Myelination in Multiple Sclerosis Lesions Is Associated with Regulation of Bone Morphogenetic Protein 4 and Its Antagonist Noggin

Abstract
:1. Introduction
2. Results
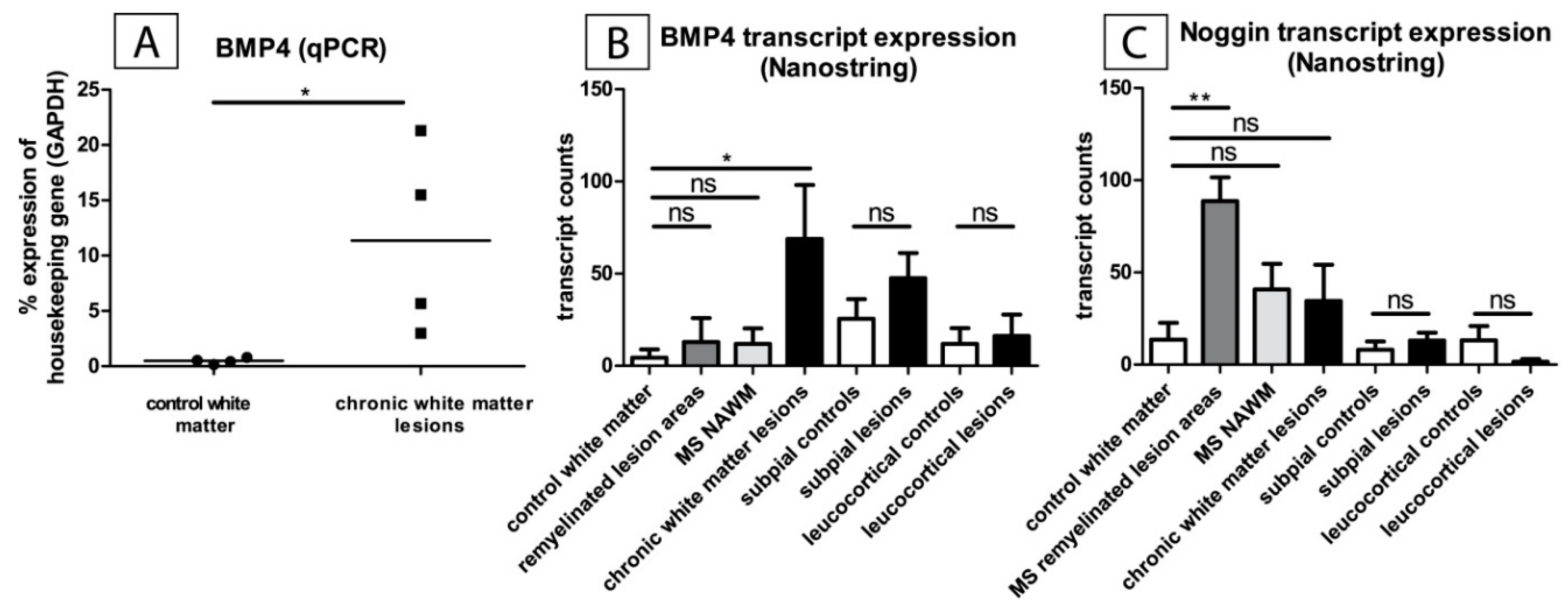
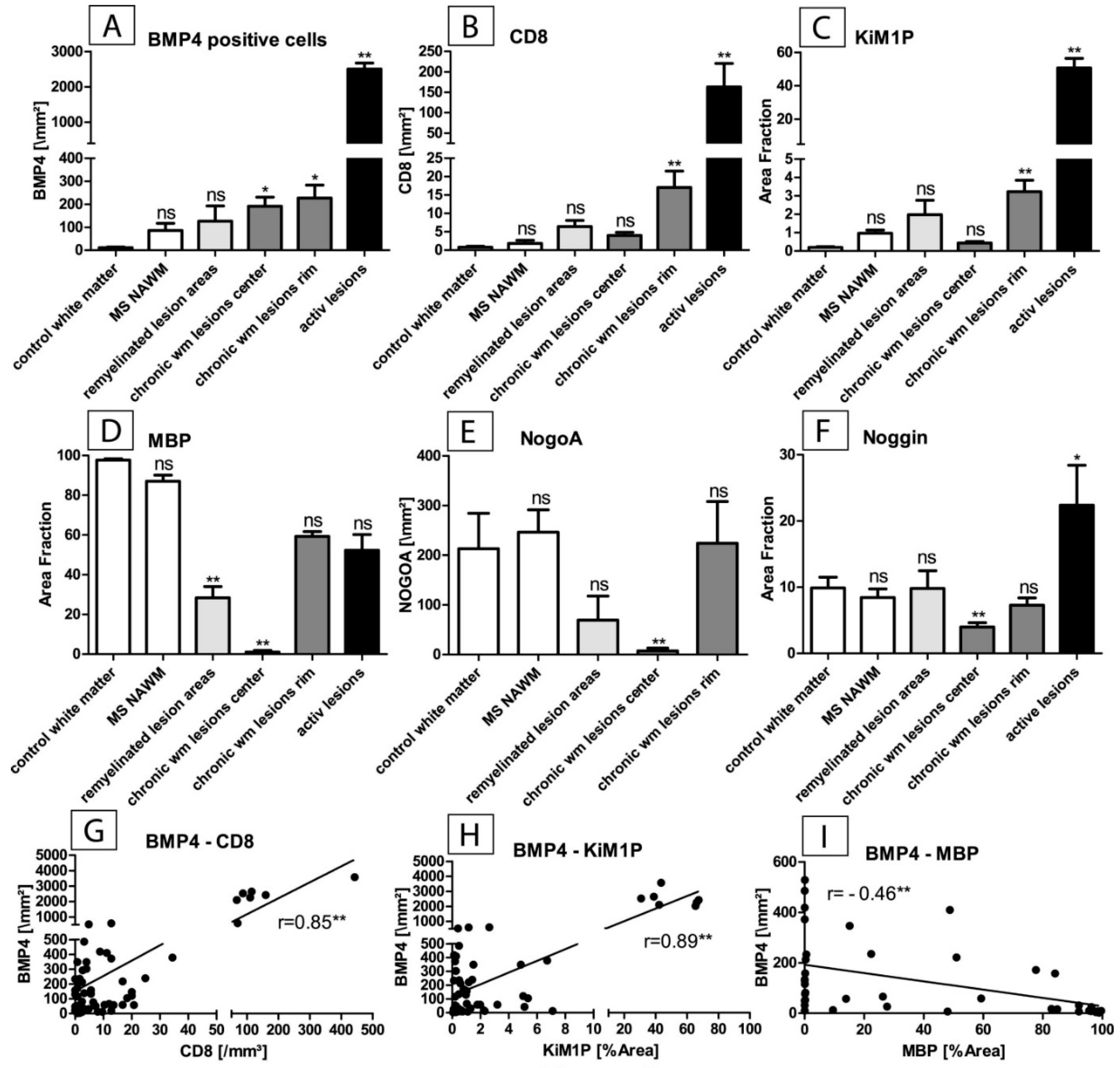
2.1. BMP4 Is Upregulated in MS-Lesions
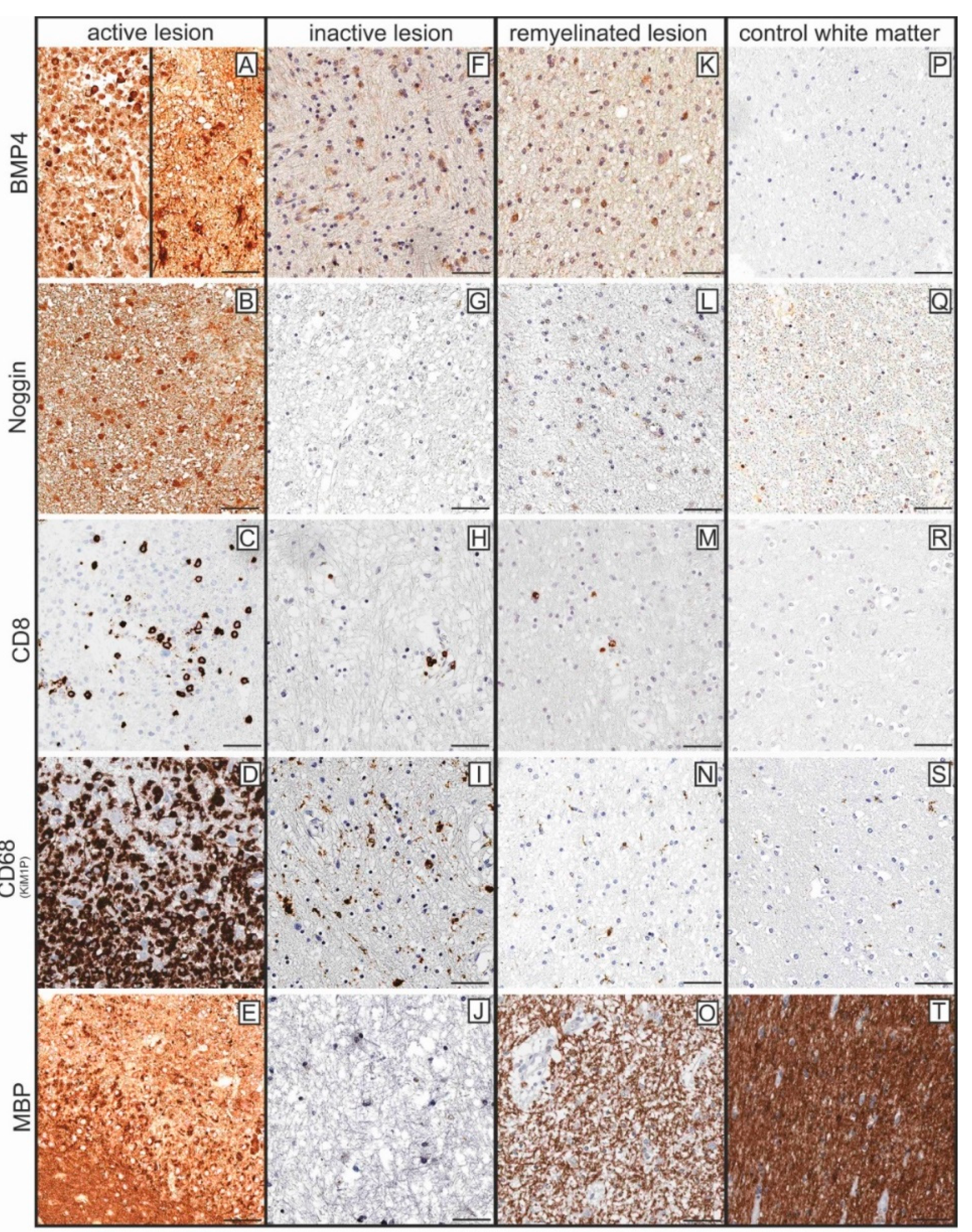
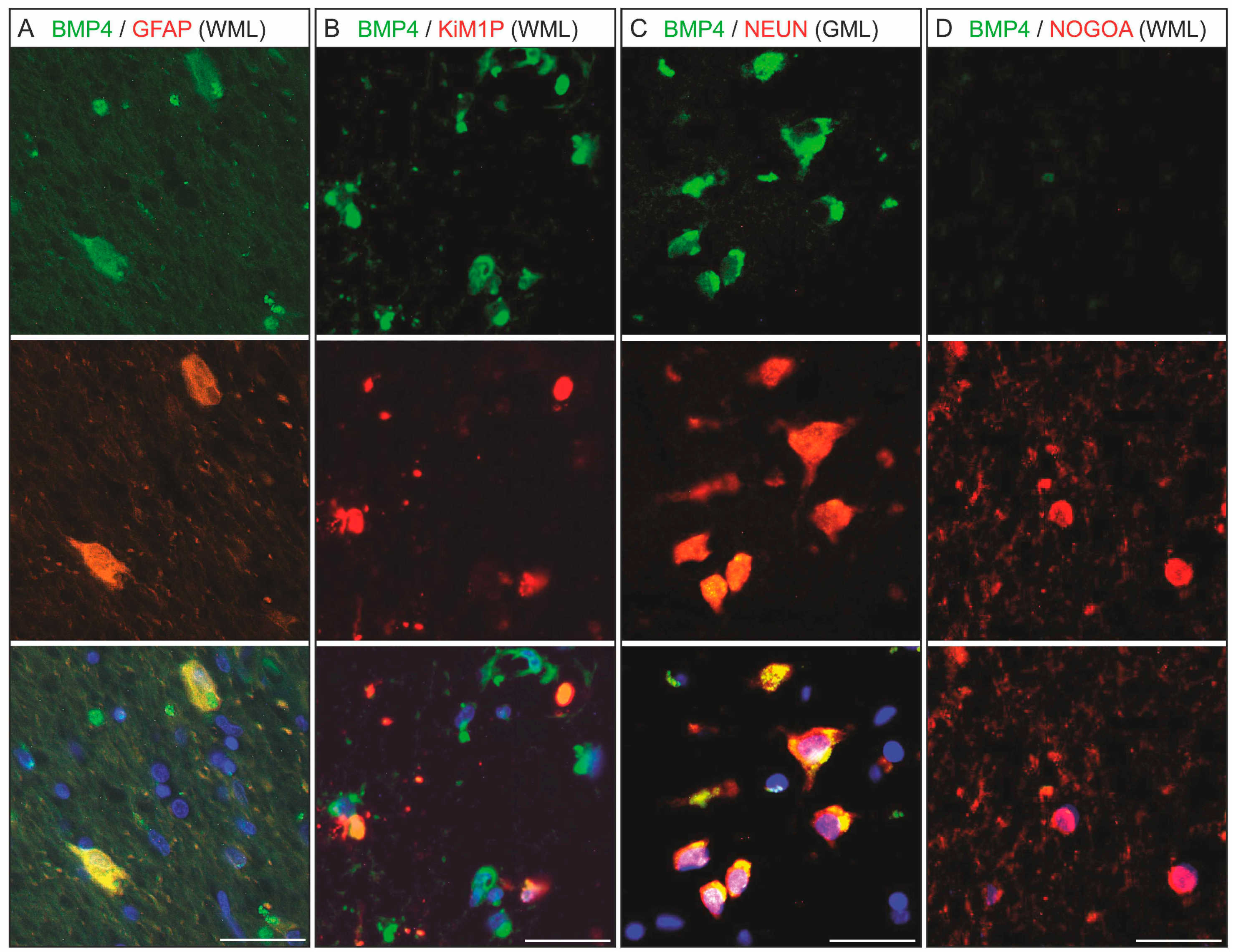
2.2. BMP4 Is Expressed in Astrocytes and Microglial Cells/Macrophages and Its Upregulation Is Associated with Inflammatory Infiltrates
3. Discussion
4. Material and Methods
4.1. Human Brain Tissue
4.2. Histology and Immunohistochemistry
4.3. cDNA Synthesis and Quantitative PCR
4.4. Nanostring
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Crawford, A.H.; Chambers, C.; Franklin, R.J. Remyelination: The true regeneration of the central nervous system. J. Comp. Pathol. 2013, 149, 242–254. [Google Scholar] [CrossRef] [PubMed]
- Hagemeier, K.; Bruck, W.; Kuhlmann, T. Multiple sclerosis—Remyelination failure as a cause of disease progression. Histol. Histopathol. 2012, 27, 277–287. [Google Scholar]
- Irvine, K.A.; Blakemore, W.F. Remyelination protects axons from demyelination-associated axon degeneration. Brain 2008, 131, 1464–1477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funfschilling, U.; Supplie, L.M.; Mahad, D.; Boretius, S.; Saab, A.S.; Edgar, J.; Brinkmann, B.G.; Kassmann, C.M.; Tzvetanova, I.D.; Mobius, W.; et al. Glycolytic oligodendrocytes maintain myelin and long-term axonal integrity. Nature 2012, 485, 517–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapp, B.D.; Peterson, J.; Ransohoff, R.M.; Rudick, R.; Mork, S.; Bo, L. Axonal transection in the lesions of multiple sclerosis. N. Engl. J. Med. 1998, 338, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Trapp, B.D.; Ransohoff, R.; Rudick, R. Axonal pathology in multiple sclerosis: Relationship to neurologic disability. Curr. Opin. Neurol. 1999, 12, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Bo, L. The histopathology of grey matter demyelination in multiple sclerosis. Acta Neurol. Scand. 2009, 120, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Compston, A.; Coles, A. Multiple sclerosis. Lancet 2002, 359, 1221–1231. [Google Scholar] [CrossRef]
- Chang, A.; Tourtellotte, W.W.; Rudick, R.; Trapp, B.D. Premyelinating oligodendrocytes in chronic lesions of multiple sclerosis. N. Engl. J. Med. 2002, 346, 165–173. [Google Scholar] [CrossRef]
- Franklin, R.J.; Ffrench-Constant, C. Remyelination in the CNS: From biology to therapy. Nat. Rev. Neurosci. 2008, 9, 839–855. [Google Scholar] [CrossRef]
- Kotter, M.R.; Stadelmann, C.; Hartung, H.P. Enhancing remyelination in disease—Can we wrap it up? Brain 2011, 134, 1882–1900. [Google Scholar] [CrossRef]
- Kuhlmann, T.; Miron, V.; Cui, Q.; Wegner, C.; Antel, J.; Bruck, W. Differentiation block of oligodendroglial progenitor cells as a cause for remyelination failure in chronic multiple sclerosis. Brain 2008, 131, 1749–1758. [Google Scholar] [CrossRef] [Green Version]
- Kremer, D.; Aktas, O.; Hartung, H.P.; Kury, P. The complex world of oligodendroglial differentiation inhibitors. Ann. Neurol. 2011, 69, 602–618. [Google Scholar] [CrossRef] [PubMed]
- See, J.M.; Grinspan, J.B. Sending mixed signals: Bone morphogenetic protein in myelination and demyelination. J. Neuropathol. Exp. Neurol. 2009, 68, 595–604. [Google Scholar] [CrossRef] [PubMed]
- Ara, J.; See, J.; Mamontov, P.; Hahn, A.; Bannerman, P.; Pleasure, D.; Grinspan, J.B. Bone morphogenetic proteins 4, 6, and 7 are up-regulated in mouse spinal cord during experimental autoimmune encephalomyelitis. J. Neurosci. Res. 2008, 86, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Deininger, M.; Meyermann, R.; Schluesener, H. Detection of two transforming growth factor-beta-related morphogens, bone morphogenetic proteins-4 and -5, in RNA of multiple sclerosis and Creutzfeldt-Jakob disease lesions. Acta Neuropathol. 1995, 90, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Lucchinetti, C.; Bruck, W.; Parisi, J.; Scheithauer, B.; Rodriguez, M.; Lassmann, H. Heterogeneity of multiple sclerosis lesions: Implications for the pathogenesis of demyelination. Ann. Neurol. 2000, 47, 707–717. [Google Scholar] [CrossRef]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Munch, A.E.; Chung, W.S.; Peterson, T.C.; et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef] [Green Version]
- Eixarch, H.; Calvo-Barreiro, L.; Montalban, X.; Espejo, C. Bone morphogenetic proteins in multiple sclerosis: Role in neuroinflammation. Brain Behav. Immun. 2018, 68, 1–10. [Google Scholar] [CrossRef]
- Kuczma, M.; Kurczewska, A.; Kraj, P. Modulation of bone morphogenic protein signaling in T-cells for cancer immunotherapy. J. Immunotoxicol. 2014, 11, 319–327. [Google Scholar] [CrossRef]
- Voumvourakis, K.I.; Antonelou, R.C.; Kitsos, D.K.; Stamboulis, E.; Tsiodras, S. TGF-beta/BMPs: Crucial crossroad in neural autoimmune disorders. Neurochem. Int. 2011, 59, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Nohe, A.; Hassel, S.; Ehrlich, M.; Neubauer, F.; Sebald, W.; Henis, Y.I.; Knaus, P. The mode of bone morphogenetic protein (BMP) receptor oligomerization determines different BMP-2 signaling pathways. J. Biol. Chem. 2002, 277, 5330–5338. [Google Scholar] [CrossRef] [PubMed]
- Mabie, P.C.; Mehler, M.F.; Marmur, R.; Papavasiliou, A.; Song, Q.; Kessler, J.A. Bone morphogenetic proteins induce astroglial differentiation of oligodendroglial-astroglial progenitor cells. J. Neurosci. 1997, 17, 4112–4120. [Google Scholar] [CrossRef] [PubMed]
- Grinspan, J.B.; Edell, E.; Carpio, D.F.; Beesley, J.S.; Lavy, L.; Pleasure, D.; Golden, J.A. Stage-specific effects of bone morphogenetic proteins on the oligodendrocyte lineage. J. Neurobiol. 2000, 43, 1–17. [Google Scholar] [CrossRef]
- Gomes, W.A.; Mehler, M.F.; Kessler, J.A. Transgenic overexpression of BMP4 increases astroglial and decreases oligodendroglial lineage commitment. Dev. Biol. 2003, 255, 164–177. [Google Scholar] [CrossRef] [Green Version]
- Miller, R.H.; Dinsio, K.; Wang, R.; Geertman, R.; Maier, C.E.; Hall, A.K. Patterning of spinal cord oligodendrocyte development by dorsally derived BMP4. J. Neurosci. Res. 2004, 76, 9–19. [Google Scholar] [CrossRef]
- See, J.; Zhang, X.; Eraydin, N.; Mun, S.B.; Mamontov, P.; Golden, J.A.; Grinspan, J.B. Oligodendrocyte maturation is inhibited by bone morphogenetic protein. Mol. Cell. Neurosci. 2004, 26, 481–492. [Google Scholar] [CrossRef]
- Guo, X.; Wang, X.F. Signaling cross-talk between TGF-beta/BMP and other pathways. Cell Res. 2009, 19, 71–88. [Google Scholar] [CrossRef]
- Gazzerro, E.; Minetti, C. Potential drug targets within bone morphogenetic protein signaling pathways. Curr. Opin. Pharmacol. 2007, 7, 325–333. [Google Scholar] [CrossRef]
- Krause, C.; Guzman, A.; Knaus, P. Noggin. Int. J. Biochem. Cell Biol. 2011, 43, 478–481. [Google Scholar] [CrossRef]
- Zimmerman, L.B.; De Jesus-Escobar, J.M.; Harland, R.M. The Spemann organizer signal noggin binds and inactivates bone morphogenetic protein 4. Cell 1996, 86, 599–606. [Google Scholar] [CrossRef]
- Song, K.; Krause, C.; Shi, S.; Patterson, M.; Suto, R.; Grgurevic, L.; Vukicevic, S.; van Dinther, M.; Falb, D.; Ten, D.P.; et al. Identification of a key residue mediating bone morphogenetic protein (BMP)-6 resistance to noggin inhibition allows for engineered BMPs with superior agonist activity. J. Biol. Chem. 2010, 285, 12169–12180. [Google Scholar] [CrossRef] [PubMed]
- Groppe, J.; Greenwald, J.; Wiater, E.; Rodriguez-Leon, J.; Economides, A.N.; Kwiatkowski, W.; Affolter, M.; Vale, W.W.; Izpisua Belmonte, J.C.; Choe, S. Structural basis of BMP signalling inhibition by the cystine knot protein Noggin. Nature 2002, 420, 636–642. [Google Scholar] [CrossRef] [PubMed]
- Cate, H.S.; Sabo, J.K.; Merlo, D.; Kemper, D.; Aumann, T.D.; Robinson, J.; Merson, T.D.; Emery, B.; Perreau, V.M.; Kilpatrick, T.J. Modulation of bone morphogenic protein signalling alters numbers of astrocytes and oligodendroglia in the subventricular zone during cuprizone-induced demyelination. J. Neurochem. 2010, 115, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabo, J.K.; Aumann, T.D.; Merlo, D.; Kilpatrick, T.J.; Cate, H.S. Remyelination is altered by bone morphogenic protein signaling in demyelinated lesions. J. Neurosci. 2011, 31, 4504–4510. [Google Scholar] [CrossRef] [PubMed]
- See, J.; Mamontov, P.; Ahn, K.; Wine-Lee, L.; Crenshaw, E.B., III; Grinspan, J.B. BMP signaling mutant mice exhibit glial cell maturation defects. Mol. Cell. Neurosci. 2007, 35, 171–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjelobaba, I.; Begovic-Kupresanin, V.; Pekovic, S.; Lavrnja, I. Animal models of multiple sclerosis: Focus on experimental autoimmune encephalomyelitis. J. Neurosci. Res. 2018, 96, 1021–1042. [Google Scholar] [CrossRef] [PubMed]
- Blaschuk, K.L.; Frost, E.E.; Ffrench-Constant, C. The regulation of proliferation and differentiation in oligodendrocyte progenitor cells by alphaV integrins. Development 2000, 127, 1961–1969. [Google Scholar] [PubMed]
- Buttery, P.C.; Ffrench-Constant, C. Laminin-2/integrin interactions enhance myelin membrane formation by oligodendrocytes. Mol. Cell. Neurosci. 1999, 14, 199–212. [Google Scholar] [CrossRef]
- Relvas, J.B.; Setzu, A.; Baron, W.; Buttery, P.C.; LaFlamme, S.E.; Franklin, R.J.; Ffrench-Constant, C. Expression of dominant-negative and chimeric subunits reveals an essential role for beta1 integrin during myelination. Curr. Biol. 2001, 11, 1039–1043. [Google Scholar] [CrossRef]
- Gutowski, N.J.; Newcombe, J.; Cuzner, M.L. Tenascin-R and C in multiple sclerosis lesions: Relevance to extracellular matrix remodelling. Neuropathol. Appl. Neurobiol. 1999, 25, 207–214. [Google Scholar] [CrossRef]
- Sobel, R.A.; Chen, M.; Maeda, A.; Hinojoza, J.R. Vitronectin and integrin vitronectin receptor localization in multiple sclerosis lesions. J. Neuropathol. Exp. Neurol. 1995, 54, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Ghiani, C.A.; Eisen, A.M.; Yuan, X.; DePinho, R.A.; McBain, C.J.; Gallo, V. Neurotransmitter receptor activation triggers p27(Kip1 )and p21(CIP1) accumulation and G1 cell cycle arrest in oligodendrocyte progenitors. Development 1999, 126, 1077–1090. [Google Scholar] [PubMed]
- Steinhauser, C.; Gallo, V. News on glutamate receptors in glial cells. Trends Neurosci. 1996, 19, 339–345. [Google Scholar] [CrossRef]
- Gottle, P.; Kremer, D.; Jander, S.; Odemis, V.; Engele, J.; Hartung, H.P.; Kury, P. Activation of CXCR7 receptor promotes oligodendroglial cell maturation. Ann. Neurol. 2010, 68, 915–924. [Google Scholar] [CrossRef] [PubMed]
- John, G.R.; Shankar, S.L.; Shafit-Zagardo, B.; Massimi, A.; Lee, S.C.; Raine, C.S.; Brosnan, C.F. Multiple sclerosis: Re-expression of a developmental pathway that restricts oligodendrocyte maturation. Nat. Med. 2002, 8, 1115–1121. [Google Scholar] [CrossRef]
- Fancy, S.P.; Baranzini, S.E.; Zhao, C.; Yuk, D.I.; Irvine, K.A.; Kaing, S.; Sanai, N.; Franklin, R.J.; Rowitch, D.H. Dysregulation of the Wnt pathway inhibits timely myelination and remyelination in the mammalian CNS. Genes Dev. 2009, 23, 1571–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charles, P.; Reynolds, R.; Seilhean, D.; Rougon, G.; Aigrot, M.S.; Niezgoda, A.; Zalc, B.; Lubetzki, C. Re-expression of PSA-NCAM by demyelinated axons: An inhibitor of remyelination in multiple sclerosis? Brain 2002, 125, 1972–1979. [Google Scholar] [CrossRef]
- Back, S.A.; Tuohy, T.M.; Chen, H.; Wallingford, N.; Craig, A.; Struve, J.; Luo, N.L.; Banine, F.; Liu, Y.; Chang, A.; et al. Hyaluronan accumulates in demyelinated lesions and inhibits oligodendrocyte progenitor maturation. Nat. Med. 2005, 11, 966–972. [Google Scholar] [CrossRef]
- Kotter, M.R.; Setzu, A.; Sim, F.J.; van, R.N.; Franklin, R.J. Macrophage depletion impairs oligodendrocyte remyelination following lysolecithin-induced demyelination. Glia 2001, 35, 204–212. [Google Scholar] [CrossRef]
- Bieber, A.J.; Kerr, S.; Rodriguez, M. Efficient central nervous system remyelination requires T cells. Ann. Neurol. 2003, 53, 680–684. [Google Scholar] [CrossRef] [PubMed]
- Foote, A.K.; Blakemore, W.F. Inflammation stimulates remyelination in areas of chronic demyelination. Brain 2005, 128, 528–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schindelin, J.; Rueden, C.T.; Hiner, M.C.; Eliceiri, K.W. The ImageJ ecosystem: An open platform for biomedical image analysis. Mol. Reprod. Dev. 2015, 82, 518–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voigt, D.; Scheidt, U.; Derfuss, T.; Bruck, W.; Junker, A. Expression of the Antioxidative Enzyme Peroxiredoxin 2 in Multiple Sclerosis Lesions in Relation to Inflammation. Int. J. Mol. Sci. 2017, 18, 760. [Google Scholar] [CrossRef] [PubMed]
- Junker, A.; Krumbholz, M.; Eisele, S.; Mohan, H.; Augstein, F.; Bittner, R.; Lassmann, H.; Wekerle, H.; Hohlfeld, R.; Meinl, E. MicroRNA profiling of multiple sclerosis lesions identifies modulators of the regulatory protein CD47. Brain 2009, 132, 3342–3352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, R.F.; Werner, R.; Vollbrecht, C.; Hager, T.; Flom, E.; Christoph, D.C.; Schmeller, J.; Schmid, K.W.; Wohlschlaeger, J.; Mairinger, F.D. ACTB, CDKN1B, GAPDH, GRB2, RHOA and SDCBP Were Identified as Reference Genes in Neuroendocrine Lung Cancer via the nCounter Technology. PLoS ONE 2016, 11, e0165181. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case | Age | Sex | Cause of Death | Inflammatory Circumstannces Perimortal |
|---|---|---|---|---|
| Multiple Sclerosis—autopsy cases | ||||
| MS-01 | 66 | female | cancer metastases in the liver resulting in severe failure of the liver functions | no |
| MS-02 | 75 | female | pneumonia | pneumonia |
| MS-03 | 68 | female | pneumonia | pneumonia |
| MS-04 | 78 | female | stroke | no |
| MS-05 | 49 | male | pneumonia | pneumonia |
| MS-06 | 55 | male | Respiratory insufficiency by pneumonia and urosepsis | pneumonia and urosepsis |
| MS-07 | 44 | male | pneumonia by aspiration | pneumonia |
| MS-08 | 44 | male | multiorgan failure | unkown |
| MS-09 + MS-10 | 57 | female | respiratory insufficiency | (uro)sepsis |
| MS-11 | 53 | male | Euthanasia | unknown |
| MS-12 | 62 | female | cachexia and pulmonary insufficiency | no |
| MS-13 + MS-14 | 56 | female | respiratory insufficiency by Pneumonia | pneumonia |
| MS-15 | 54 | female | heart failure | unknown |
| MS-16 | 58 | male | terminal renal insufficiency | pneumonia |
| MS-17 | 63 | male | pneumonia | pneumonia |
| MS-18 | 48 | female | respiratory failure | Unknown |
| Multiple Sclerosis—biopsy cases | ||||
| MS-19 | 48 | male | ||
| MS-20 | 46 | female | ||
| MS-21 | 57 | female | ||
| MS-22 | 36 | female | ||
| MS-23 | 47 | male | ||
| MS-24 | 43 | male | ||
| MS-25 | 61 | female | ||
| MS-26 | 46 | female | ||
| Controls—autopsy cases | ||||
| CON-01 | 46 | female | haemorrhagic shock, sepsis | sepsis |
| CON-02 | 55 | male | aortic dissection | pancreatitis |
| CON-03 | 56 | female | pulmonary embolism | no |
| CON-04 | 56 | male | hepatic insufficiency after liver transplantation | no |
| CON-05 | 58 | female | cardiogenic shock | no |
| CON-06 | 63 | male | pulmonary insufficiency in pneumonia | pneumonia |
| CON-07 | 66 | female | multiorgan failure | no |
| CON-08 | 68 | female | cardiorespiratory insufficiency | no |
| Antigen | Company | Pre-Treatment | Dilution |
|---|---|---|---|
| BMP4, ab39973, rabbit pc | Abcam | citrate | 1:60 |
| Nogo A,sc-25660,clone H-300, rabbit pc | SantaCruz Biotechnology | citrate | 1:100 |
| CD8, clone C8/144B, mouse mc | DAKO | citrate | 1:150 |
| CD3,cloneSP7, mouse mc | DCS | citrate | 1:100 |
| C3D, A0063 | DAKO | citrate | 1:400 |
| S100A10,MA5-15326, clone 4E7E10, mouse mc | ThermoFisher | citrate | 1:1000 |
| KiM1P (CD68) | kind gift from Prof. Klapper, Institute for Pathology, Kiel, Germany | none | 1:10.000 |
| MBP, REF A0623, rabbit pc | DAKO | none | 1:1000 |
| Noggin, ab16054, rabbit pc NeuN, ab104225 | Abcam Abcam | Citrate Citrate | 1:800 1:1000 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harnisch, K.; Teuber-Hanselmann, S.; Macha, N.; Mairinger, F.; Fritsche, L.; Soub, D.; Meinl, E.; Junker, A. Myelination in Multiple Sclerosis Lesions Is Associated with Regulation of Bone Morphogenetic Protein 4 and Its Antagonist Noggin. Int. J. Mol. Sci. 2019, 20, 154. https://doi.org/10.3390/ijms20010154
Harnisch K, Teuber-Hanselmann S, Macha N, Mairinger F, Fritsche L, Soub D, Meinl E, Junker A. Myelination in Multiple Sclerosis Lesions Is Associated with Regulation of Bone Morphogenetic Protein 4 and Its Antagonist Noggin. International Journal of Molecular Sciences. 2019; 20(1):154. https://doi.org/10.3390/ijms20010154
Chicago/Turabian StyleHarnisch, Kim, Sarah Teuber-Hanselmann, Nicole Macha, Fabian Mairinger, Lena Fritsche, Daniel Soub, Edgar Meinl, and Andreas Junker. 2019. "Myelination in Multiple Sclerosis Lesions Is Associated with Regulation of Bone Morphogenetic Protein 4 and Its Antagonist Noggin" International Journal of Molecular Sciences 20, no. 1: 154. https://doi.org/10.3390/ijms20010154
APA StyleHarnisch, K., Teuber-Hanselmann, S., Macha, N., Mairinger, F., Fritsche, L., Soub, D., Meinl, E., & Junker, A. (2019). Myelination in Multiple Sclerosis Lesions Is Associated with Regulation of Bone Morphogenetic Protein 4 and Its Antagonist Noggin. International Journal of Molecular Sciences, 20(1), 154. https://doi.org/10.3390/ijms20010154