Genome-Wide Identification and Analysis of Polygalacturonase Genes in Solanum lycopersicum


 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Identification and Genomic Distribution of Tomato PG Genes
2.2. Sequence Analysis and Genomic Distribution of Tomato PG Genes
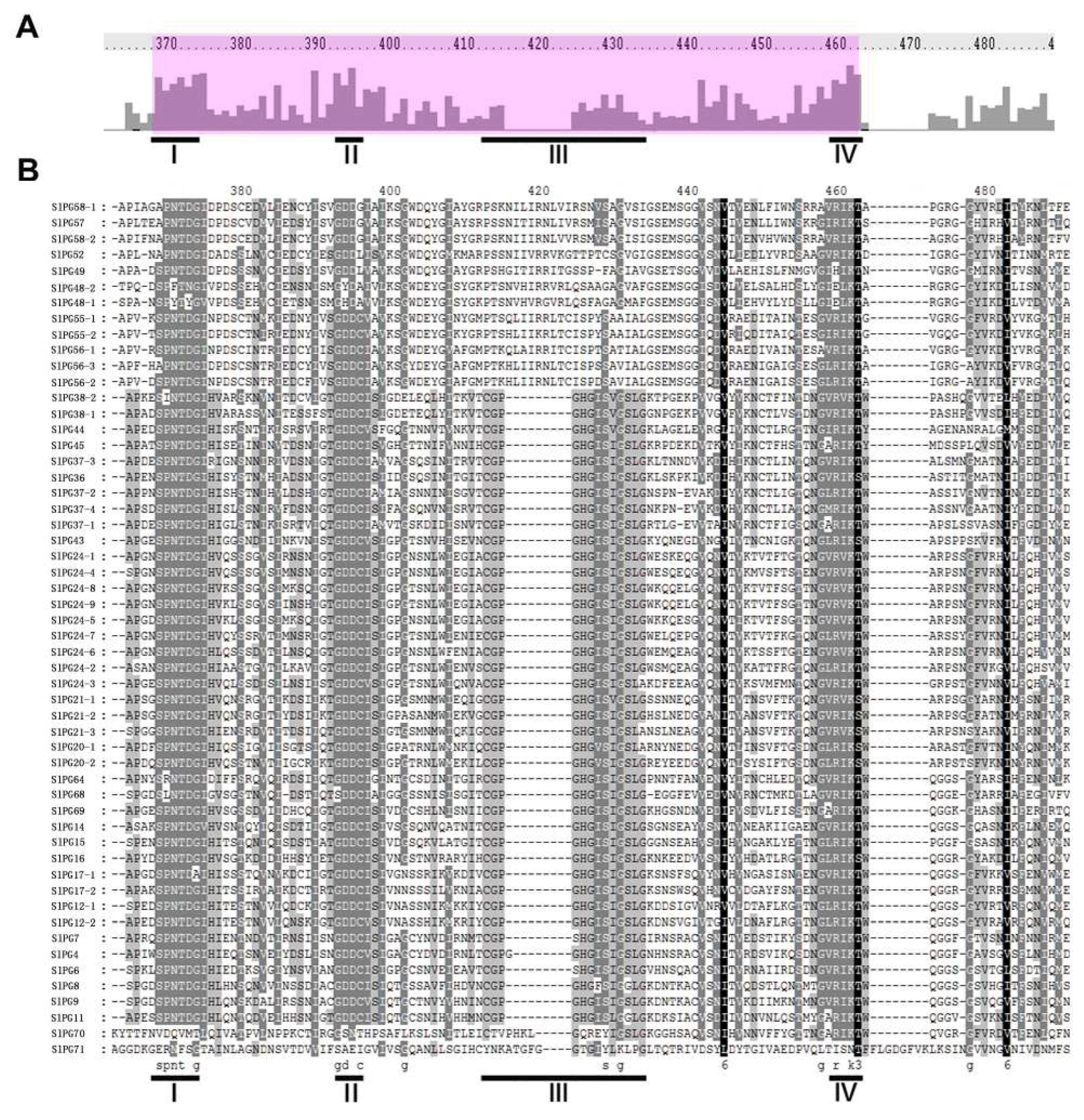
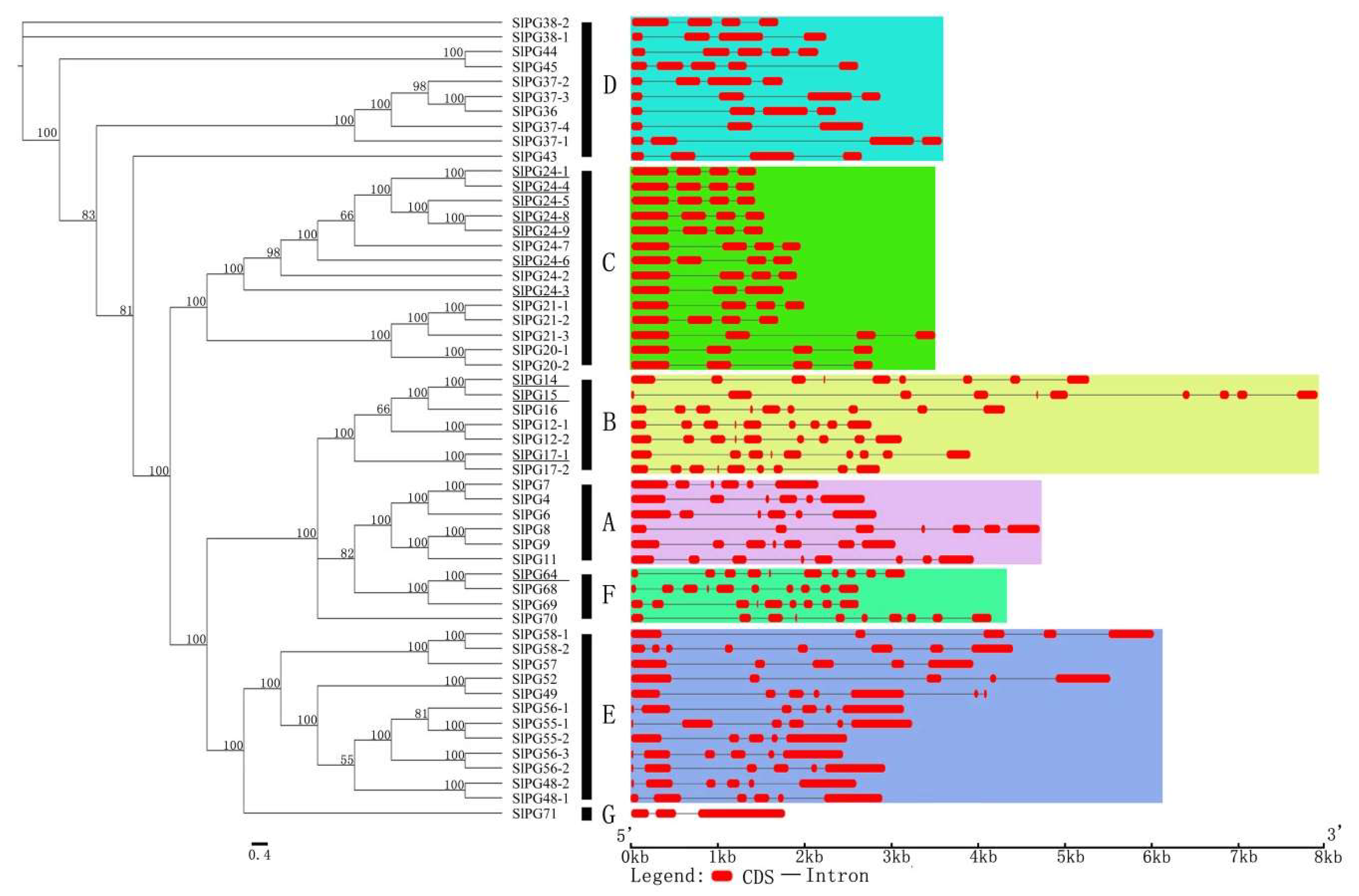
2.3. Phylogenetic and Gene Structure Analysis
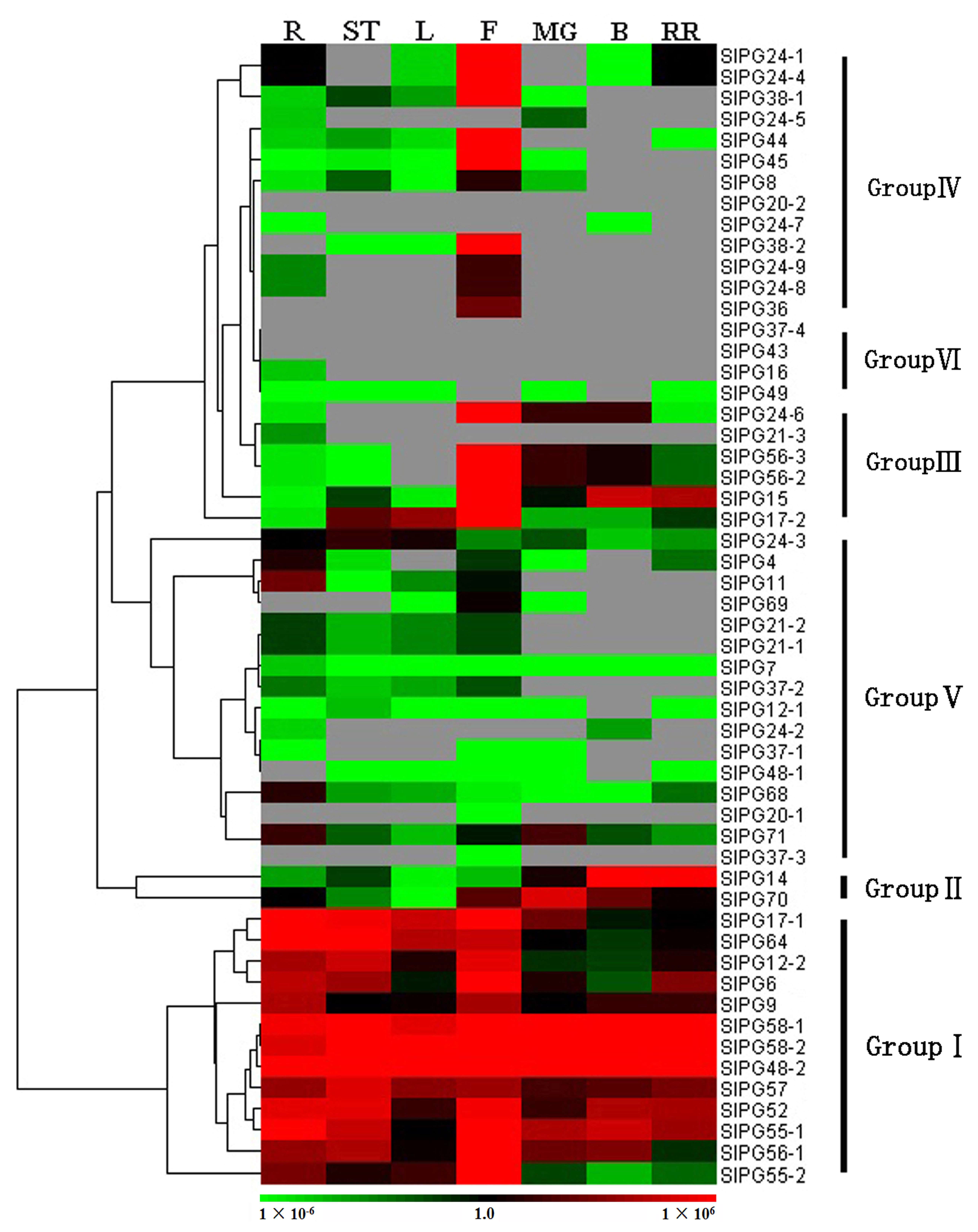
2.4. Expression Patterns of SlPG Genes
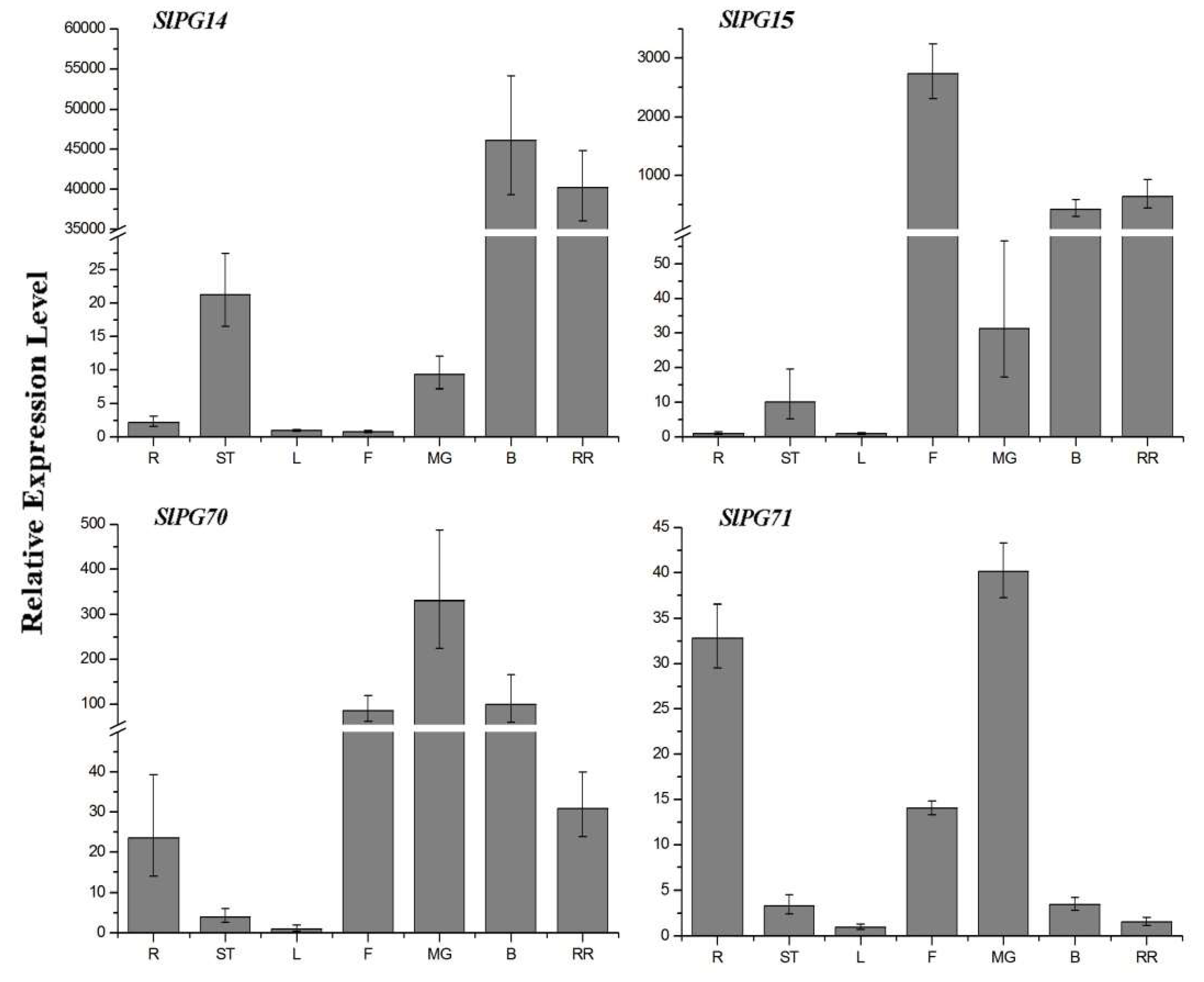
2.5. Specific Expression of PG Genes in Three Developmental Stages of Tomato Fruit
2.6. Promoter Analysis of the SlPGs
3. Discussion
4. Materials and Methods
4.1. Identification of Tomato PG Family Genes
4.2. Chromosome Localization and Sequence Characterization
4.3. Intron-Exon Structure and Phylogenetic Analyses
4.4. Plant Materials and Treatment
4.5. Expression Analysis of SlPG Genes by qRT-PCR
4.6. Cis-Regulatory Elements Analysis of SlPG Genes
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Caffall, K.H.; Mohnen, D. The structure, function, and biosynthesis of plant cell wall pectic polysaccharides. Carbohydr. Res. 2009, 344, 1879–1900. [Google Scholar] [CrossRef] [PubMed]
- Hadfield, K.A.; Bennett, A.B. Polygalacturonases: Many genes in search of a function. Plant Physiol. 1998, 117, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Shiu, S.H.; Thoma, S.; Li, W.H.; Patterson, S.E. Patterns of expansion and expression divergence in the plant polygalacturonase gene family. Genome Biol. 2006, 7, R87. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, M.; Kay, P.; Wilson, S.; Swain, S.M. Arabidopsis dehiscence zone polygalacturonase1 (ADPG1), ADPG2, and QUARTET2 are polygalacturonases required for cell separation during reproductive development in Arabidopsis. Plant Cell 2009, 21, 216–233. [Google Scholar] [CrossRef] [PubMed]
- Markovič, O.; Janeček, Š. Pectin degrading glycoside hydrolases of family 28: Sequence-structural features, specificities and evolution. Protein Eng. 2001, 14, 615–631. [Google Scholar] [CrossRef] [PubMed]
- Mohnen, D. Pectin structure and biosynthesis. Curr. Opin. Plant Biol. 2008, 11, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Yu, Y.; Cui, J.; Lyu, M.; Xu, L.; Cao, J. A comparative analysis of the evolution, expression, and cis-regulatory element of polygalacturonase genes in grasses and dicots. Funct. Integr. Genom. 2016, 16, 641–656. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Liang, Y.; Lyu, M.; Wu, J.; Lu, G.; Cao, J. Genome-wide identification and characterization of polygalacturonase genes in Cucumis sativus and Citrullus lanatus. Plant Physiol. Biochem. 2014, 74, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Torki, M.; Mandaron, P.; Mache, R.; Falconet, D. Characterization of a ubiquitous expressed gene family encoding polygalacturonase in Arabidopsis thaliana. Gene 2000, 242, 427–436. [Google Scholar] [CrossRef]
- Park, K.C.; Kwon, S.J.; Kim, N.S. Intron loss mediated structural dynamics and functional differentiation of the polygalacturonase gene family in land plants. Genes Genom. 2010, 32, 570–577. [Google Scholar] [CrossRef]
- Chen, H.; Shao, H.; Fan, S.; Ma, J.; Zhang, D.; Han, M. Identification and phylogenetic analysis of the POLYGALACTURONASE gene gamily in apple. Hortic. Plant J. 2016, 2, 241–252. [Google Scholar] [CrossRef]
- Liang, Y.; Yu, Y.; Shen, X.; Dong, H.; Lyu, M.; Xu, L.; Ma, Z.; Liu, T.; Cao, J. Dissecting the complex molecular evolution and expression of polygalacturonase gene family in Brassica rapa ssp. Chinensis. Plant Mol. Biol. 2015, 89, 629–646. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Somerville, C.; Anderson, C.T. Polygalacturonase involved in expansion1 functions in cell elongation and flower development in Arabidopsis. Plant Cell 2014, 26, 1018–1035. [Google Scholar] [CrossRef] [PubMed]
- Wakasa, Y.; Kudo, H.; Ishikawa, R.; Akada, S.; Senda, M.; Niizeki, M.; Harada, T. Low expression of an endopolygalacturonase gene in apple fruit with long-term storage potential. Postharvest Biol. Technol. 2006, 39, 193–198. [Google Scholar] [CrossRef]
- Asif, M.H.; Nath, P. Expression of multiple forms of polygalacturonase gene during ripening in banana fruit. Plant Physiol. Biochem. 2005, 43, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Sekine, D.; Munemura, I.; Gao, M.; Mitsuhashi, W.; Toyomasu, T.; Murayama, H. Cloning of cDNAs encoding cell-wall hydrolases from pear (Pyrus communis) fruit and their involvement in fruit softening and development of melting texture. Physiol. Plant. 2006, 126, 163–174. [Google Scholar] [CrossRef]
- Deytieux-Belleau, C.; Vallet, A.; Donèche, B.; Geny, L. Pectin methylesterase and polygalacturonase in the developing grape skin. Plant Physiol. Biochem. 2008, 46, 638–646. [Google Scholar] [CrossRef] [PubMed]
- Grierson, D. Control of ribonucleic acid and enzyme synthesis during fruit ripening. In Post-Harvest Physiology and Crop Preservation; Springer: New York, NY, USA, 1983; Volume 46, pp. 45–60. [Google Scholar]
- Hobson, G.E. The ripening of tomato fruit as affected by the injection of certain chemicals. J. Exp. Bot. 1965, 16, 411–422. [Google Scholar] [CrossRef]
- Tucker, G.A.; Robertson, N.G.; Grierson, D. Changes in polygalacturonase isoenzymes during the ‘ripening’ of normal and mutant tomato fruit. FEBS J. 1980, 112, 119–124. [Google Scholar] [CrossRef]
- Sheehy, R.E.; Kramer, M.; Hiatt, W.R. Reduction of polygalacturonase activity in tomato fruit by antisense RNA. Proc. Natl. Acad. Sci. USA 1988, 85, 8805–8809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giovannoni, J.J.; DellaPenna, D.; Bennett, A.B.; Fischer, R.L. Expression of a chimeric polygalacturonase gene in transgenic rin (ripening inhibitor) tomato fruit results in polyuronide degradation but not fruit softening. Plant Cell 1989, 1, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Langley, K.R.; Martin, A.; Stenning, R.; Murray, A.J.; Hobson, G.E.; Schuch, W.W.; Bird, C.R. Mechanical and optical assessment of the ripening of tomato fruit with reduced polygalacturonase activity. J. Sci. Food Agric. 1994, 66, 547–554. [Google Scholar] [CrossRef]
- Kramer, M.; Sanders, R.; Bolkan, H.; Waters, C.; Sheeny, R.E.; Hiatt, W.R. Postharvest evaluation of transgenic tomatoes with reduced levels of polygalacturonase: Processing, firmness and disease resistance. Postharvest Biol. Technol. 1992, 1, 241–255. [Google Scholar] [CrossRef]
- Fabi, J.P.; Broetto, S.G.; Da, S.S.; Zhong, S.; Lajolo, F.M.; do Nascimento, J.R. Analysis of papaya cell wall-related genes during fruit ripening indicates a central role of polygalacturonases during pulp softening. PLoS ONE 2013, 9, e105685. [Google Scholar] [CrossRef] [PubMed]
- Quesada, M.A.; Blanco-Portales, R.; Posé, S.; García-Gago, J.A.; Jiménez-Bermúdez, S.; Muñoz-Serrano, A.; Caballero, J.L.; Pliego-Alfaro, F.; Mercado, J.A.; Muñoz-Blanco, J. Antisense down-regulation of the FaPG1 gene reveals an unexpected central role for polygalacturonase in strawberry fruit softening. Plant Physiol. 2009, 150, 1022–1032. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, R.G.; Sutherland, P.W.; Johnston, S.L.; Kularajathevan, G.; Hallett, I.C.; Deepali, M.; Brummell, D.A.; Roswitha, S.; Johnston, J.W.; Schaffer, R.J. Down-regulation of POLYGALACTURONASE1 alters firmness, tensile strength and water loss in apple (Malus x domestica) fruit. BMC Plant Biol. 2012, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Uluisik, S.; Chapman, N.H.; Smith, R.; Poole, M.; Adams, G.; Gillis, R.B.; Besong, T.M.; Sheldon, J.; Stiegelmeyer, S.; Perez, L.; et al. Genetic improvement of tomato by targeted control of fruit softening. Nat. Biotechnol. 2016, 34, 950–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, K.C.; Kwon, S.J.; Kim, P.H.; Bureau, T.; Kim, N.S. Gene structure dynamics and divergence of the polygalacturonase gene family of plants and fungus. Genome 2008, 51, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.Y.; Osborne, E.; Poindexter, P.D.; Somerville, C.R. Microspore separation in the quartet 3 mutants of Arabidopsis is impaired by a defect in a developmentally regulated polygalacturonase required for pollen mother cell wall degradation. Plant Physiol. 2003, 133, 1170–1180. [Google Scholar] [CrossRef] [PubMed]
- Lyons, E.; Freeling, M. How to usefully compare homologous plant genes and chromosomes as DNA sequences. Plant J. 2008, 53, 661–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomato Genome Consortium. The tomato genome sequence provides insights into fleshy fruit evolution. Nature 2012, 485, 635–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinozaki, Y.; Nicolas, P.; Fernandez-Pozo, N.; Ma, Q.; Evanich, D.; Shi, Y.; Xu, Y.; Zheng, Y.; Snyder, S.I.; Martin, L.B.B.; et al. High-resolution spatiotemporal transcriptome mapping of tomato fruit development and ripening. Nat. Commun. 2018, 9, 364. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, S.; Zhang, J.; Sun, H.; Salse, J.; Lucas, W.J.; Zhang, H.; Zheng, Y.; Mao, L.; Ren, Y.; Wang, Z. The draft genome of watermelon (Citrullus lanatus) and resequencing of 20 diverse accessions. Nat. Genet. 2013, 45, 51–82. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, H.; Wang, J.; Sun, R.; Wu, J.; Liu, S.; Bai, Y.; Mun, J.H.; Bancroft, I.; Cheng, F. The genome of the mesopolyploid crop species Brassica rapa. Nat. Genet. 2011, 43, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
- Grierson, D.; Tucker, G.A.; Keen, J.; Ray, J.; Bird, C.R.; Schuch, W. Sequencing and identification of a cDNA clone for tomato polygalacturonase. Nucleic Acids Res. 1986, 14, 8595–8603. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.J.; Watson, C.F.; Morris, P.C.; Bird, C.R.; Seymour, G.B.; Gray, J.E.; Arnold, C.; Tucker, G.A.; Schuch, W.; Harding, S. Inheritance and effect on ripening of antisense polygalacturonase genes in transgenic tomatoes. Plant Mol. Biol. 1990, 14, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Brummell, D.A.; Labavitch, J.M. Effect of antisense suppression of endopolygalacturonase activity on polyuronide molecular weight in ripening tomato fruit and in fruit homogenates. Plant Physiol. 1997, 115, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Giovannoni, J. Molecular biology of fruit maturation and ripening. Annu. Rev. Plant. Physiol. Plant Mol. Biol. 2001, 52, 725–749. [Google Scholar] [CrossRef] [PubMed]
- Gorguet, B.; Schipper, D.; van Lammeren, A.; Visser, R.G.; van Heusden, A.W. ps-2, the gene responsible for functional sterility in tomato, due to non-dehiscent anthers, is the result of a mutation in a novel polygalacturonase gene. Theor. Appl. Genet. 2009, 118, 1199–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlot, A.C.; Dempsey, D.A.; Klessig, D.F. Salicylic Acid, a multifaceted hormone to combat disease. Annu. Rev. Phytopathol. 2009, 47, 177–206. [Google Scholar] [CrossRef] [PubMed]
- Cara, B.; Giovannoni, J.J. Molecular biology of ethylene during tomato fruit development and maturation. Plant Sci. 2008, 175, 106–113. [Google Scholar] [CrossRef]
- Petersen, T.N.; Brunak, S.; Von, H.G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Emanuelsson, O.; Nielsen, H.; Brunak, S.; von Heijne, G. Predicting subcellular localization of proteins based on their N-terminal amino acid sequence. J. Mol. Biol. 2000, 300, 1005–1016. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Liu, S. Genome-wide analysis of the MADS-box gene family in cucumber. Genome 2012, 55, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Ronquist, F. Bayesian analysis of molecular evolution using MrBayes. In Statistical Methods in Molecular Evolution; Springer: New York, NY, USA, 2005; Volume 113, pp. 183–226. [Google Scholar]
- Lanave, C.; Preparata, G.; Saccone, C.; Serio, G. A new method for calculating evolutionary substitution rates. J. Mol. Evol. 1984, 20, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Tavarè, S. Some Probabilistic and Statistical Problems on the Analysis of DNA Sequences. In American Mathematical Society (Ed.), Lectures on Mathematics in the Life Sciences; American Mathematical Society: Providence, RI, USA, 1986; pp. 57–86. [Google Scholar]
- Yang, Z.; Goldman, N.; Friday, A. Comparison of models for nucleotide substitution used in maximum-likelihood phylogenetic estimation. Mol. Biol. Evol. 1994, 11, 316–324. [Google Scholar] [PubMed]
- Meli, V.S.; Ghosh, S.; Prabha, T.N.; Chakraborty, N.; Chakraborty, S.; Datta, A. Enhancement of fruit shelf life by suppressing N-glycan processing enzymes. Proc. Natl. Acad. Sci. USA 2010, 107, 2413–2418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, F.; Wang, J.; Cheng, L.; Liu, S.; Wu, J.; Peng, Z.; Lu, G. Genome-wide analysis of the mitogen-activated protein kinase gene family in Solanum lycopersicum. Gene 2012, 499, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Eisen, M.B.; Spellman, P.T.; Brown, P.O.; Botstein, D. Cluster analysis and display of genome-wide expression patterns. Proc. Natl. Acad. Sci. USA 1998, 95, 14863–14868. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene * | Solanum lycopersicum Locus | Chr | Location (5′–3′) | Predicted Protein (aa) | Molecular Weight (kDa) | Isoelectric Points (pI) | Signal Peptides (aa) | Sub ** Cellular Locationaa | Domains |
|---|---|---|---|---|---|---|---|---|---|
| SlPG38-1 | Solyc00g030510.2.1 | 0 | 12605208–12607452 | 403 | 42.47 | 6.96 | 1–23 | S | I II III IV |
| SlPG37-4 | Solyc01g009640.1.1 | 1 | 3883446–3879488 | 391 | 42 | 6.81 | 1–22 | S | I II III IV |
| SlPG44 | Solyc01g066070.2.1 | 1 | 65439007–65436858 | 405 | 43.51 | 7.92 | 1–25 | S | I II III IV |
| SlPG4 | Solyc01g079130.1.1 | 1 | 70719258–70721952 | 472 | 51.51 | 5.96 | 1–30 | S | I II III IV |
| SlPG16 | Solyc01g087280.1.1 | 1 | 73976402–73980711 | 425 | 47.65 | 9.46 | 1–22 | S | I II III IV |
| SlPG55-2 | Solyc01g094970.2.1 | 1 | 78127060–78124570 | 475 | 51.83 | 6.63 | 1–24 | S | I II IV |
| SlPG7 | Solyc01g100980.2.1 | 1 | 82616598–82614439 | 476 | 52.16 | 7.44 | S | I II III IV | |
| SlPG24-8 (TAPG1) | Solyc02g067630.2.1 | 2 | 32341911–32343450 | 392 | 41.85 | 8.87 | 1–17 | S | I II III IV |
| SlPG24-9 (TAPG2) | Solyc02g067640.2.1 | 2 | 32347519–32349041 | 392 | 41.87 | 8.49 | 1–17 | S | I II III IV |
| SlPG24-5 (TAPG3) | Solyc02g067650.1.1 | 2 | 32355651–32357075 | 389 | 41.75 | 8.34 | 1–17 | S | I II III IV |
| SlPG71 | Solyc02g068400.2.1 | 2 | 32931014–32932787 | 481 | 51.66 | 5.92 | 1–22 | S | |
| SlPG43 | Solyc02g069480.1.1 | 2 | 33931720–33929059 | 393 | 42.08 | 6.4 | 1–28 | S | I II III IV |
| SlPG56-2 | Solyc03g007940.2.1 | 3 | 2435889–2432960 | 467 | 50.81 | 5.67 | 1–31 | S | I II IV |
| SlPG56-3 | Solyc03g007950.2.1 | 3 | 2441660–2439216 | 467 | 50.95 | 6 | 1–31 | S | I II IV |
| SlPG69 | Solyc03g113230.1.1 | 3 | 57495929–57493305 | 394 | 43.12 | 5.72 | 1–22 | S | I II III IV |
| SlPG17-1 (XPG1) | Solyc03g116500.2.1 | 3 | 59953586–59957497 | 452 | 49.44 | 8.83 | S | I II III IV | |
| SlPG68 | Solyc03g116580.2.1 | 3 | 59990295–59986839 | 437 | 47.05 | 8.11 | 1–24 | S | I II III IV |
| SlPG57 | Solyc03g117750.2.1 | 3 | 60847079–60843135 | 487 | 54.84 | 9.26 | M | I II IV | |
| SlPG6 | Solyc04g008230.2.1 | 4 | 1904669–1901842 | 490 | 52.87 | 4.88 | 1–24 | C | I II III IV |
| SlPG15 (PS-2) | Solyc04g015530.1.1 | 4 | 5755310–5764191 | 463 | 50.99 | 7.49 | I II III IV | ||
| SlPG12-2 | Solyc04g025440.2.1 | 4 | 20488981–20492100 | 460 | 49.82 | 6.33 | 1–18 | S | I II III IV |
| SlPG11 | Solyc05g005040.2.1 | 5 | 62026–58076 | 471 | 52.07 | 9.04 | 1–28 | S | I II III IV |
| SlPG9 | Solyc05g005170.2.1 | 5 | 159216–162264 | 508 | 55.67 | 8.7 | 1–30 | S | I II III IV |
| SlPG55-1 | Solyc05g049980.2.1 | 5 | 58973161–58969921 | 486 | 53.4 | 5.57 | S | I II IV | |
| SlPG45 | Solyc06g009200.2.1 | 6 | 3145482–3148100 | 410 | 43.42 | 6.78 | 1–31 | S | I II III IV |
| SlPG48-1 | Solyc06g009790.2.1 | 6 | 3813956–3809878 | 457 | 49.35 | 5.09 | 1–22 | S | I II IV |
| SlPG52 | Solyc06g060170.2.1 | 6 | 34525051–34519529 | 488 | 53.94 | 8.63 | I II IV | ||
| SlPG17-2 | Solyc06g068040.2.1 | 6 | 38555860–38558728 | 437 | 48.58 | 9.19 | S | I II III IV | |
| SlPG8 | Solyc07g015870.2.1 | 7 | 5634411–5629702 | 449 | 49.21 | 5.9 | 1–26 | S | I II III IV |
| SlPG37-2 | Solyc07g041650.1.1 | 7 | 51162465–51164213 | 389 | 41.82 | 7.53 | 1–22 | S | I II III IV |
| SlPG58-2 | Solyc07g042160.2.1 | 7 | 52482303–52486747 | 493 | 55.05 | 9.3 | I II IV | ||
| SlPG38-2 | Solyc07g044870.2.1 | 7 | 55248946–55251510 | 405 | 43.3 | 7 | 1–23 | S | I II III IV |
| SlPG37-1 | Solyc07g056290.1.1 | 7 | 61470406–61473987 | 401 | 43.96 | 8.54 | S | I II III IV | |
| SlPG20-1 | Solyc08g014540.1.1 | 8 | 4615191–4612405 | 395 | 42.92 | 9.2 | 1–25 | S | I II III IV |
| SlPG21-3 | Solyc08g014560.1.1 | 8 | 4650562–4654908 | 396 | 43.39 | 9.47 | S | I II III IV | |
| SlPG64 (PGcat) | Solyc08g060970.2.1 | 8 | 43357711–43354556 | 423 | 46.31 | 8.2 | S | I II III IV | |
| SlPG56-1 | Solyc08g081480.2.1 | 8 | 61693642–61696788 | 483 | 52.9 | 5.18 | S | I II IV | |
| SlPG49 | Solyc08g082170.2.1 | 8 | 62200995–62196897 | 467 | 50.97 | 8.35 | 1–25 | S | I II IV |
| SlPG48-2 | Solyc09g075460.2.1 | 9 | 62665686–62668281 | 446 | 48.62 | 5.3 | 1–28 | S | I II IV |
| SlPG12-1 | Solyc09g098270.2.1 | 9 | 67421613–67424507 | 433 | 47.3 | 4.95 | I II III IV | ||
| SlPG36 | Solyc10g047570.1.1 | 10 | 36889700–36892063 | 392 | 41.9 | 6.29 | 1–22 | S | I II III IV |
| SlPG37-3 | Solyc10g047590.1.1 | 10 | 36929234–36932108 | 387 | 42.31 | 9.01 | 1–22 | S | I II III IV |
| SlPG14 (pTOM6) | Solyc10g080210.1.1 | 10 | 60890335–60883700 | 457 | 50.05 | 6.4 | 1–24 | S | I II III IV |
| SlPG58-1 | Solyc12g009210.1.1 | 12 | 2490735–2496842 | 495 | 55.53 | 8.98 | M | I II IV | |
| SlPG70 | Solyc12g009420.1.1 | 12 | 2702056–2697739 | 426 | 46.34 | 7.47 | S | IV | |
| SlPG20-2 | Solyc12g019120.1.1 | 12 | 9156572–9152746 | 395 | 43.08 | 8.9 | 1–27 | S | I II III IV |
| SlPG21-2 | Solyc12g019130.1.1 | 12 | 9245985–9244294 | 390 | 42.5 | 9.66 | 1–24 | S | I II III IV |
| SlPG21-1 | Solyc12g019140.1.1 | 12 | 9293132–9295123 | 389 | 42.4 | 9.77 | 1–23 | S | I II III IV |
| SlPG24-6 (TPG7) | Solyc12g019180.1.1 | 12 | 9461467–9459617 | 397 | 42.15 | 6.05 | 1–25 | S | I II III IV |
| SlPG24-2 | Solyc12g019220.1.1 | 12 | 9700427–9702341 | 395 | 42.09 | 9.08 | 1–25 | S | I II III IV |
| SlPG24-7 | Solyc12g019230.1.1 | 12 | 9708477–9710424 | 391 | 42.59 | 8.7 | 1–24 | S | I II III IV |
| SlPG24-3 (TAPG6) | Solyc12g096730.1.1 | 12 | 63866593–63868349 | 395 | 42.42 | 8.34 | S | I II III IV | |
| SlPG24-1 (TAPG5) | Solyc12g096740.1.1 | 12 | 63870294–63868858 | 387 | 41.31 | 8.5 | 1–17 | S | I II III IV |
| SlPG24-4 (TAPG4) | Solyc12g096750.1.1 | 12 | 63877420–63878834 | 387 | 41.37 | 6.44 | 1–17 | S | I II III IV |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ke, X.; Wang, H.; Li, Y.; Zhu, B.; Zang, Y.; He, Y.; Cao, J.; Zhu, Z.; Yu, Y. Genome-Wide Identification and Analysis of Polygalacturonase Genes in Solanum lycopersicum. Int. J. Mol. Sci. 2018, 19, 2290. https://doi.org/10.3390/ijms19082290
Ke X, Wang H, Li Y, Zhu B, Zang Y, He Y, Cao J, Zhu Z, Yu Y. Genome-Wide Identification and Analysis of Polygalacturonase Genes in Solanum lycopersicum. International Journal of Molecular Sciences. 2018; 19(8):2290. https://doi.org/10.3390/ijms19082290
Chicago/Turabian StyleKe, Xubo, Huasen Wang, Yang Li, Biao Zhu, Yunxiang Zang, Yong He, Jiashu Cao, Zhujun Zhu, and Youjian Yu. 2018. "Genome-Wide Identification and Analysis of Polygalacturonase Genes in Solanum lycopersicum" International Journal of Molecular Sciences 19, no. 8: 2290. https://doi.org/10.3390/ijms19082290
APA StyleKe, X., Wang, H., Li, Y., Zhu, B., Zang, Y., He, Y., Cao, J., Zhu, Z., & Yu, Y. (2018). Genome-Wide Identification and Analysis of Polygalacturonase Genes in Solanum lycopersicum. International Journal of Molecular Sciences, 19(8), 2290. https://doi.org/10.3390/ijms19082290