Kinesin-2 Controls the Motility of RAB5 Endosomes and Their Association with the Spindle in Mitosis

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Kinesin-2 Silencing Affects Long-Range Movement of Early Endosomes in Interphase Cells
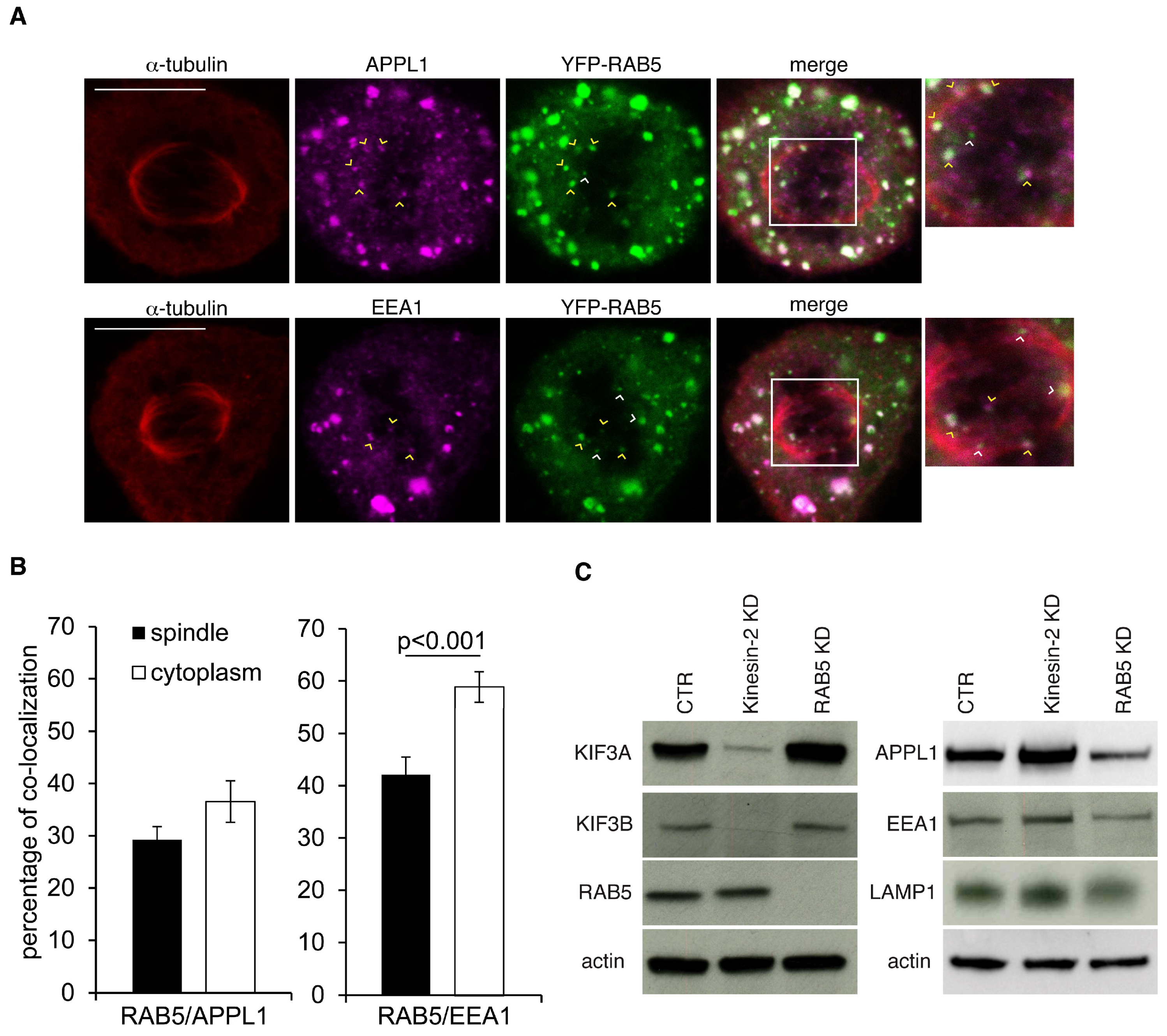
2.2. Depletion of Kinesin-2 Impairs Localization of RAB5 Endosomes to the Mitotic Spindle
2.3. The Mitotic Spindle Is Populated by RAB5 Early Endosomes That Carry the APPL1 Marker
2.4. Kinesin-2 Silencing Delays Nuclear Envelope Breakdown and Entry into Mitosis
3. Discussion
4. Material and Methods
4.1. Cells Culture and Expression Vectors
4.2. Antibodies
4.3. Immunofluorescence
4.4. Live Imaging and Analysis
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Zeigerer, A.; Gilleron, J.; Bogorad, R.L.; Marsico, G.; Nonaka, H.; Seifert, S.; Epstein-Barash, H.; Kuchimanchi, S.; Peng, C.G.; Ruda, V.M.; et al. Rab5 is necessary for the biogenesis of the endolysosomal system in vivo. Nature 2012, 485, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Gruenberg, J. The endocytic pathway: A mosaic of domains. Nat. Rev. Mol. Cell Biol. 2001, 2, 721–730. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Hong, W. From endosomes to the trans-golgi network. Semin. Cell Dev. Biol. 2014, 31, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Kalaidzidis, I.; Miaczynska, M.; Brewińska-Olchowik, M.; Hupalowska, A.; Ferguson, C.; Parton, R.G.; Kalaidzidis, Y.; Zerial, M. Appl endosomes are not obligatory endocytic intermediates but act as stable cargo-sorting compartments. J. Cell Biol. 2015, 211, 123–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonifacino, J.S.; Neefjes, J. Moving and positioning the endolysosomal system. Curr. Opin. Cell Biol. 2017, 47, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.R.; Maritzen, T.; Kukhtina, V.; Higman, V.A.; Doglio, L.; Barak, N.N.; Strauss, H.; Oschkinat, H.; Dotti, C.G.; Haucke, V. Regulation of endosomal membrane traffic by a gadkin/ap-1/kinesin kif5 complex. Proc. Natl. Acad. Sci. USA 2009, 106, 15344–15349. [Google Scholar] [CrossRef] [PubMed]
- Farkhondeh, A.; Niwa, S.; Takei, Y.; Hirokawa, N. Characterizing kif16b in neurons reveals a novel intramolecular “Stalk inhibition” Mechanism that regulates its capacity to potentiate the selective somatodendritic localization of early endosomes. J. Neurosci. 2015, 35, 5067–5086. [Google Scholar] [CrossRef] [PubMed]
- Hoepfner, S.; Severin, F.; Cabezas, A.; Habermann, B.; Runge, A.; Gillooly, D.; Stenmark, H.; Zerial, M. Modulation of receptor recycling and degradation by the endosomal kinesin kif16b. Cell 2005, 121, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, H.; Nakata, T.; Okada, Y.; Hirokawa, N. Kif3a/b: A heterodimeric kinesin superfamily protein that works as a microtubule plus end-directed motor for membrane organelle transport. J. Cell Biol. 1995, 130, 1387–1399. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, H.; Nakata, T.; Okada, Y.; Hirokawa, N. Cloning and characterization of kap3: A novel kinesin superfamily-associated protein of kif3a/3b. Proc. Natl. Acad. Sci. USA 1996, 93, 8443–8448. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, S.P.; Guzik-Lendrum, S.; Rayment, I. Kinesin-2 motors: Kinetics and biophysics. J. Biol. Chem. 2018, 293, 4510–4518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, S.; Yonekawa, Y.; Tanaka, Y.; Okada, Y.; Nonaka, S.; Hirokawa, N. Left-right asymmetry and kinesin superfamily protein kif3a: New insights in determination of laterality and mesoderm induction by kif3a-/-mice analysis. J. Cell Biol. 1999, 145, 825–836. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, S.; Tanaka, Y.; Okada, Y.; Takeda, S.; Harada, A.; Kanai, Y.; Kido, M.; Hirokawa, N. Randomization of left-right asymmetry due to loss of nodal cilia generating leftward flow of extraembryonic fluid in mice lacking kif3b motor protein. Cell 1998, 95, 829–837. [Google Scholar] [CrossRef]
- Huangfu, D.; Liu, A.; Rakeman, A.S.; Murcia, N.S.; Niswander, L.; Anderson, K.V. Hedgehog signalling in the mouse requires intraflagellar transport proteins. Nature 2003, 426, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.L.; Maier, K.C.; Stauber, T.; Ginkel, L.M.; Wordeman, L.; Vernos, I.; Schroer, T.A. Kinesin-2 is a motor for late endosomes and lysosomes. Traffic 2005, 6, 1114–1124. [Google Scholar] [CrossRef] [PubMed]
- Loubéry, S.; Wilhelm, C.; Hurbain, I.; Neveu, S.; Louvard, D.; Coudrier, E. Different microtubule motors move early and late endocytic compartments. Traffic 2008, 9, 492–509. [Google Scholar] [CrossRef] [PubMed]
- Stenmark, H. Rab gtpases as coordinators of vesicle traffic. Nat. Rev. Mol. Cell Biol. 2009, 10, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Kerr, M.; Teasdale, R.D. Live imaging of endosome dynamics. Semin. Cell Dev. Biol. 2014, 31, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Hirokawa, N.; Noda, Y.; Tanaka, Y.; Niwa, S. Kinesin superfamily motor proteins and intracellular transport. Nat. Rev. Mol. Cell Biol. 2009, 10, 682–696. [Google Scholar] [CrossRef] [PubMed]
- Gasman, S.; Kalaidzidis, Y.; Zerial, M. Rhod regulates endosome dynamics through diaphanous-related formin and src tyrosine kinase. Nat. Cell Biol. 2003, 5, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Serio, G.; Margaria, V.; Jensen, S.; Oldani, A.; Bartek, J.; Bussolino, F.; Lanzetti, L. Small gtpase rab5 participates in chromosome congression and regulates localization of the centromere-associated protein cenp-f to kinetochores. Proc. Natl. Acad. Sci. USA 2011, 108, 17337–17342. [Google Scholar] [CrossRef] [PubMed]
- Haraguchi, K.; Hayashi, T.; Jimbo, T.; Yamamoto, T.; Akiyama, T. Role of the kinesin-2 family protein, kif3, during mitosis. J. Biol. Chem. 2006, 281, 4094–4099. [Google Scholar] [CrossRef] [PubMed]
- Hehnly, H.; Doxsey, S. Rab11 endosomes contribute to mitotic spindle organization and orientation. Dev. Cell 2014, 28, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Capalbo, L.; D’Avino, P.P.; Archambault, V.; Glover, D.M. Rab5 gtpase controls chromosome alignment through lamin disassembly and relocation of the numa-like protein mud to the poles during mitosis. Proc. Natl. Acad. Sci. USA 2011, 108, 17343–17348. [Google Scholar] [CrossRef] [PubMed]
- Audhya, A.; Desai, A.; Oegema, K. A role for rab5 in structuring the endoplasmic reticulum. J. Cell Biol. 2007, 178, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Disanza, A.; Frittoli, E.; Palamidessi, A.; Scita, G. Endocytosis and spatial restriction of cell signaling. Mol. Oncol. 2009, 3, 280–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onnis, A.; Finetti, F.; Patrussi, L.; Gottardo, M.; Cassioli, C.; Spanò, S.; Baldari, C.T. The small gtpase rab29 is a common regulator of immune synapse assembly and ciliogenesis. Cell Death Differ. 2015, 22, 1687–1699. [Google Scholar] [CrossRef] [PubMed]
- Imamura, T.; Huang, J.; Usui, I.; Satoh, H.; Bever, J.; Olefsky, J.M. Insulin-induced glut4 translocation involves protein kinase c-lambda-mediated functional coupling between rab4 and the motor protein kinesin. Mol. Cell Biol. 2003, 23, 4892–4900. [Google Scholar] [CrossRef] [PubMed]
- De Renzis, S.; Sönnichsen, B.; Zerial, M. Divalent rab effectors regulate the sub-compartmental organization and sorting of early endosomes. Nat. Cell Biol. 2002, 4, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Sonnichsen, B.; De Renzis, S.; Nielsen, E.; Rietdorf, J.; Zerial, M. Distinct membrane domains on endosomes in the recycling pathway visualized by multicolor imaging of rab4, rab5, and rab11. J. Cell Biol. 2000, 149, 901–914. [Google Scholar] [CrossRef] [PubMed]
- Kälin, S.; Buser, D.P.; Spiess, M. A fresh look at the function of rabaptin5 on endosomes. Small GTPases 2016, 7, 34–37. [Google Scholar] [CrossRef] [PubMed]
- Kutay, U.; Hetzer, M.W. Reorganization of the nuclear envelope during open mitosis. Curr. Opin. Cell Biol. 2008, 20, 669–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miaczynska, M.; Christoforidis, S.; Giner, A.; Shevchenko, A.; Uttenweiler-Joseph, S.; Habermann, B.; Wilm, M.; Parton, R.G.; Zerial, M. Appl proteins link rab5 to nuclear signal transduction via an endosomal compartment. Cell 2004, 116, 445–456. [Google Scholar] [CrossRef]
- Pupo, E.; Ducano, N.; Lupo, B.; Vigna, E.; Avanzato, D.; Perera, T.; Trusolino, L.; Lanzetti, L.; Comoglio, P.M. Rebound effects caused by withdrawal of met kinase inhibitor are quenched by a met therapeutic antibody. Cancer Res. 2016, 76, 5019–5029. [Google Scholar] [CrossRef] [PubMed]
- DiMilla, P.A.; Stone, J.A.; Quinn, J.A.; Albelda, S.M.; Lauffenburger, D.A. Maximal migration of human smooth muscle cells on fibronectin and type iv collagen occurs at an intermediate attachment strength. J. Cell Biol. 1993, 122, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Zaman, M.H.; Matsudaira, P.; Lauffenburger, D.A. Understanding effects of matrix protease and matrix organization on directional persistence and translational speed in three-dimensional cell migration. Ann. Biomed. Eng. 2007, 35, 91–100. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pupo, E.; Avanzato, D.; Scianna, M.; Oldani, A.; Serini, G.; Lanzetti, L. Kinesin-2 Controls the Motility of RAB5 Endosomes and Their Association with the Spindle in Mitosis. Int. J. Mol. Sci. 2018, 19, 2575. https://doi.org/10.3390/ijms19092575
Pupo E, Avanzato D, Scianna M, Oldani A, Serini G, Lanzetti L. Kinesin-2 Controls the Motility of RAB5 Endosomes and Their Association with the Spindle in Mitosis. International Journal of Molecular Sciences. 2018; 19(9):2575. https://doi.org/10.3390/ijms19092575
Chicago/Turabian StylePupo, Emanuela, Daniele Avanzato, Marco Scianna, Amanda Oldani, Guido Serini, and Letizia Lanzetti. 2018. "Kinesin-2 Controls the Motility of RAB5 Endosomes and Their Association with the Spindle in Mitosis" International Journal of Molecular Sciences 19, no. 9: 2575. https://doi.org/10.3390/ijms19092575
APA StylePupo, E., Avanzato, D., Scianna, M., Oldani, A., Serini, G., & Lanzetti, L. (2018). Kinesin-2 Controls the Motility of RAB5 Endosomes and Their Association with the Spindle in Mitosis. International Journal of Molecular Sciences, 19(9), 2575. https://doi.org/10.3390/ijms19092575