [Google Scholar] [CrossRef]
- Inglese, S.J.; Paul, N.D. Tolerance of Senecio vulgaris to infection and disease caused by native and alien rust fungi. Phytopathology 2006, 96, 718–726. [Google Scholar] [CrossRef] [PubMed]
- Salvaudon, L.; Héraudet, V.; Shykoff, J.A. Genotype-specific interactions and the trade-off between host and parasite fitness. BMC Evol. Biol. 2007, 7, 189. [Google Scholar] [CrossRef] [PubMed]
- Salvaudon, L.; Héraudet, V.; Shykoff, J.A. Arabidopsis thaliana and the Robin Hood parasite: A chivalrous oomycete that steals fitness from fecund hosts and benefits the poorest one? Biol. Lett. 2008, 4, 526–529. [Google Scholar] [CrossRef] [PubMed]
- Salvaudon, L.; Shykoff, J.A. Variation in Arabidopsis developmental responses to oomycete infection: Resilience vs. changes in life history traits. New Phytol. 2013, 197, 919–926. [Google Scholar] [CrossRef] [PubMed]
- Pilowsky, M.; Cohen, S. Tolerance to Tomato yellow leaf curl virus derived from Lycopersicon peruvianum. Plant Dis. 1990, 74, 248–250. [Google Scholar] [CrossRef]
- Rubio, L.; Herrero, J.R.; Sarrió, J.; Moreno, P.; Guerri, J. A new approach to evaluate relative resistance and tolerance of tomato cultivars to begomoviruses causing the tomato yellow leaf curl disease in Spain. Plant Pathol. 2003, 52, 763–769. [Google Scholar] [CrossRef]
- Jenkins, G. Comparison of tolerance to Barley yellow dwarf virus in barley and oats. Ann. Appl. Biol. 1966, 57, 163–168. [Google Scholar] [CrossRef]
- Jin, H.; Domier, L.L.; Kolb, F.L.; Brown, C.M. Identification of quantitative loci for tolerance to barley yellow dwarf virus in oat. Phytopathology 1998, 88, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Foresman, B.J.; Oliver, R.E.; Jackson, E.W.; Chao, S.; Arruda, M.P.; Kolb, F.L. Genome-wide association mapping of Barley yellow dwarf virus tolerance in spring oat (Avena sativa L.). PLoS ONE 2016, 11, e0155376. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.J.; Kline, A.S.; Morelock, T.E.; McNew, R.W. Tolerance to Blackeye cowpea mosaic potyvirus not correlated with decreased virus accumulation or protection from cowpea stunt disease. Plant Dis. 1996, 80, 847–852. [Google Scholar] [CrossRef]
- Desbiez, C.; Gal-On, A.; Girard, M.; Wipf-Scheibel, C.; Lecoq, H. Increase in Zucchini yellow mosaic virus symptom severity in tolerant zucchini cultivars is related to a point mutation in P3 protein and is associated with a loss of relative fitness on susceptible plants. Phytopathology 2003, 93, 1478–1484. [Google Scholar] [CrossRef] [PubMed]
- Desbiez, C.; Wipf-Scheibel, C.; Granier, F.; Robaglia, C.; Delaunay, T.; Lecoq, H. Biological and molecular variability of Zucchini yellow mosaic virus in the island of Martinique. Plant Dis. 1996, 80, 203–207. [Google Scholar] [CrossRef]
- Desbiez, C.; Wipf-Scheibel, C.; Lecoq, H. Biological and serological variability, evolution and molecular epidemiology of Zucchini yellow mosaic virus (ZYMV, Potyvirus) with special reference to Caribbean islands. Virus Res. 2002, 85, 5–16. [Google Scholar] [CrossRef]
- Carr, D.E.; Murphy, J.F.; Eubanks, M.D. Genetic variation and covariation for resistance and tolerance to Cucumber mosaic virus in Mimulus guttatus (Phrymaceae): A test for costs and constraints. Heredity 2006, 96, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Shuckla, A.; Pagán, I.; García-Arenal, F. Effective tolerance based on resource reallocation is a virus- specific defence in Arabidopsis thaliana. Mol. Plant Pathol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Pagán, I.; Alonso-Blanco, C.; García-Arenal, F. Differential tolerance to direct and indirect density-dependent costs of viral infection in Arabidopsis thaliana. PLoS Pathog. 2009, 5, e1000531. [Google Scholar]
- Xu, P.; Chen, F.; Mannas, J.P.; Feldman, T.; Sumner, L.W.; Roossinck, M.J. Virus infection improves drought tolerance. New Phytol. 2008, 180, 911–921. [Google Scholar] [CrossRef] [PubMed]
- Hily, J.M.; García, A.; Moreno, A.; Plaza, M.; Wilkinson, M.D.; Fereres, A.; Fraile, A.; García-Arenal, F. The relationship between host lifespan and pathogen reservoir potential: An analysis in the system Arabidopsis thaliana-Cucumber mosaic virus. PLoS Pathog. 2014, 10, e1004492. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, J.; Somerville, S.C.; Hammerschmidt, R. Identification of a gene in Arabidopsis thaliana that controls resistance to Xanthomonas campestris pv. campestris. Physiol. Mol. Plant Pathol. 1991, 38, 57–65. [Google Scholar] [CrossRef]
- Buell, C.R.; Somerville, S.C. Expression of defense-related and putative signaling genes during tolerant and susceptible interations of Arabidopsis with Xanthomonas campestris pv. campestris. Mol. Plant-Microbe Interact. 1995, 8, 435–443. [Google Scholar] [CrossRef]
- Jakob, K.; Goss, E.M.; Araki, H.; Van, T.; Kreitman, M.; Bergelson, J. Pseudomonas viridiflava and P. syringae--natural pathogens of Arabidopsis thaliana. Mol. Plant Microbe Interact. 2002, 15, 1195–1203. [Google Scholar] [CrossRef] [PubMed]
- Kover, P.X.; Schaal, B.A. Genetic variation for disease resistance and tolerance among Arabidopsis thaliana accessions. Proc. Natl. Acad. Sci. USA 2002, 99, 11270–11274. [Google Scholar] [CrossRef] [PubMed]
- Goss, E.M.; Bergelson, J. Variation in resistance and virulence in the interaction between Arabidopsis thaliana and a bacterial pathogen. Evolution 2006, 60, 1562–1573. [Google Scholar] [CrossRef] [PubMed]
- Medel, R. Assessment of correlational selection on tolerance and resistance traits in a host plant-parasitic plant interaction. Evol. Ecol. 2001, 15, 37–52. [Google Scholar] [CrossRef]
- Koskela, T.; Puustinen, S.; Salonen, V.; Mutikainen, P. Resistance and tolerance in a host plant–holoparasitic plant interaction: Genetic variation and costs. Evolution 2002, 56, 899–908. [Google Scholar] [PubMed]
- Rowntree, J.; Cameron, D.D.; Preziosi, R.F. Genetic variation changes the interactions between the parasitic plant-ecosystem engineer Rhinanthus and its hosts. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2011, 366, 1380–1388. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Kolb, F.L.; Kaeppler, H.F. Molecular mapping of genomic regions underlying barley yellow dwarf tolerance in cultivated oat (Avena sativa L.). Theor. Appl. Genet. 2003, 106, 1300–1306. [Google Scholar] [CrossRef] [PubMed]
- del Blanco, I.A.; Hegarty, J.; Gallagher, L.; Falk, B.W.; Brown-Guedira, G.; Pellerin, E.; Dubcovsky, J. Mapping of QTL for tolerance to Cereal yellow dwarf virus in two-rowed spring barley. Crop Sci. 2014, 54, 1468–1475. [Google Scholar] [CrossRef] [PubMed]
- Orcutt, D.M.; Nilsen, E.T. Physiology of Plants under Stress: Soul and Biotic Factors; John Wiley & Sons: New York, NY, USA, 2000. [Google Scholar]
- Scholes, J.D.; Farrar, J.F. Increased rates of photosynthesis in localized regions of a barley leaf infected with brown rust. New Phytol. 1986, 104, 601–612. [Google Scholar] [CrossRef]
- Stare, T.; Ramšak, Ž.; Blejec, A.; Stare, K.; Turnšek, N.; Weckwerth, W.; Wienkoop, S.; Vodnik, D.; Gruden, K. Bimodal dynamics of primary metabolism-related responses in tolerant potato-Potato virus Y interaction. BMC Genom. 2015, 16, 716. [Google Scholar] [CrossRef] [PubMed]
- Abbes, Z.; Kharrat, M.; Delavault, P.; Chaïbi, W.; Simier, P. Nitrogen and carbon relationships between the parasitic weed Orobanche foetida and susceptible and tolerant faba bean lines. Plant Physiol. Biochem. 2009, 47, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Stearns, S.C. The Evolution of Life Histories; Oxford University Press: London, UK, 1992. [Google Scholar]
- Perrin, N.; Christe, P. On host life-history response to parasitism. Oikos 1996, 75, 317–320. [Google Scholar] [CrossRef]
- Gandon, S.; Agnew, P.; Michalakis, Y. Coevolution between parasite virulence and host life-history traits. Am. Nat. 2002, 160, 374–388. [Google Scholar] [PubMed]
- Narayanasamy, P. Molecular Biology in Plant Pathogenesis and Disease Management: Disease Development; Springer: New York, NY, USA, 2008. [Google Scholar]
- Bent, A.F.; Innes, R.W.; Ecker, J.R.; Staskawitcz, B.J. Disease development in ethylene-insensitive Arabidopsis thaliana infected with virulent and avirulent Pseudomonas and Xanthomonas pathogens. Mol. Plant Microbe Interact. 1992, 5, 372–378. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, P.J.; Jones, J.B.; Antoine, F.R.; Ciardi, J.; Klee, H.J. Ethylene-dependent salicylic acid regulates an expanded cell death response to a plant pathogen. Plant J. 2001, 25, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Block, A.; Schmelz, E.; O’Donnell, P.J.; Jones, J.B.; Klee, H.J. Systemic acquired tolerance to virulent bacterial pathogens in tomato. Plant Physiol. 2005, 138, 1481–1490. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Devaiah, S.P.; Wang, C.; Li, M.; Welti, R.; Wang, X. Arabidopsis phospholipase Dβ1 modulates defense responses to bacterial and fungal pathogens. New Phytol. 2013, 199, 228–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Qin, L.; Zhao, J.; Muhammad, T.; Cao, H.; Li, H.; Zhang, Y.; Liang, Y. SlMAPK3 enhances tolerance to Tomato yellow leaf curl virus (TYLCV) by regulating salicylic acid and jasmonic acid signaling in tomato (Solanum lycopersicum). PLoS ONE 2017, 12, e0172466. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Tolerance | Pathogen | Species | Factor | Host | Fitness | Reference |
|---|---|---|---|---|---|---|
| RT | Fungus | Puccinia coronata | Host genotype | Avena sativa | Kernel weight | Polotowski & Browning, 1978 [71] |
| Fungus | Puccinia recondita | Host genotype | Triticum aestivum | Grain production | Roberts et al., 1984 [72] | |
| Fungus | Colletotricum dematium | Pathogen isolate; Host population site | Ipomea purpurea | Flower production | Simms & Triplett, 1994 [56] | |
| Fungus | Coleosporium tussilginis | Pathogen isolate | Senecio vulgaris | Reproductive biomass | Inglese & Paul, 2006 [76] | |
| Fungus | Puccinia lagenophorae | Pathogen isolate | Senecio vulgaris | Reproductive biomass | Inglese & Paul, 2006 [76] | |
| Oomycete | Hyaloperonospora arabidopsidis | Host genotype; Pathogen genotype | Arabidopsis thaliana | Seed production | Salvaudon et al., 2007 [77]; 2008 [78] | |
| Virus | Cucumber mosaic virus | Host genotype | Mimulus gutattus | Flower production | Carr et al., 2006 [89] | |
| Virus | Turnip mosaic virus | Host species; Pathogen genotype | Arabidopsis thaliana | Lifespan | Vijayan et al., 2017 [67] | |
| Bacteria | Pseudomoas syringae | Host genotype | Arabidopsis thaliana | Seed production | Kover & Schaal 2002 [97] | |
| Bacteria | Pseudomoas viridiflava | Host genotype; Pathogen isolate | Arabidopsis thaliana | Seed production % Mortality | Jakob et al., 2002 [96]; Goss & Bergelson, 2007 [70] | |
| Plant | Tristerix aphyllus | Infection status | Echinopsis chilensis | Branching | Medel 2001 [99] | |
| Plant | Cuscuta europea | Host family; Sex of host plant | Urtica dioica | Reproductive biomass | Koskela et al., 2002 [100] | |
| PT | Fungus | Erysiphe fischeri | Host genotype | Senecio vulgaris | Seed production | Ben-Kalio & Clarke, 1979 [74] |
| Fungus | Puccinia spp. | Host genotype; Pathogen genotype | Various hosts | Seed production Mortality | Summarized in: Roy & Kirchner, 2000 (Table 2) [16] | |
| Fungus | Uromyces spp. | Host genotype; Pathogen genotype | Various hosts | Seed production Mortality | Summarized in: Roy & Kirchner, 2000 (Table 2) [16] | |
| Oomycete | Hyaloperonospora arabidopsidis | Host family | Arabidopsis thaliana | Seed production | Salvaudon & Shykoff, 2013 [79] | |
| Virus | Cucumber mosaic virus | Host inbreeding level | Mimulus gutattus | Flower production | Carr et al., 2003 [58] | |
| Virus | Cucumber mosaic virus | Host genotype; Host allometric group; Host density; Pathogen isolate | Arabidopsis thaliana | Seed production | Pagán et al., 2007 [65]; 2008 [66]; 2009 [91] | |
| Cucumber mosaic virus | Host genotype; Host allometric group | Arabidopsis thaliana | Seed production Lifespan | Hily et al., 2016 [59] | ||
| Bacteria | Xhantomonas campestris | Host genotype | Arabidopsis thaliana | Chlorophyll content | Tsuji et al., 1991 [94] | |
| Bacteria | Xhantomonas campestris | Host genotype | Arabidopsis thaliana | Symptoms | Buell & Somerville, 1995 [95] | |
| Plant | Rhinanthus minor | Host genotype; Pathogen population | Hordeum vulgare | Seed production | Rowntree et al., 2011 [101] | |
| Rhinanthus angustifolius | Host genotype; Pathogen population | Hordeum vulgare | Seed production | Rowntree et al., 2011 [101] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pagán, I.; García-Arenal, F. Tolerance to Plant Pathogens: Theory and Experimental Evidence. Int. J. Mol. Sci. 2018, 19, 810. https://doi.org/10.3390/ijms19030810
Pagán I, García-Arenal F. Tolerance to Plant Pathogens: Theory and Experimental Evidence. International Journal of Molecular Sciences. 2018; 19(3):810. https://doi.org/10.3390/ijms19030810
Chicago/Turabian StylePagán, Israel, and Fernando García-Arenal. 2018. "Tolerance to Plant Pathogens: Theory and Experimental Evidence" International Journal of Molecular Sciences 19, no. 3: 810. https://doi.org/10.3390/ijms19030810
APA StylePagán, I., & García-Arenal, F. (2018). Tolerance to Plant Pathogens: Theory and Experimental Evidence. International Journal of Molecular Sciences, 19(3), 810. https://doi.org/10.3390/ijms19030810