Genome-Wide Identification and Characterization of Warming-Related Genes in Brassica rapa ssp. pekinensis

 , ,
, ,
Abstract
:1. Introduction
2. Results
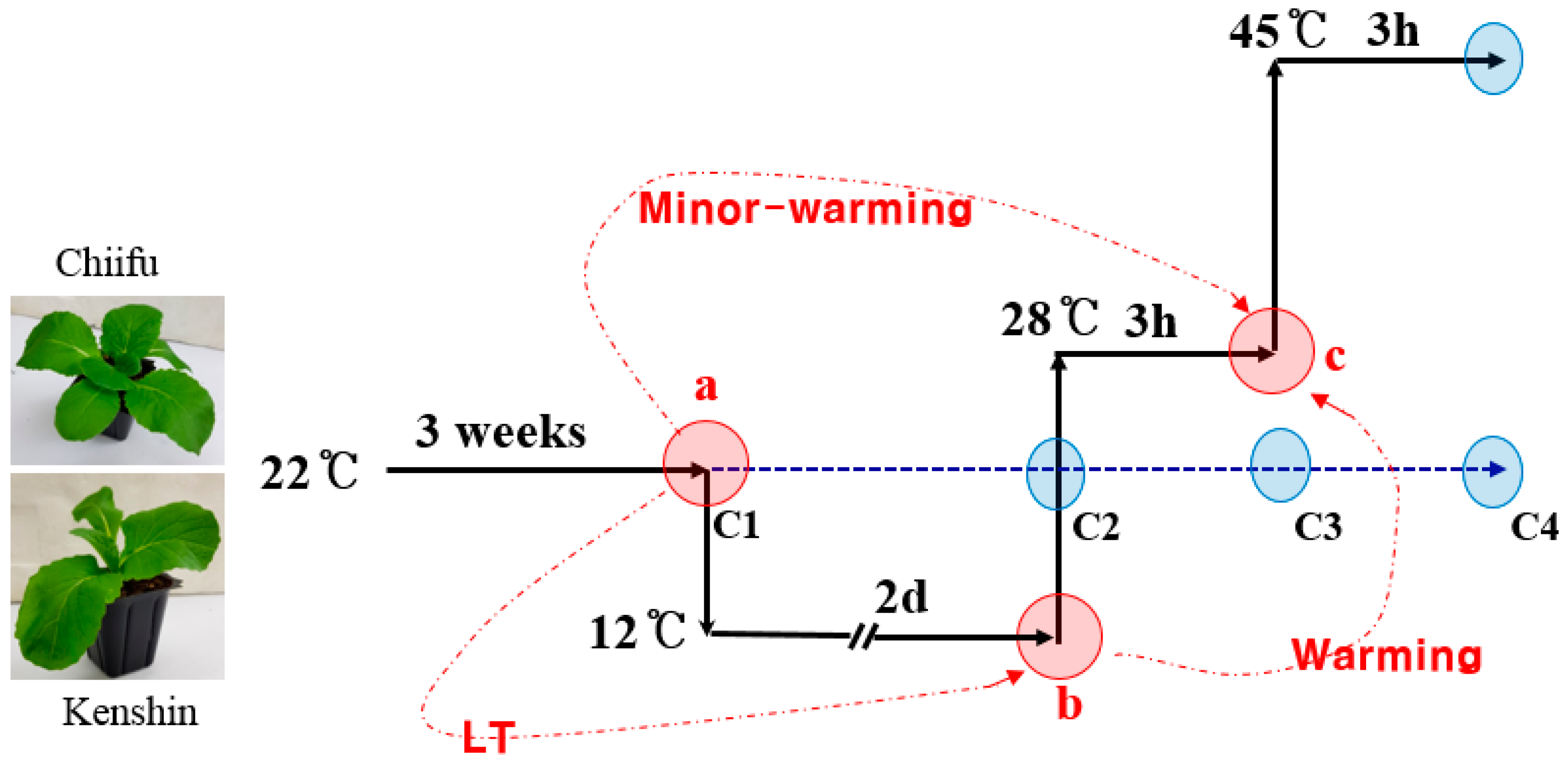
2.1. Transcriptome Analysis of Plants under Warming, Minor-Warming, and Low-Temperature (LT) Conditions Using the Br135K Microarray
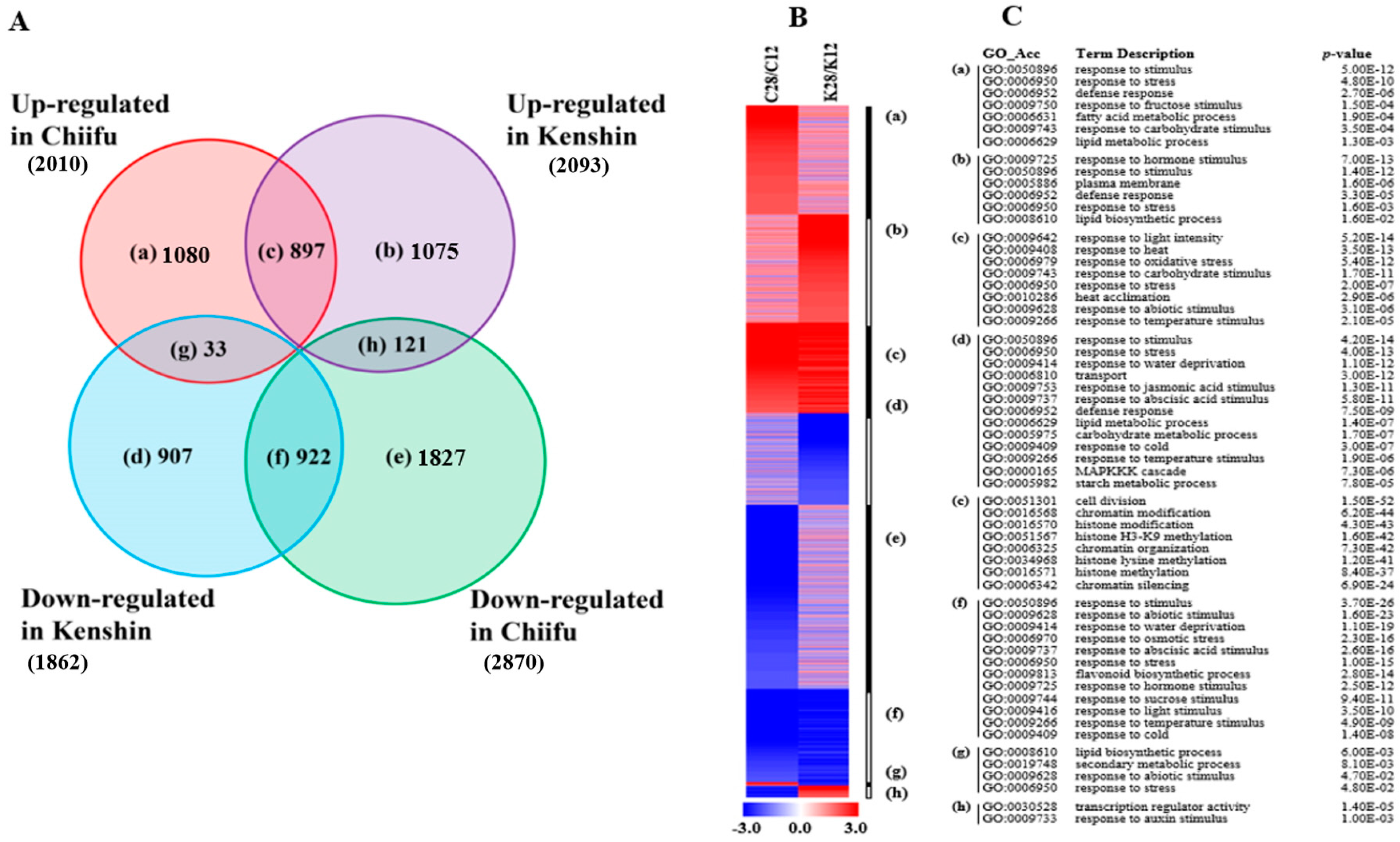
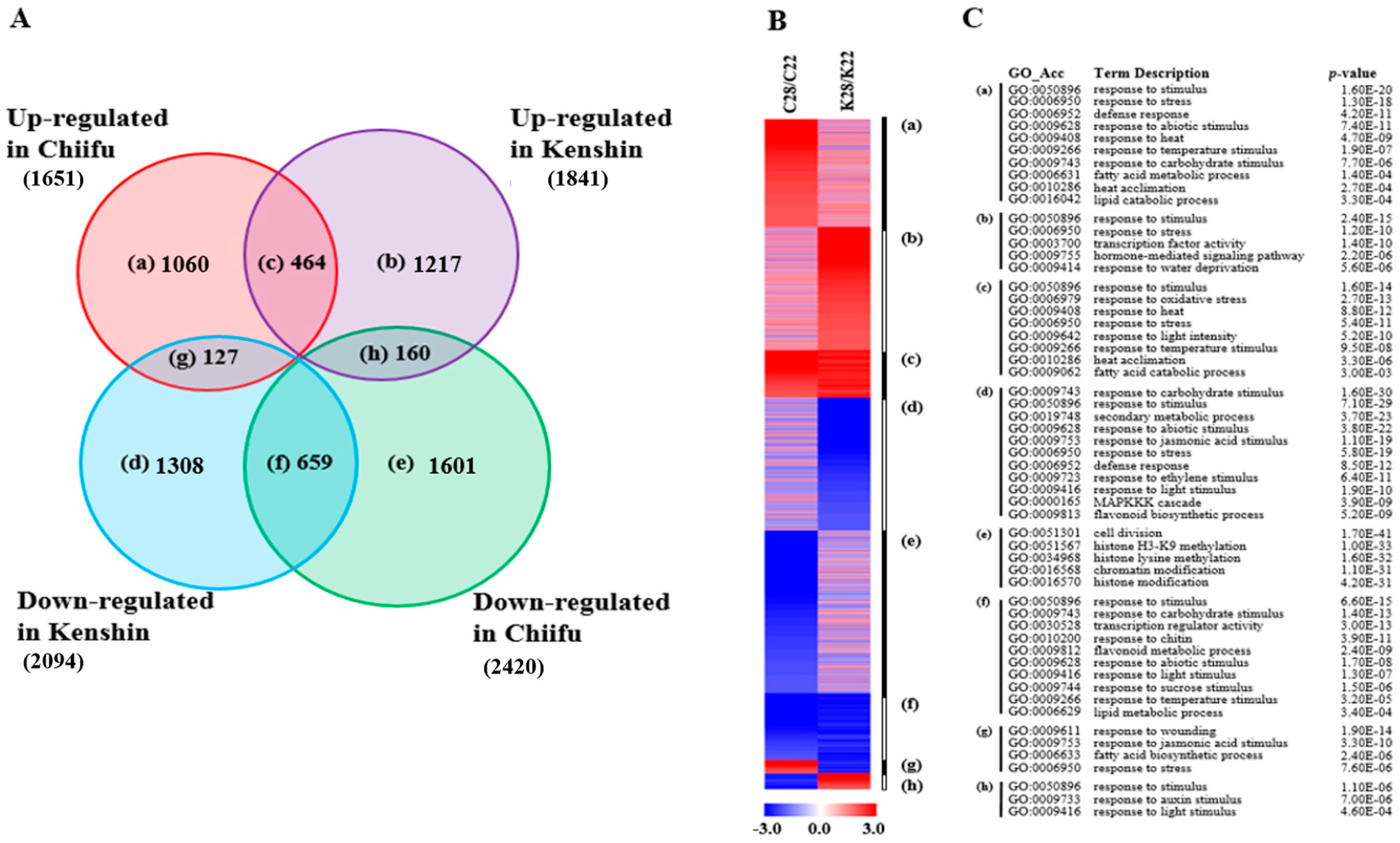
2.1.1. Warming-Responsive Genes
2.1.2. Minor Warming-Responsive Genes
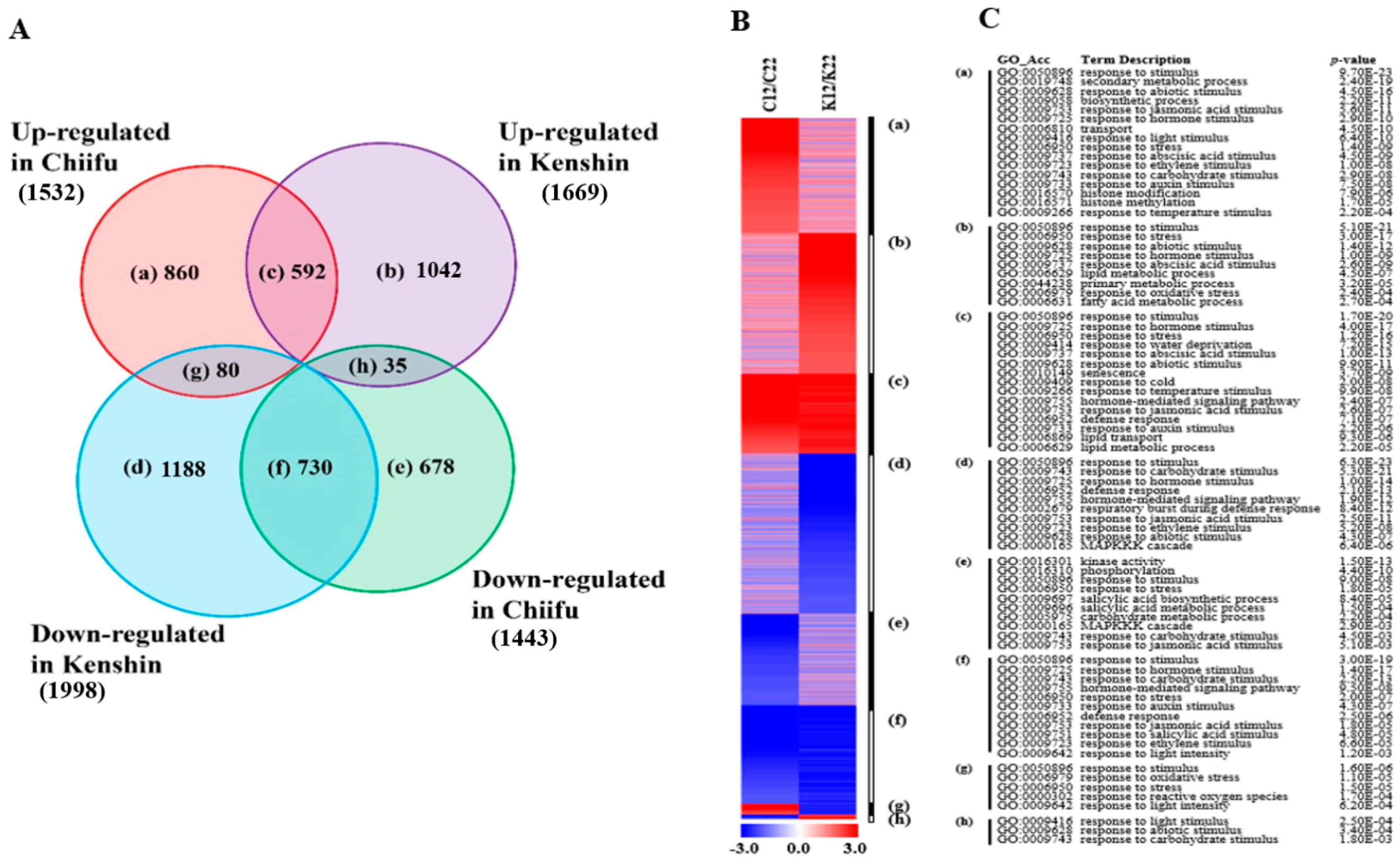
2.1.3. LT-Responsive Genes
2.1.4. Genes Upregulated by Both Warming and Minor-Warming Treatment
2.1.5. Genes Upregulated by Both LT and Warming Treatment
2.2. GO Analysis of Warming- and Minor Warming-Responsive Genes
2.3. Identification of Genes Associated with HT Adaptation in Kenshin
2.4. Comparison of HT-Related Gene Expression between B. rapa and Arabidopsis
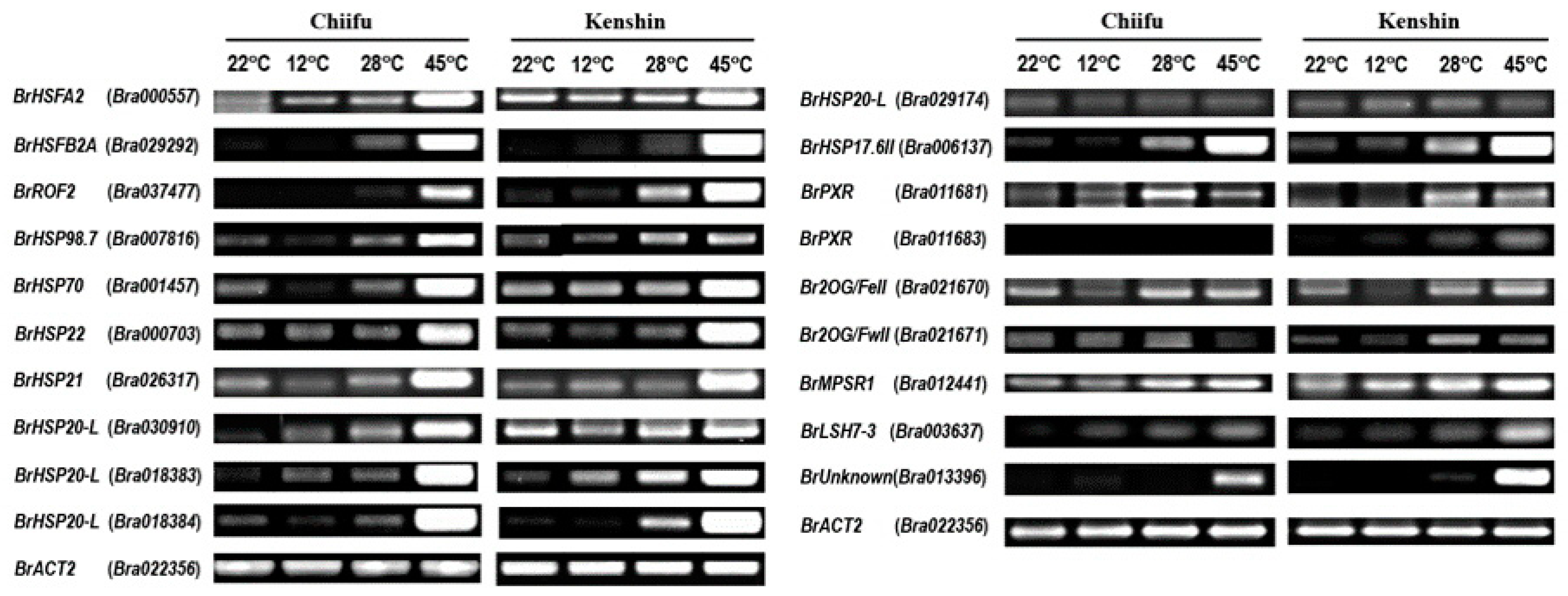
2.5. Confirmation of Microarray Data via qRT-PCR
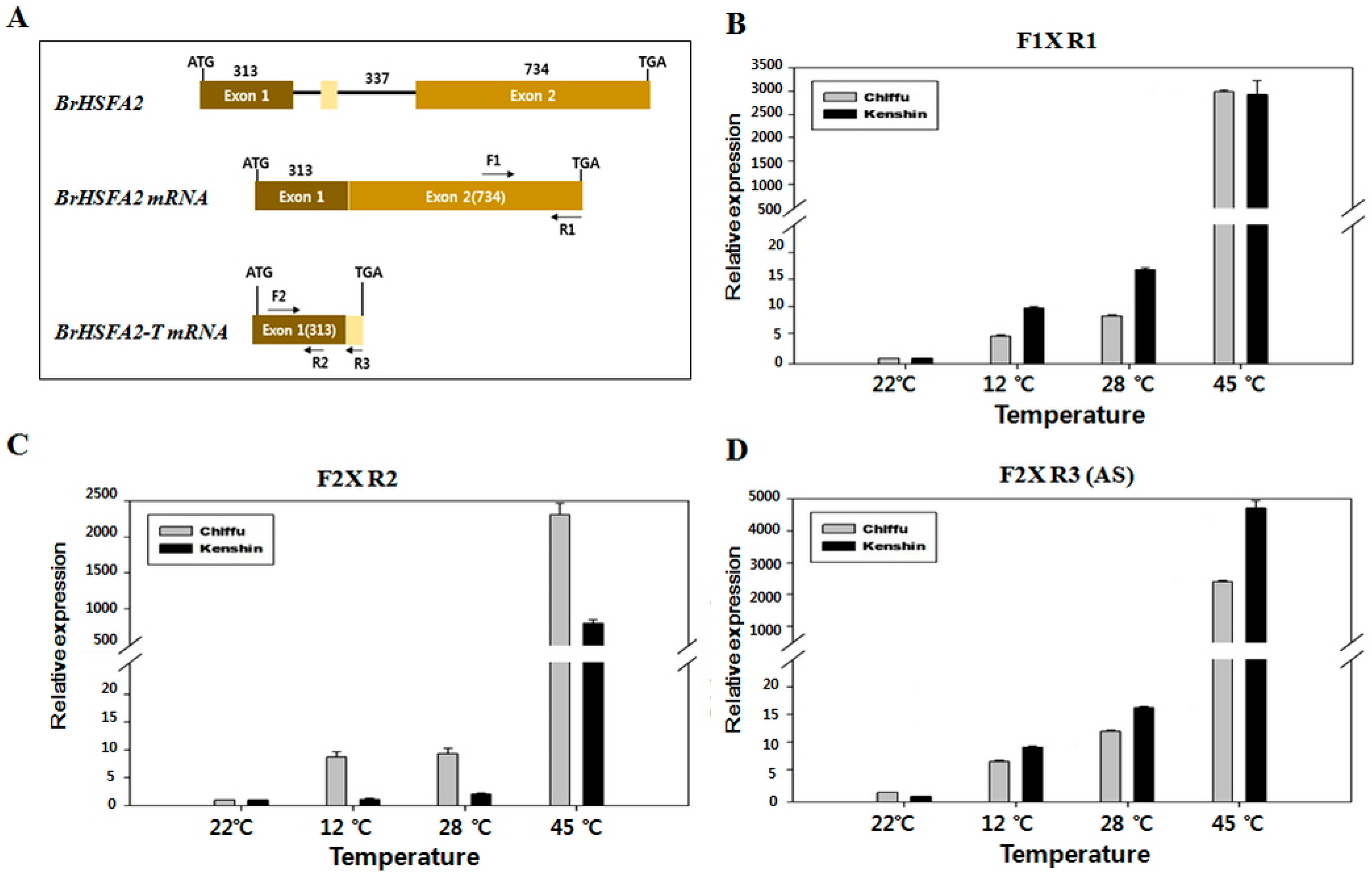
2.6. Expression of BrHSFA2 and BrHSP18.2
2.7. BrHSP18.2 Promoter Analysis
3. Discussion
3.1. Transcriptome Analysis
3.2. Functional Classification of Putative Warming Genes
3.3. Putative HT Adaptation-Related Genes in Kenshin
3.4. BrHSFA2 and Its Target BrHSP18.2s
3.5. BrHSP18.2s Promoters and Their Possible Control
4. Materials and Methods
4.1. Plant Materials
4.2. Br135K Microarray Analysis
4.3. RNA Extraction, RT-PCR, and qRT-PCR
4.4. Gene Cloning and Sequence Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Bita, C.E.; Gerats, T. Plant tolerance to high temperature in a changing environment, scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 2013, 4, 273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Driedonks, N.; Rieu, I.; Vrienzen, W.H. Breeding for plant heat tolerance at vegetative and reproductive stages. Plant Reprod. 2016, 29, 67–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fragkostefanakis, S.; Röth, S.; Schleiff, E.; Scharf, K.D. Prospects of engineering thermotolerance in crops through modulation of heat stress transcription factor and heat shock protein networks. Plant Cell Environ. 2015, 38, 1881–1895. [Google Scholar] [CrossRef] [PubMed]
- Kole, C.; Muthamilarasan, M.; Henry, R.; Edwards, D.; Sharma, R.; Abberton, M.; Batley, J.; Bentley, A.; Blakeney, M.; Bryant, J.; et al. Application of genomics-assisted breeding for generation of climate resilient crops, progress and prospects. Front. Plant Sci. 2015, 6, 563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larkindale, J.; Vierling, E. Core genome responses involved in acclimation to high temperature. Plant Physiol. 2008, 146, 748–761. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.V.; Wigge, P.A. H2A.Z-containing nucleosomes mediate the thermosensory response in Arabidopsis. Cell 2010, 140, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Finka, A.; Goloubinoff, P. How do plants fee the heat? Trends Biochem. Sci. 2012, 37, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.H.; Kaplinsky, N.J.; Hu, C.; Charng, Y.Y. Some like it hot, some like it warm, phenotyping to explore thermotolerance diversity. Plant Sci. 2012, 195, 10–23. [Google Scholar] [CrossRef] [PubMed]
- Koini, M.A.; Alvey, L.; Allen, T.; Tilley, C.A.; Harberd, N.P.; Whitelam, G.C.; Franklin, K.A. High temperature-mediated adaptations in plant architecture require the bHLH transcription factor PIF4. Curr. Biol. 2009, 19, 408–413. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.V.; Lucyshyn, D.; Jaeger, K.E.; Alos, E.; Alvey, E.; Harberd, N.P.; Wigge, P.A. Transcription factor PIF4 controls the thermosensory activation of flowering. Nature 2012, 484, 242–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quint, M.; Delker, C.; Franklin, K.A.; Wigge, P.A.; Halliday, K.J.; van Zanten, M. Molecular and genetic control of plant thermomorphogenesis. Nat. Plants 2016, 2, 15190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proveniers, M.C.; van Zanten, M. High temperature acclimation through PIF4 signaling. Trends Plant Sci. 2013, 18, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.H.; Domijan, M.; Klose, C.; Biswas, S.; Ezer, D.; Gao, M.; Khattak, A.K.; Box, M.S.; Charoensawan, V.; Cortijo, S.; et al. Phytochromes function as thermosensors in Arabidopsis. Science 2016, 354, 886–889. [Google Scholar] [CrossRef] [PubMed]
- Verhage, L.; Severing, E.I.; Bucher, J.; Lammers, M.; Busscher-Lange, J.; Bonnema, G.; Rodenburg, N.; Proveniers, M.C.; Angenent, G.C.; Immink, R.G. Splicing-related genes are alternatively spliced upon changes in ambient temperatures in plants. PLoS ONE 2017, 12, e0172950. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.M.; Inden, H.; Asahira, T. Interspecific hybrids between Brassica campestris L. and B. oleracea L. through embryo and ovary culture. Mem. Coll. Agric. Kyoto Univ. 1989, 135, 21–30. [Google Scholar]
- Yamagishi, H.; Hossain, M.M.; Yonezawa, K. Morphology, fertility and cross-compatibility of somatic hybrids between Brassica oleracea L. and B. campestris L. Sci. Horticult. 1994, 58, 283–288. [Google Scholar] [CrossRef]
- Dong, X.; Yi, H.; Lee, J.; Nou, I.S.; Han, C.T.; Hur, Y. Global gene-expression analysis to identify differentially expressed genes critical for the heat stress response in Brassica rapa. PLoS ONE 2015, 10, e0130451. [Google Scholar] [CrossRef] [PubMed]
- Chinnusamy, V.; Zhu, J.; Zhu, J.K. Cold stress regulation of gene expression in plants. Trends Plant Sci. 2007, 12, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Scharf, K.D.; Berberich, T.; Ebersberger, I.; Nover, L. The plant heat stress transcription factor (Hsf) family, structure, function and evolution. Biochim. Biophys. Acta 2012, 1819, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Abiotic stress signaling and response in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Li, Y.; Daun, W.; Huang, F.; Hou, X. Cold acclimation alters DNA methylation patterns and confers tolerance to heat and increase growth rate in Brassica rapa. J. Exp. Bot. 2017, 68, 1213–1224. [Google Scholar] [CrossRef] [PubMed]
- Stief, A.; Altmann, S.; Hoffmann, K.; Pant, B.D.; Scheible, W.R.; Bäurle, I. Arabidopsis miR156 regulates tolerance to recurring environmental stress through SPL transcription factors. Plant Cell 2014, 26, 1792–1807. [Google Scholar] [CrossRef] [PubMed]
- Lämke, J.; Brzezinka, K.; Bäurle, I. HSFA2 orchestrates transcriptional dynamics after heat stress in Arabidopsis thaliana. Transcription 2016, 7, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Lämke, J.; Brzezinka, K.; Altmann, S.; Bäurle, I. A hit-and-run heat shock factor governs sustained histone methylation and transcriptional stress memory. EMBO J. 2016, 35, 162–175. [Google Scholar] [CrossRef] [PubMed]
- Perisic, O.; Xiao, H.; Lis, J.T. Stable binding of Drosophia heat shock factor to head-to-head and tail-to-tail repeats of conserved 5 bp recongnition unit. Cell 1989, 59, 797–806. [Google Scholar] [CrossRef]
- Nishizawa-Yokoi, A.; Yoshida, E.; Yabuta, Y.; Shigeoka, S. Analysis of the regulation of target genes by an Arabidopsis heat shock transcription factor, HsfA2. Biosci. Biotechnol. Biochem. 2009, 73, 890–895. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef] [PubMed]
- Oshino, T.; Miura, S.; Kikuchi, S.; Hamada, K.; Yano, K.; Watanabe, M.; Higashitani, A. Auxin depletion in barley plants under high-temperature conditions represses DNA proliferation in organelles and nuclei via transcriptional alterations. Plant Cell Environ. 2011, 34, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Li, J.; Hao, R.; Guo, Y. Activation of catalase activity by a peroxisome-localized small heat shock protein Hsp17.6CII. J. Genet. Genom. 2017, 44, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, E.; Mano, S.; Hara-Nishimura, I.; Nishimura, M.; Yamada, K. HSP90 stabilizes auxin receptor TIR1 and ensures plasticity of auxin responses. Plant Signal. Behav. 2017, 12, e1311439. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, A.; Yabuta, Y.; Yoshida, E.; Maruta, T.; Yoshimura, K.; Shigeoka, S. Arabidopsis heat shock transcription factor A2 as a key regulator in response to several types of environmental stress. Plant J. 2006, 48, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Charng, Y.Y.; Liu, H.C.; Liu, N.Y.; Chi, W.T.; Wang, C.N.; Chang, S.H.; Wang, T.T. A heat-inducible transcription factor, HsfA2, is required for extension of acquired thermotolerance in Arabidopsis. Plant Physiol. 2007, 143, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.; Mittler, R. Could heat shock transcription factors function as hydrogen peroxide sensors in plants? Ann. Bot. 2006, 98, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, D.; Yamaguchi, K.; Nishiuchi, T. High-level overexpression of the Arabidopsis HsfA2 gene confers not only increased themotolerance but also salt/osmotic stress tolerance and enhanced callus growth. J. Exp. Bot. 2007, 58, 3373–3383. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.J.; Yang, K.A.; Hong, J.K.; Choi, J.S.; Yun, D.J.; Hong, J.C.; Chung, W.S.; Lee, S.Y.; Cho, M.J.; Lim, C.O. Gene expression profiles during heat acclimation in Arabidopsis thaliana suspension-culture cell. J. Plant Res. 2006, 119, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Lin, S.Y.; Chi, W.T.; Charng, Y.Y. Recent gene duplication and subfunctionalization produced a mitochondrial GrpE, the nucleotide exchange factor of the Hsp70 complex, specialized in thermotolerance to chronic heat stress in Arabidopsis. Plant Physiol. 2012, 158, 747–758. [Google Scholar] [CrossRef] [PubMed]
- Clay, N.K.; Nelson, T. The recessive epigenetic swellmap mutation affects the expression of two stop II splicing factors required for the transcription of the cell proliferation gene STRUWWELPETER and for the the timing of cell cycle arrest in the Arabidopsis leaf. Plant Cell 2005, 17, 1994–2008. [Google Scholar] [CrossRef] [PubMed]
- Lázaro, A.; Gómez-Zambrano, A.; López-González, L.; Piñeiro, M.; Jarillo, J.A. Mutations in the Arabidopsis SWC6 gene, encoding a component of the SWR1 chromatin remodelling complex, accelerate flowering time and alter leaf and flower development. J. Exp. Bot. 2008, 59, 653–666. [Google Scholar] [CrossRef] [PubMed]
- Nomoto, Y.; Kubozono, S.; Miyachi, M.; Yamashino, T.; Nakamichi, N.; Mizuno, T. A circadian clock- and PIF4-mediated double coincidence mechanism is implicated in the thermosensitive photoperiodic control of plant architectures in Arabidopsis thaliana. Plant Cell Physiol. 2012, 53, 1965–1973. [Google Scholar] [CrossRef] [PubMed]
- Nomoto, Y.; Kubozono, S.; Miyachi, M.; Yamashino, T.; Nakamichi, N.; Mizuno, T. Circadian clock and PIF4-mediated external coincidence mechanism coordinately integrates both of the cues from seasonal changes in photoperiod and temperature to regulate plant growth in Arabidopsis thaliana. Plant Signal. Behav. 2013, 8, e22863. [Google Scholar] [CrossRef] [PubMed]
- Franklin, K.A.; Lee, S.H.; Patel, D.; Kumar, S.V.; Spartz, A.K.; Gu, C.; Ye, S.; Yu, P.; Breen, G.; Cohen, J.D.; et al. Phytochrome-interacting factor 4 (PIF4) regulates auxin biosynthesis at high temperature. Proc. Natl. Acad. Sci. USA 2011, 108, 20231–20235. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Qi, L.; Li, Y.; Chu, J.; Li, C. PIF4-mediated activation of YUCCA8 expression integrates temperature into the auxin pathway in regulating Arabidopsis hypocotyl growth. PLoS Genet. 2012, 8, e1002594. [Google Scholar] [CrossRef] [PubMed]
- Stavang, J.A.; Gallego-Bartolomé, J.; Gómez, M.D.; Yoshida, S.; Asami, T.; Olsen, J.E.; García-Martínez, J.L.; Alabadí, D.; Blázquez, M.A. Hormonal regulation of temperature-induced growth in Arabidopsis. Plant J. 2009, 60, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Oh, E.; Zhu, J.Y.; Wang, Z.Y. Interaction between BZR1 and PIF4 integrates brassinosteroid and environmental responses. Nat. Cell Biol. 2012, 14, 802–809. [Google Scholar] [CrossRef] [PubMed]
- Bernfur, K.; Rutsdottir, G.; Emanuelsson, C. The chloroplast-localized small heat shock protein Hsp21 associates with the thylakoid membranes in heat-stressed plants. Protein Sci. 2017, 26, 1773–1784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shigeto, J.; Tsutsumi, Y. Diverse functions and reactions of class III peroxidases. New Phytol. 2015, 209, 1395–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, S.; Fartyal, D.; Agarwal, A.; Shukla, T.; James, D.; Kaul, T.; Negi, Y.K.; Arora, S.; Reddy, M.K. Abiotic stress tolerance in plants: Myriad roles of ascorbate peroxidase. Front. Plant Sci. 2017, 8, 581. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Cho, S.K.; Oh, T.R.; Ryu, M.Y.; Yang, S.W.; Kim, W.T. MPSR1 is a cytoplasmic PQC E3 ligase for eliminating emergent misfolded proteins in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2017, 114, E10009. [Google Scholar] [CrossRef] [PubMed]
- Charng, Y.Y.; Liu, H.C.; Liu, N.Y.; Hsu, F.C.; Ko, S.S. Arabidopsis Hsa32, a novel heat shock protein, is essential for acquired thermotolerance during long recovery after acclimation. Plant Physiol. 2006, 140, 1297–1305. [Google Scholar] [CrossRef] [PubMed]
- Meiri, D.; Breiman, A. Arabidopsis ROF1 (FKBP62) modulates thermotolerance by interacting with HSP90.1 and affecting the accumulation of HsfA2-regulated sHSPs. Plant J. 2009, 59, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Lämke, J.; Bäurle, I. Epigenetic and chromatin-based mechanisms in environmental stress adaptation and stress memory in plants. Genome Biol. 2017, 18, 124. [Google Scholar] [CrossRef] [PubMed]
- Sugio, A.; Dreos, R.; Apricio, F.; Maule, A.J. The cytosolic protein response as a subcomponent of the wider heat shock response in Arabidopsis. Plant Cell 2009, 21, 642–654. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhuang, L.; Shi, Y.; Huang, B. Up-regulation of HSFA2c and HSPs by ABA contributing to improved heat tolerance in tall fescue and Arabidopsis. Int. J. Mol. Sci. 2017, 18, 1981. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Sun, N.; Liu, M.; Liu, J.; Du, B.; Wang, X.; Qi, X. An autoregulatory loop controlling Arabidopsis HsfA2 expression, role of heat shock-induced alternative splicing. Plant Physiol. 2013, 162, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Naito, S.; Komeda, Y. The Arabidopsis HSP18.2promoter/GUS gene fusion in transgenic Arabidopsis plants, a powerful tool for the isolation of regulatory mutants of the heat-shock response. Plant J. 1992, 2, 751–761. [Google Scholar] [CrossRef]
- Jung, H.J.; Dong, X.; Park, J.I.; Thamilarasan, S.K.; Lee, S.S.; Kim, Y.K.; Lim, Y.P.; Nou, I.S.; Hur, Y. Genome-wide transcriptome analysis of two contrasting Brassica rapa doubled haploid lines under cold-stresses using Br135K oligomeric chip. PLoS ONE 2014, 9, e106069. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology, tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Zhou, X.; Ling, Y.; Zhang, Z.; Su, Z. agriGO, A GO analysis toolkit for the agricultural community. Nucleic Acids Res. 2010, 38, W64–W70. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inbred Line (Treatment) | Comparison | No. of Genes (Table S5) | SEGs (Tables S7–S12) | Comparison | No. of Genes (Table S6) |
|---|---|---|---|---|---|
| Chiifu (22 °C ⟶ 12 °C ⟶ 28 °C) | 28 °C/22 °C | 1651 | 121 | 12 °C/22 °C | 1532 |
| 28 °C/12 °C | 2010 | 49 | 28 °C/12 °C | 2010 | |
| Both | 759 | 85 | Both | 40 | |
| Kenshin (22 °C ⟶ 12 °C ⟶ 28 °C) | 28 °C/22 °C | 1841 | 193 | 12 °C/22 °C | 1669 |
| 28 °C/12 °C | 2093 | 146 | 28 °C/12 °C | 2093 | |
| Both | 726 | 86 | Both | 59 |
| Classification | Kenshin (W) | Kenshin (W + MW) | Kenshin/Chiifu (W + MW) |
|---|---|---|---|
| Heat acclimation | - | 1 | 6 |
| Response to heat | - | - | 13 |
| Response to stress | 8 | 6 | 8 |
| Transcription factor activity | 9 | 9 | 9 |
| Transferase activity | 9 | 5 | 12 |
| Transport | 7 | 6 | 6 |
| Carbohydrate metabolic process | 8 | 4 | 4 |
| Ligase activity | - | 5 | 1 |
| Lipid biosynthetic process | 14 | 4 | 3 |
| Oxidation reduction | - | 1 | 2 |
| Response to auxin stimulus | - | 3 | 1 |
| Response to oxidative stress | - | 3 | 2 |
| Response to salicylic acid stimulus | - | 3 | 3 |
| Chromosome organization | - | - | 5 |
| Response to hormone stimulus | 22 | - | - |
| Intracellular membrane bound organelle | 23 | - | - |
| Signal transduction | 6 | - | - |
| Primary metabolic process | 7 | - | - |
| Catalytic activity | 7 | - | - |
| Plasma membrane | 3 | - | - |
| Ion binding | 5 | - | - |
| Unclassified | 4 | 15 | 34 |
| Unknown protein | 6 | 5 | 14 |
| Not annotated | 8 | 16 | 34 |
| Total | 132 | 86 | 157 |
| Classification | At_Locus | Gene Description | Br_SEQ_ID | Expression Level (Probe Intensity) | Fold Change | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chiifu (C) | Kenshin (K) | Intrinsic | Warming | Warming | Minor-W | ||||||||
| 22 °C | 12 °C | 28 °C | 22 °C | 12 °C | 28 °C | K22/C22 | K28/K12 | C28/C12 | K28/K22 | ||||
| HSR | AT2G26150 | Heat shock transcription factor A2 (HSFA2) | Bra000557 * | 68 | 223 | 1256 | 256 | 542 | 1786 | 3.8 | 3.3 | 5.6 | 7.0 |
| AT5G62020 | Heat shock transcription factor B2A (HSFB2A) | Bra029292 * | 385 | 374 | 3164 | 875 | 587 | 3814 | 2.3 | 6.5 | 8.5 | 4.4 | |
| AT4G25200 | Mitochondrion-localized small heat shock protein 23.6 (HSP23.6-MITO) | Bra013872 * | 333 | 323 | 1308 | 984 | 265 | 3551 | 3.0 | 13.4 | 4.0 | 3.6 | |
| AT4G10250 | HSP20-like chaperones superfamily protein (HSP22.0) | Bra027999 * | 27 | 71 | 378 | 85 | 37 | 677 | 3.2 | 18.1 | 5.3 | 7.9 | |
| AT1G54050 | HSP20-like chaperones superfamily protein | Bra030910 * | 827 | 945 | 4041 | 2406 | 2649 | 9204 | 2.9 | 3.5 | 4.3 | 3.8 | |
| AT2G29500 | HSP20-like chaperones superfamily protein | Bra018383 * | 1870 | 2774 | 11,166 | 5211 | 2428 | 19,026 | 2.8 | 7.8 | 4.0 | 3.7 | |
| Bra018384 | 2575 | 5095 | 6789 | 5906 | 5362 | 14,692 | 2.3 | 2.7 | 1.3 | 2.5 | |||
| Bra031725 | 1996 | 4409 | 5909 | 6292 | 5275 | 13,450 | 3.2 | 2.5 | 1.3 | 2.1 | |||
| AT5G51440 | HSP20-like chaperones superfamily protein | Bra029174 * | 406 | 376 | 5772 | 1657 | 292 | 7627 | 4.1 | 26.1 | 15.3 | 4.6 | |
| AT5G59720 | Heat shock protein 18.2 (HSP18.2) | Bra002539 * | 73 | 259 | 747 | 206 | 58 | 5114 | 2.8 | 88.5 | 2.9 | 24.8 | |
| AT5G12020 | 17.6 kDa class II heat shock protein (HSP17.6II) | Bra006137 * | 52 | 271 | 2513 | 909 | 310 | 2036 | 17.6 | 6.6 | 9.3 | 2.2 | |
| AT5G48570 | FKBP-type peptidyl-prolyl cis-trans isomerase family protein (ROF2) | Bra037477 * | 309 | 283 | 4967 | 1632 | 352 | 6607 | 5.3 | 18.8 | 17.6 | 4.0 | |
| AT1G72660 | P-loop containing nucleoside triphosphate hydrolases superfamily protein | Bra016043 * | 212 | 185 | 2034 | 967 | 889 | 2723 | 4.6 | 3.1 | 11.0 | 2.8 | |
| AT5G47830 | Unknown protein | Bra020728 * | 523 | 544 | 4910 | 1310 | 1076 | 4854 | 2.5 | 4.5 | 9.0 | 3.7 | |
| AT3G14200 | Chaperone DnaJ-domain superfamily protein | Bra027363 * | 1333 | 2129 | 5365 | 2857 | 2634 | 7477 | 2.1 | 2.8 | 2.5 | 2.6 | |
| TF | AT2G23690 | HTH-type transcriptional regulator | Bra039208 | 1122 | 1383 | 3449 | 2567 | 1072 | 6076 | 2.3 | 5.7 | 2.5 | 2.4 |
| AT5G56840 | MYB-like transcription factor family protein | Bra002790 * | 89 | 154 | 589 | 251 | 144 | 767 | 2.8 | 5.3 | 3.8 | 3.0 | |
| AT5G52600 | MYB domain protein 82 (MYB82) | Bra029113 | 64 | 334 | 132 | 205 | 371 | 1579 | 3.2 | 4.3 | 0.4 | 7.7 | |
| AT2G24645 | Transcriptional factor B3 family protein | Bra032079 | 43 | 85 | 77 | 182 | 244 | 1013 | 4.3 | 4.2 | 0.9 | 5.6 | |
| AT1G70270 | Transcription factor | Bra007905 | 246 | 797 | 368 | 1127 | 952 | 3529 | 4.6 | 3.7 | 0.5 | 3.1 | |
| AT5G15150 | Homeobox 3 (HB3) | Bra023506 | 197 | 389 | 165 | 389 | 372 | 1184 | 2.0 | 3.2 | 0.4 | 3.0 | |
| AT5G66940 | Dof-type zinc finger DNA-binding family protein | Bra012119 | 488 | 1459 | 209 | 1159 | 983 | 2746 | 2.4 | 2.8 | 0.1 | 2.4 | |
| AT3G62090 | Phytochrome interacting factor 3-like 2 (PIL2/PIF6) | Bra007660 * | 218 | 239 | 537 | 426 | 684 | 1883 | 2.0 | 2.8 | 2.2 | 4.4 | |
| AT5G10970 | C2H2 and C2HC zinc fingers superfamily protein | Bra009000 | 303 | 776 | 668 | 823 | 878 | 2180 | 2.7 | 2.5 | 0.9 | 2.6 | |
| AT1G23380 | KNOTTED1-like homeobox gene 6 (KNAT6) | Bra016348 | 140 | 970 | 475 | 273 | 1004 | 2202 | 2.0 | 2.2 | 0.5 | 8.1 | |
| AT4G18610 | Light-dependent short hypocotyl 9 (LSH9) | Bra021000 | 83 | 85 | 77 | 189 | 112 | 605 | 2.3 | 5.4 | 0.9 | 3.2 | |
| AT2G42610 | Light-dependent short hypocotyl 10 (LSH10) | Bra016865 | 422 | 617 | 197 | 1008 | 1013 | 2079 | 2.4 | 2.1 | 0.3 | 2.1 | |
| SF | AT1G65660 | Pre-mRNA splicing Prp18-interacting factor (SMP1) | Bra023741 | 141 | 232 | 1190 | 664 | 589 | 1712 | 4.7 | 2.9 | 5.1 | 2.6 |
| Others | AT4G36430 | Peroxidase superfamily protein | Bra017761 | 125 | 381 | 182 | 543 | 686 | 2278 | 4.4 | 3.3 | 0.5 | 4.2 |
| AT1G16530 | ASYMMETRIC LEAVES 2-like 9 (LBD3/ASL9) | Bra026042 | 171 | 393 | 272 | 522 | 824 | 1729 | 3.1 | 2.1 | 0.7 | 3.3 | |
| Bra026716 | 209 | 325 | 291 | 454 | 534 | 1063 | 2.2 | 2.0 | 0.9 | 2.3 | |||
| AT5G59670 | Leucine-rich repeat protein kinase family protein | Bra020300 | 129 | 93 | 80 | 288 | 147 | 622 | 2.2 | 4.2 | 0.9 | 2.2 | |
| AT4G19530 | Disease resistance protein (TIR-NBS-LRR class) family | Bra027594 | 158 | 206 | 202 | 532 | 536 | 1591 | 3.4 | 3.0 | 1.0 | 3.0 | |
| AT2G32660 | Receptor like protein 22 (RLP22) | Bra021803 | 163 | 72 | 201 | 960 | 963 | 2013 | 5.9 | 2.1 | 2.8 | 2.1 | |
| AT1G51860 | Leucine-rich repeat protein kinase family protein | Bra030411 | 92 | 75 | 41 | 874 | 1051 | 1992 | 9.5 | 1.9 | 0.5 | 2.3 | |
| AT1G53350 | Disease resistance protein (CC-NBS-LRR class) family | Bra037453 * | 185 | 155 | 420 | 539 | 706 | 1425 | 2.9 | 2.0 | 2.7 | 2.6 | |
| AT4G08570 | Heavy metal transport/detoxification superfamily protein | Bra037865 | 37 | 45 | 505 | 255 | 67 | 2044 | 6.9 | 30.5 | 11.1 | 8.0 | |
| AT5G66110 | Heavy metal transport/detoxification superfamily protein (HIPP27) | Bra009662 | 115 | 329 | 449 | 1102 | 1195 | 7086 | 9.5 | 5.9 | 1.4 | 6.4 | |
| AT1G79360 | Organic cation/carnitine transporter 2 (OCT2) | Bra035111 | 53 | 37 | 23 | 135 | 111 | 568 | 2.5 | 5.1 | 0.6 | 4.2 | |
| AT2G04100 | MATE efflux family protein | Bra015133 | 78 | 181 | 255 | 350 | 510 | 1175 | 4.5 | 2.3 | 1.4 | 3.4 | |
| AT2G35460 | Late embryogenesis abundant (LEA) hydroxyproline-rich glycoprotein family | Bra028562 | 215 | 78 | 879 | 575 | 123 | 2595 | 2.7 | 21.1 | 11.3 | 4.5 | |
| AT2G25450 | 2-oxoglutarate (2OG) and Fe(II)-dependent oxygenase superfamily protein | Bra021671 | 246 | 68 | 240 | 530 | 644 | 5146 | 2.2 | 8.0 | 3.5 | 9.7 | |
| AT1G28030 | 2-oxoglutarate (2OG) and Fe(II)-dependent oxygenase superfamily protein | Bra021552 | 59 | 73 | 121 | 129 | 169 | 1079 | 2.2 | 6.4 | 1.7 | 8.4 | |
| AT2G39310 | Jacalin-related lectin 22 (JAL22) | Bra005053 | 16 | 99 | 45 | 36 | 109 | 826 | 2.3 | 7.6 | 0.5 | 23.1 | |
| AT3G16900 | LURP-one-like protein | Bra021211 | 217 | 205 | 358 | 1035 | 524 | 2674 | 4.8 | 5.1 | 1.7 | 2.6 | |
| AT4G36380 | Cytochrome P450 superfamily protein (ROT3) | Bra011678 * | 52 | 139 | 350 | 395 | 205 | 842 | 7.6 | 4.1 | 2.5 | 2.1 | |
| AT4G24110 | NADP-specific glutamate dehydrogenase | Bra013763 | 92 | 79 | 129 | 204 | 176 | 637 | 2.2 | 3.6 | 1.6 | 3.1 | |
| AT3G51000 | Alpha/beta-Hydrolases superfamily protein | Bra036841 | 69 | 21 | 23 | 676 | 873 | 3074 | 9.8 | 3.5 | 1.1 | 4.5 | |
| AT4G22460 | Bifunctional inhibitor/lipid-transfer protein/seed storage 2S albumin superfamily protein | Bra013619 | 152 | 251 | 304 | 541 | 555 | 1932 | 3.6 | 3.5 | 1.2 | 3.6 | |
| AT4G13410 | Nucleotide-diphospho-sugar transferases superfamily protein (CSLA15) | Bra008638 | 276 | 385 | 600 | 656 | 523 | 1384 | 2.4 | 2.6 | 1.6 | 2.1 | |
| AT5G40650 | Succinate dehydrogenase 2-2 (SDH2-2) | Bra028469 | 468 | 531 | 545 | 2517 | 2395 | 6064 | 5.4 | 2.5 | 1.0 | 2.4 | |
| AT5G05390 | Laccase 12 (LAC12) | Bra009111 | 279 | 1072 | 1201 | 1118 | 1176 | 2906 | 4.0 | 2.5 | 1.1 | 2.6 | |
| AT3G17820 | Glutamine synthetase 1.3 (GLN1.3) | Bra021276 * | 1197 | 1053 | 2579 | 2798 | 3121 | 7326 | 2.3 | 2.3 | 2.4 | 2.6 | |
| AT5G61260 | Plant calmodulin-binding protein-related | Bra029324 | 256 | 194 | 163 | 811 | 1078 | 2480 | 3.2 | 2.3 | 0.8 | 3.1 | |
| AT5G64870 | SPFH/Band 7/PHB domain-containing membrane-associated protein family | Bra024333 | 270 | 569 | 701 | 559 | 484 | 1113 | 2.1 | 2.3 | 1.2 | 2.0 | |
| AT1G20575 | Nucleotide-diphospho-sugar transferases superfamily protein | Bra025828 | 1293 | 888 | 2710 | 3467 | 3248 | 7310 | 2.7 | 2.3 | 3.1 | 2.1 | |
| AT4G39140 | RING/U-box superfamily protein | Bra025860 | 27 | 27 | 59 | 2422 | 2404 | 5394 | 88.5 | 2.2 | 2.2 | 2.2 | |
| AT3G09260 | Glycosyl hydrolase superfamily protein (BGLU23) | Bra034060 | 278 | 412 | 393 | 1034 | 995 | 2106 | 3.7 | 2.1 | 1.0 | 2.0 | |
| AT3G06550 | O-acetyltransferase family protein | Bra040276 | 250 | 250 | 310 | 492 | 762 | 1570 | 2.0 | 2.1 | 1.2 | 3.2 | |
| AT1G29590 | Eukaryotic initiation factor 4E protein (eIF4E3) | Bra032325 | 299 | 445 | 486 | 1304 | 1365 | 2571 | 4.4 | 1.9 | 1.1 | 2.0 | |
| AT4G19430 | Unknown protein | Bra013396 | 103 | 16 | 42 | 252 | 17 | 726 | 2.5 | 41.6 | 2.7 | 2.9 | |
| NA | NA | Bra012220 | 183 | 112 | 97 | 5190 | 4062 | 11,440 | 28.4 | 2.8 | 0.9 | 2.2 | |
| NA | NA | Bra025861 | 53 | 59 | 147 | 1979 | 1928 | 4522 | 37.3 | 2.3 | 2.5 | 2.3 | |
| NA | NA | Bra010352 | 596 | 604 | 1785 | 4135 | 4187 | 9370 | 6.9 | 2.2 | 3.0 | 2.3 | |
| Marker | At_Locus | Gene Description | Br_SEQ_ID | Expression Level (Probe Intensity) | Fold Change | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chiifu | Kenshin | Chiifu | Kenshin | ||||||||||
| 22 °C | 12 °C | 28 °C | 22 °C | 12 °C | 28 °C | 28/22 °C | 28/12 °C | 28/22 °C | 28/12 °C | ||||
| Basal thermotolerance | AT1G74310 | Heat shock protein 101 (HSP101) | Bra003807 | 1457 | 1411 | 3457 | 1696 | 2238 | 3594 | 2.4 | 2.5 | 2.1 | 1.6 |
| Bra015922 | 2725 | 3674 | 6053 | 5409 | 5601 | 8821 | 2.2 | 1.6 | 1.6 | 1.6 | |||
| AT5G17020 | Exportin 1A (XPO1A) | Bra006382 | 9010 | 8711 | 9368 | 8345 | 9747 | 11,271 | 1.0 | 1.1 | 1.4 | 1.2 | |
| Bra008580 | 10,093 | 10,241 | 10,261 | 9834 | 8047 | 12,608 | 1.0 | 1.0 | 1.3 | 1.6 | |||
| Bra023593 | 6942 | 7056 | 7948 | 6939 | 6511 | 7561 | 1.1 | 1.1 | 1.1 | 1.2 | |||
| Acquired thermotolerance | AT3G25230 | Rotamase FKBP 1 (ROF1/FKBP62) | Bra013224 | 11 | 18 | 42 | 15 | 59 | 40 | 3.8 | 2.4 | 2.7 | 0.7 |
| AT5G48570 | Rotamase FKBP 2 (ROF2/FKBP65) | Bra037477 * | 309 | 283 | 4967 | 1632 | 352 | 6607 | 16.1 | 17.6 | 4.0 | 18.8 | |
| AT2G26150 | Heat shock transcription factor A2 (HSFA2) | Bra000557 * | 68 | 223 | 1256 | 256 | 542 | 1786 | 18.5 | 5.6 | 7.0 | 3.3 | |
| Warming | AT2G18790 | Phytochrome B (PHYB) | Bra001650 | 717 | 770 | 829 | 402 | 1083 | 527 | 1.2 | 1.1 | 1.3 | 0.5 |
| Bra022192 | 13,874 | 10,184 | 7989 | 16,575 | 11,087 | 13,685 | 0.6 | 0.8 | 0.8 | 1.2 | |||
| AT3G12580 | Heat shock protein 70 (HSP70) | Bra001457 | 1560 | 2512 | 8099 | 9758 | 6732 | 13,032 | 5.2 | 3.2 | 1.3 | 1.9 | |
| Bra038734 | 1049 | 1159 | 7871 | 6488 | 4093 | 7597 | 7.5 | 6.8 | 1.2 | 1.9 | |||
| AT2G43010 | Phytochrome interacting factor 4 (PIF4) | Bra000283 * | 19,533 | 12,261 | 15,413 | 13,610 | 9359 | 24,186 | 0.8 | 1.3 | 1.8 | 2.6 | |
| Bra037742 | 3718 | 4145 | 3350 | 1988 | 3218 | 4296 | 0.9 | 0.8 | 2.2 | 1.3 | |||
| Other HSPs | AT2G25140 | Casein lytic proteinase B4 (HSP98.7/CLPB4) | Bra007816 | 645 | 702 | 1353 | 1332 | 1269 | 2425 | 2.1 | 1.9 | 1.8 | 1.9 |
| AT4G16660 | HSP 70 family protein | Bra038496 | 1160 | 1868 | 2286 | 1503 | 2091 | 1505 | 2.0 | 1.2 | 1.0 | 0.7 | |
| AT4G25200 | Mitochondrion-localized small heat shock protein 23.6 (HSP23.6-MITO) | Bra013872 * | 333 | 323 | 1308 | 984 | 265 | 3551 | 3.9 | 4.0 | 3.6 | 13.4 | |
| AT4G10250 | HSP20-like chaperones superfamily protein (HSP22.0-L) | Bra000703 * | 1174 | 1055 | 1638 | 1352 | 2898 | 6069 | 1.4 | 1.6 | 4.5 | 2.1 | |
| Bra027999 * | 27 | 71 | 378 | 85 | 37 | 677 | 14.1 | 5.3 | 7.9 | 18.1 | |||
| AT4G27670 | Heat shock protein 21 (HSP21) | Bra026317 | 45 | 166 | 729 | 310 | 54 | 603 | 16.1 | 4.4 | 1.9 | 11.2 | |
| AT5G47590 | Heat shock protein HSP20/alpha crystallin family | Bra022051 | 1119 | 1560 | 1914 | 1045 | 3295 | 2588 | 1.7 | 1.2 | 2.5 | 0.8 | |
| Bra022079 | 4462 | 6564 | 7455 | 4309 | 9320 | 12,845 | 1.7 | 1.1 | 3.0 | 1.4 | |||
| Bra022083 | 1201 | 1103 | 1815 | 939 | 1902 | 1852 | 1.5 | 1.6 | 2.0 | 1.0 | |||
| Bra022084 | 1042 | 1120 | 1737 | 760 | 1635 | 1540 | 1.7 | 1.6 | 2.0 | 0.9 | |||
| AT1G53540 | HSP20-like chaperones superfamily protein | Bra018216 * | 10,773 | 3454 | 10,773 | 7716 | 3159 | 16,904 | 1.0 | 3.1 | 2.2 | 5.4 | |
| AT1G54050 | HSP20-like chaperones superfamily protein | Bra012949 | 5473 | 4945 | 10796 | 1568 | 1799 | 1843 | 2.0 | 2.2 | 1.2 | 1.0 | |
| Bra030910 * | 827 | 945 | 4041 | 2406 | 2649 | 9204 | 4.9 | 4.3 | 3.8 | 3.5 | |||
| AT2G29500 | HSP20-like chaperones superfamily protein | Bra018383 * | 1870 | 2774 | 11,166 | 5211 | 2428 | 19,026 | 6.0 | 4.0 | 3.7 | 7.8 | |
| Bra018384 | 2575 | 5095 | 6789 | 5906 | 5362 | 14,692 | 2.6 | 1.3 | 2.5 | 2.7 | |||
| Bra031725 | 1996 | 4409 | 5909 | 6292 | 5275 | 13,450 | 3.0 | 1.3 | 2.1 | 2.5 | |||
| AT4G27890 | HSP20-like chaperones superfamily protein | Bra040837 | 77 | 29 | 232 | 128 | 199 | 637 | 3.0 | 8.1 | 5.0 | 3.2 | |
| AT5G51440 | HSP20-like chaperones superfamily protein | Bra029174 | 406 | 376 | 5772 | 1657 | 292 | 7627 | 14.2 | 15.3 | 4.6 | 26.1 | |
| AT5G59720 | Heat shock protein 18.2 (HSP18.2) | Bra002539 * | 73 | 259 | 747 | 206 | 58 | 5114 | 10.2 | 2.9 | 24.8 | 88.5 | |
| Bra006697 * | 1129 | 779 | 2431 | 765 | 426 | 5739 | 2.2 | 3.1 | 7.5 | 13.5 | |||
| Bra020295 * | 1659 | 383 | 1167 | 2436 | 197 | 5383 | 0.7 | 3.0 | 2.2 | 27.3 | |||
| AT5G12020 | 17.6 kDa class II heat shock protein (HSP17.6II) | Bra006137 * | 52 | 271 | 2513 | 909 | 310 | 2036 | 48.6 | 9.3 | 2.2 | 6.6 | |
| Bra008920 | 3065 | 1599 | 5017 | 1267 | 1426 | 3693 | 1.6 | 3.1 | 2.9 | 2.6 | |||
| Gene | Line | Expression (Fold Change) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Heat Shock | Warming | |||||||||
| 22 °C | 27 °C | 32 °C | 37 °C | 42 °C | 22 °C | 12 °C | 28 °C | 45 °C | ||
| BrHSP18.2A | Chiifu | 1.0 | 2.2 | 10.3 | 3220.4 | 4132.9 | 1.0 | 0.5 | 8.4 | 7154.1 |
| Kenshin | 1.0 | 0.8 | 5.5 | 4921.5 | 49,617.7 | 1.0 | 0.5 | 106.7 | 34,142.0 | |
| BrHSP18.2B | Chiifu | 1.0 | 0.5 | 8.7 | 74.4 | 357.9 | 1.0 | 1.2 | 1.3 | 241.4 |
| Kenshin | 1.0 | 3.9 | 5.7 | 160.5 | 1738.6 | 1.0 | 1.8 | 42.3 | 31,296.8 | |
| BrHSP18.2C | Chiifu | 1.0 | 1.4 | 13.8 | 140.0 | 646.8 | 1.0 | 1.8 | 7.8 | 7540.8 |
| Kenshin | 1.0 | 2.3 | 4.0 | 84.1 | 2960.7 | 1.0 | 1.5 | 3.1 | 733.5 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, H.; Dong, X.; Yi, H.; Ahn, J.Y.; Yun, K.; Song, M.; Han, C.-T.; Hur, Y. Genome-Wide Identification and Characterization of Warming-Related Genes in Brassica rapa ssp. pekinensis. Int. J. Mol. Sci. 2018, 19, 1727. https://doi.org/10.3390/ijms19061727
Song H, Dong X, Yi H, Ahn JY, Yun K, Song M, Han C-T, Hur Y. Genome-Wide Identification and Characterization of Warming-Related Genes in Brassica rapa ssp. pekinensis. International Journal of Molecular Sciences. 2018; 19(6):1727. https://doi.org/10.3390/ijms19061727
Chicago/Turabian StyleSong, Hayoung, Xiangshu Dong, Hankuil Yi, Ju Young Ahn, Keunho Yun, Myungchul Song, Ching-Tack Han, and Yoonkang Hur. 2018. "Genome-Wide Identification and Characterization of Warming-Related Genes in Brassica rapa ssp. pekinensis" International Journal of Molecular Sciences 19, no. 6: 1727. https://doi.org/10.3390/ijms19061727
APA StyleSong, H., Dong, X., Yi, H., Ahn, J. Y., Yun, K., Song, M., Han, C. -T., & Hur, Y. (2018). Genome-Wide Identification and Characterization of Warming-Related Genes in Brassica rapa ssp. pekinensis. International Journal of Molecular Sciences, 19(6), 1727. https://doi.org/10.3390/ijms19061727