ConA-Like Lectins: High Similarity Proteins as Models to Study Structure/Biological Activities Relationships


Abstract
:1. Introduction
2. Carbohydrate-Specificity Overview
3. Overview of ConA-Like Lectins Purification
4. Biological Activities of ConA-Like Lectins
4.1. Inflammatory and Nociceptive
4.2. Vasoactive
4.3. Antiproliferative
4.4. Antibacterial and Antifungal
4.5. Mitogenic
4.6. Immunomodulatory
4.7. Antidepressant and Neuroprotective
4.8. Insecticidal
5. Structure of ConA-like Lectins
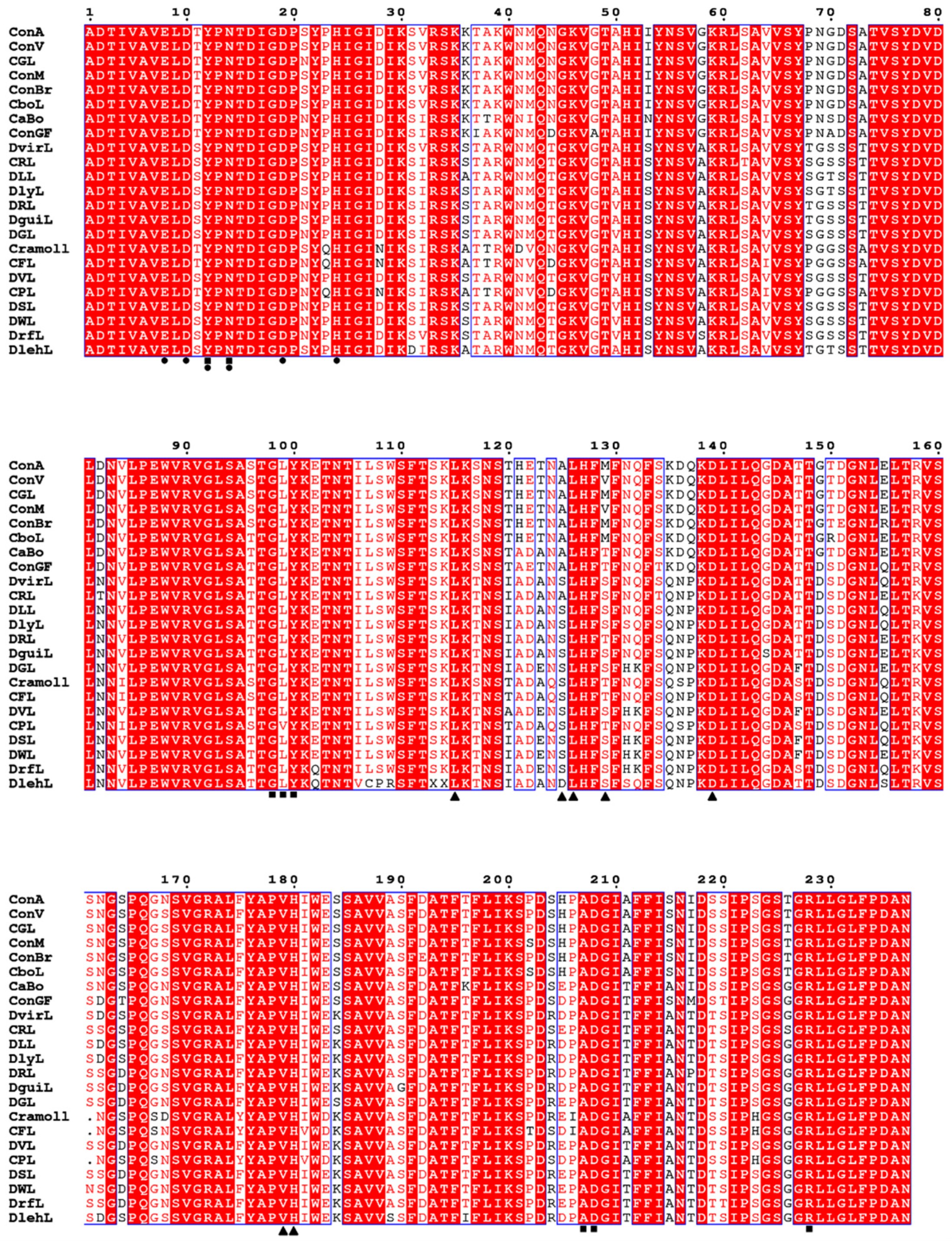
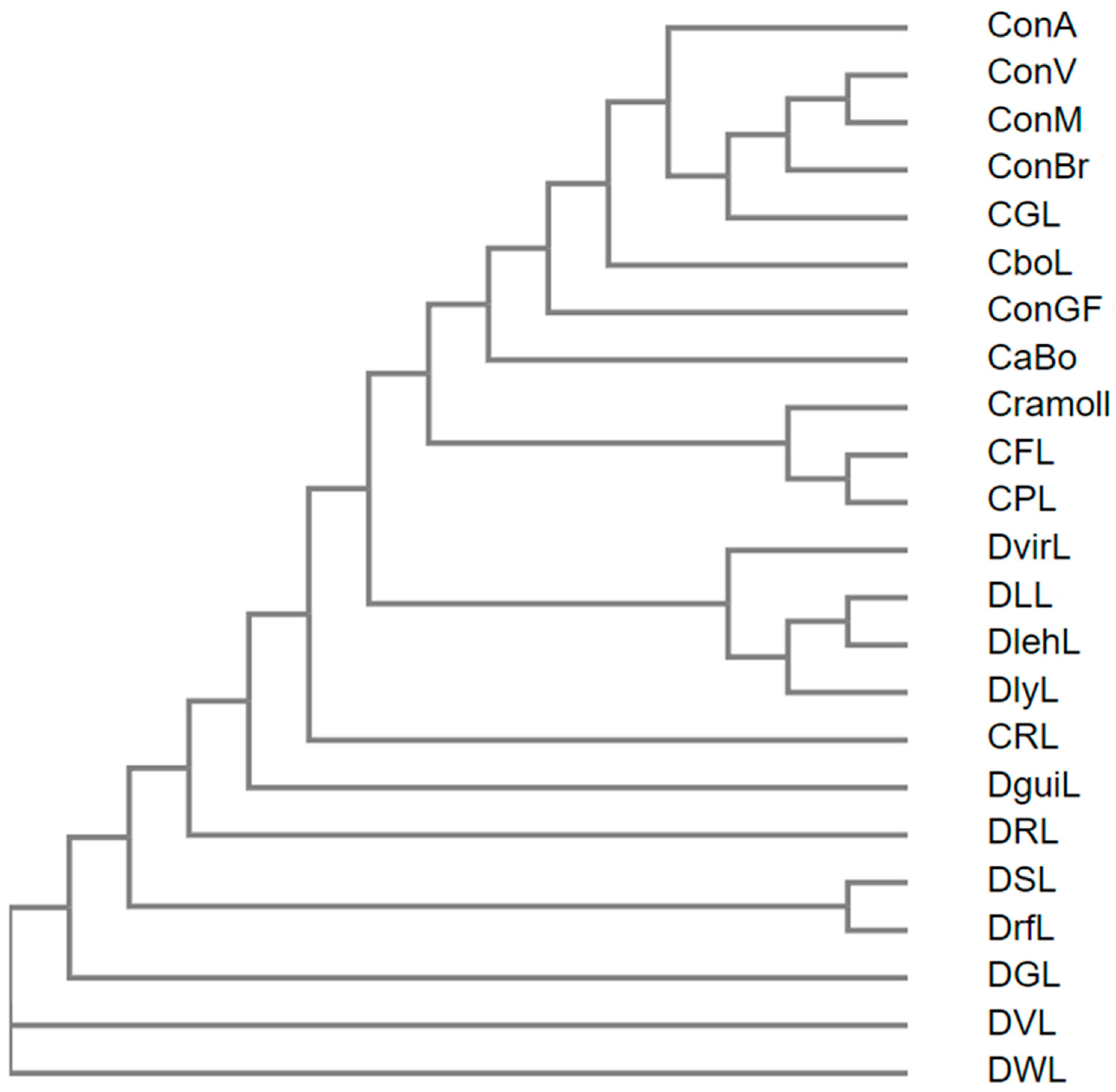
5.1. Primary Structure
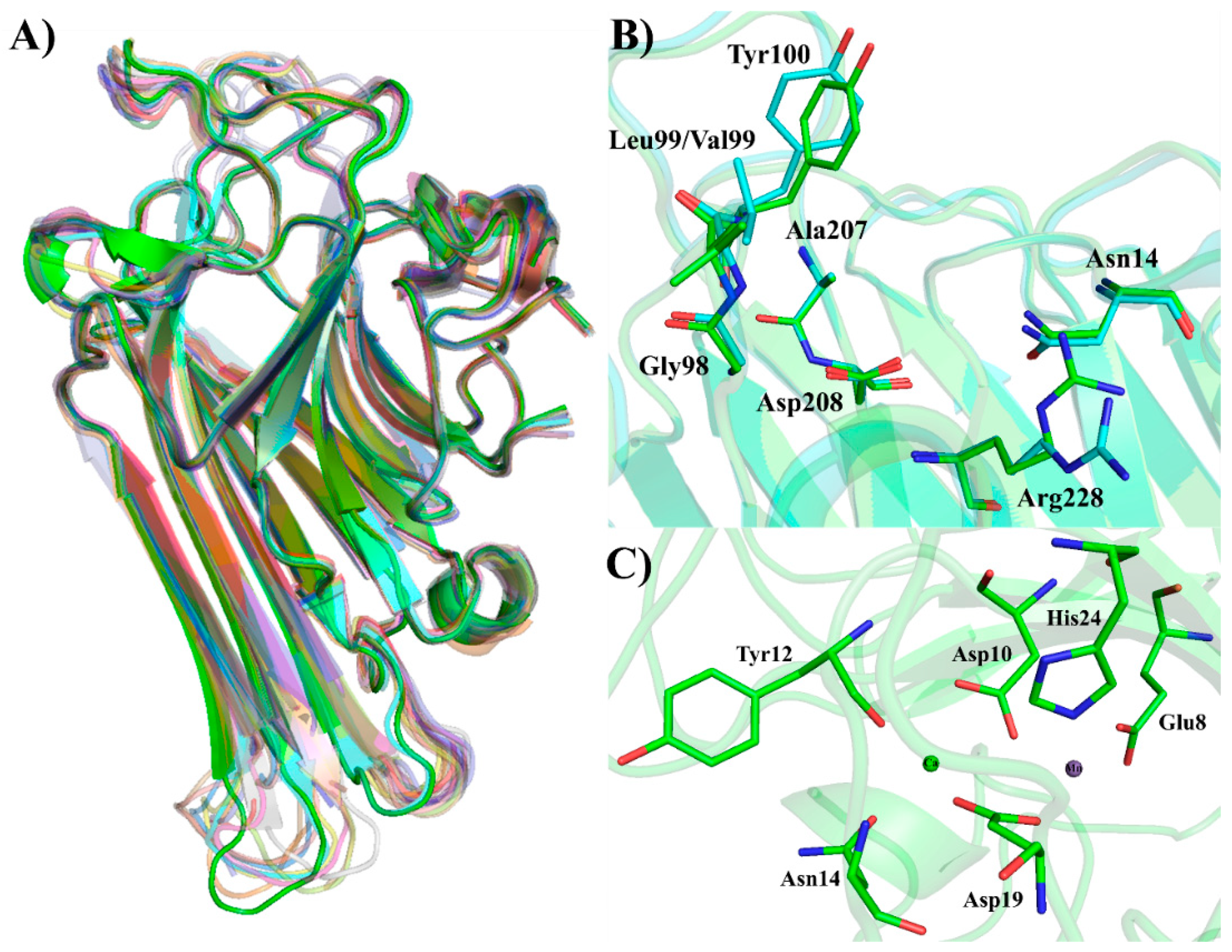
5.2. Three-Dimensional Structure
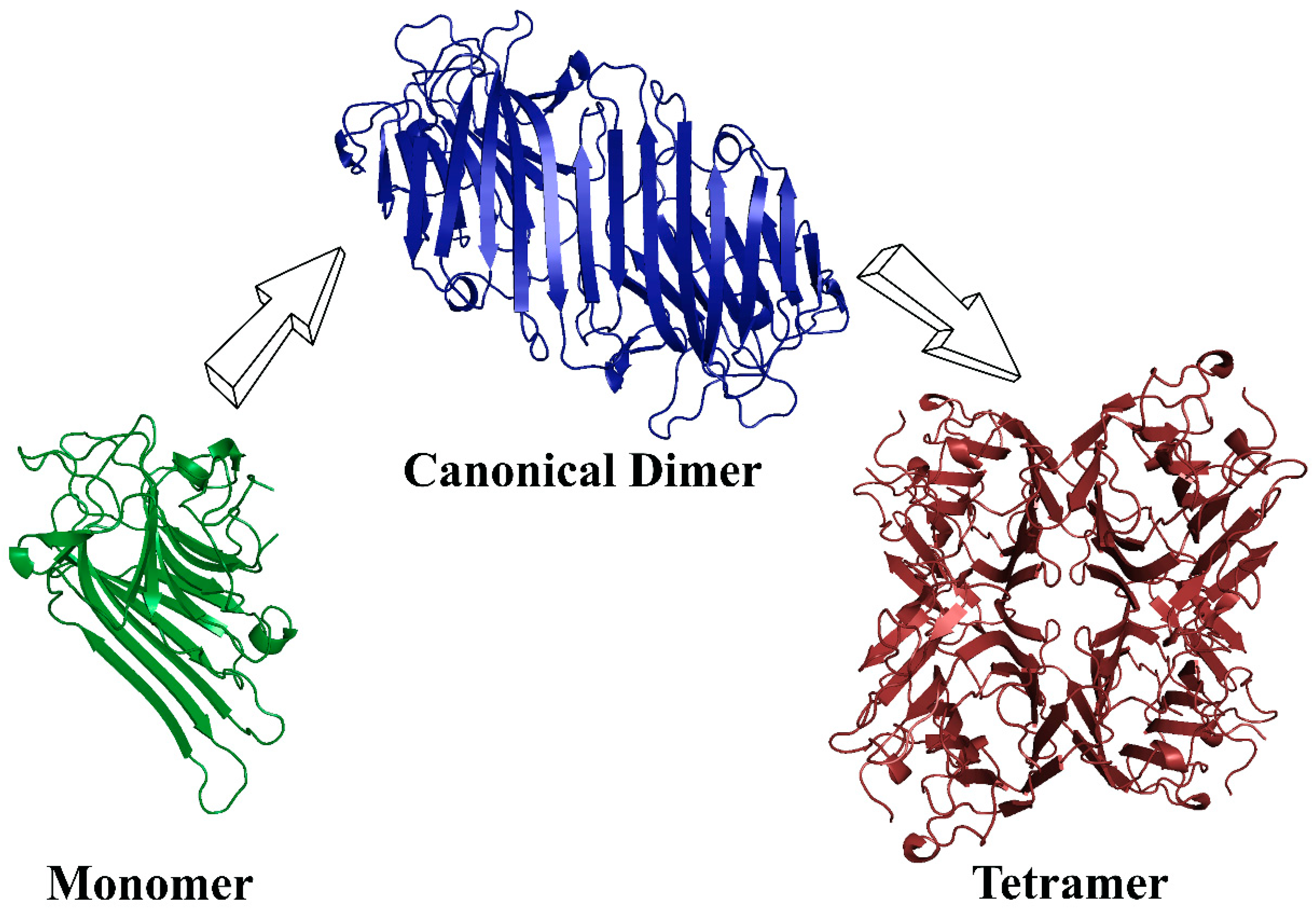
5.3. Quaternary Structure
6. Physicochemical Properties
7. Structure/Biological Activities Relationships
8. Conclusions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ConA | Canavalia ensiformis lectin |
| CRD | Carbohydrate-recognition domain |
| Man | Mannose |
| SBA | Soybean agglutinin |
| ConGF | Canavalia grandiflora lectin |
| PEG | Polyethylene glycol |
| CGL | Canavalia gladiata lectin |
| ConM | Canavalia maritima lectin |
| ConBr | Canavalia brasiliensis lectin |
| ConV | Canavalia virosa lectin |
| CoxyL | Canavalia oxyphylla lectin |
| CvilL | Canavalia villosa lectin |
| DRL | Dioclea rostrata lectin |
| DWL | Dioclea wilsonii lectin |
| DvirL | Dioclea virgata lectin |
| DlyL | Dioclea lasiophylla lectin |
| DrfL | Dioclea reflexa lectin |
| CRL I | Cymbosema roseum lectin |
| DguiL | Dioclea guianensis lectin |
| DVL | Dioclea violacea lectin |
| CFL | Cratylia floribunda lectin |
| L-NAME | N(ω)-nitro-L-arginine methyl ester |
| DSL | Dioclea sclerocarpa lectin |
| eNOS | Endothelial nitric oxide synthase |
| DLL | Dioclea lasiocarpa lectin |
| IFN-γ | Interferon-γ |
| IL | Interleukin |
| TNF-α | Tumor Necrosis Factor-α |
| NMDA | N-methyl-D-aspartate |
| DlehL | Dioclea lehmani lectin |
| SDS | Sodium dodecyl sulfate |
| HS | Hydophobic site |
| MBS | Metal-binding site |
| CaBo | Canavalia bonariensis lectin |
| ConBol | Canavalia boliviana lectin |
| Cramoll | Cratylia mollis lectin |
| CPL | Camptosema pedicallatum lectin |
| CNS | Central Nervous System |
| NO | Nitric Oxide |
References
- Kilpatrick, D.C. Animal lectins: A historical introduction and overview. Biochim. Biophys. Acta 2002, 1572, 187–197. [Google Scholar] [CrossRef]
- Van Breedam, W.; Pöhlmann, S.; Favoreel, H.W.; de Groot, R.J.; Nauwynck, H.J. Bitter-sweet symphony: Glycan-lectin interactions in virus biology. FEMS Microbiol. Rev. 2014, 38, 598–632. [Google Scholar] [CrossRef] [PubMed]
- Peumans, W.J.; Van Damme, E.J. Lectins as plant defense proteins. Plant Physiol. 1995, 109, 347–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lannoo, N.; Van Damme, E.J.M. Lectin domains at the frontiers of plant defense. Front. Plant Sci. 2014, 5, 397. [Google Scholar] [CrossRef] [PubMed]
- Mann, K.; Farias, C.M.; Del Sol, F.G.; Santos, C.F.; Grangeiro, T.B.; Nagano, C.S.; Cavada, B.S.; Calvete, J.J. The amino-acid sequence of the glucose/mannose-specific lectin isolated from Parkia platycephala seeds reveals three tandemly arranged jacalin-related domains. Eur. J. Biochem. 2001, 268, 4414–4422. [Google Scholar] [CrossRef] [Green Version]
- Lackey, J.A. Phaseolae. In Advances in Legume Systematics; HMSO: London, UK, 1981; pp. 301–327. [Google Scholar]
- Varela, E.S.; Lima, J.P.M.S.; Galdino, A.S.; da Silva Pinto, L.; Bezerra, W.M.; Nunes, E.P.; Alves, M.A.O.; Grangeiro, T.B. Relationships in subtribe Diocleinae (Leguminosae; Papilionoideae) inferred from internal transcribed spacer sequences from nuclear ribosomal DNA. Phytochemistry 2004, 65, 59–69. [Google Scholar] [CrossRef]
- Cavada, B.S.; Barbosa, T.; Arruda, S.; Grangeiro, T.B.; Barral-Netto, M. Revisiting proteus: Do minor changes in lectin structure matter in biological activity? Lessons from and potential biotechnological uses of the Diocleinae subtribe lectins. Curr. Protein Pept. Sci. 2001, 2, 123–135. [Google Scholar] [CrossRef]
- Li, C.-Y.; Xu, H.-L.; Liu, B.; Bao, J.-K. Concanavalin A, from an old protein to novel candidate anti-neoplastic drug. Curr. Mol. Pharmacol. 2010, 3, 123–128. [Google Scholar] [CrossRef]
- Lagarda-Diaz, I.; Guzman-Partida, A.M.; Vazquez-Moreno, L. Legume Lectins: Proteins with Diverse Applications. Int. J. Mol. Sci. 2017, 18, 1242. [Google Scholar] [CrossRef]
- Marques, G.F.O.; Osterne, V.J.S.; Almeida, L.M.; Oliveira, M.V.; Brizeno, L.A.C.; Pinto-Junior, V.R.; Santiago, M.Q.; Neco, A.H.B.; Mota, M.R.L.; Souza, L.A.G.; et al. Contribution of the carbohydrate-binding ability of Vatairea guianensis lectin to induce edematogenic activity. Biochimie 2017, 140, 58–65. [Google Scholar] [CrossRef]
- Nagano, C.S.; Calvete, J.J.; Barettino, D.; Pérez, A.; Cavada, B.S.; Sanz, L. Insights into the structural basis of the pH-dependent dimer–tetramer equilibrium through crystallographic analysis of recombinant Diocleinae lectins. Biochem. J. 2008, 409, 417–428. [Google Scholar] [CrossRef] [PubMed]
- Ramos, M.V.; Grangeiro, T.B.; Cavada, B.S.; Shepherd, I.; de Melo Lopes, R.O.; Sampaio, A.H. Carbohydrate/glycan-binding specificity of legume lectins in respect to their proposed biological functions. Braz. Arch. Biol. Technol. 2000, 43, 349–359. [Google Scholar] [CrossRef]
- Dam, T.K.; Cavada, B.S.; Grangeiro, T.B.; Santos, C.F.; de Sousa, F.A.; Oscarson, S.; Brewer, C.F. Diocleinae lectins are a group of proteins with conserved binding sites for the core trimannoside of asparagine-linked oligosaccharides and differential specificities for complex carbohydrates. J. Biol. Chem. 1998, 273, 12082–12088. [Google Scholar] [CrossRef] [PubMed]
- Maupin, K.A.; Liden, D.; Haab, B.B. The fine specificity of mannose-binding and galactose-binding lectins revealed using outlier motif analysis of glycan array data. Glycobiology 2012, 22, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Ramos, M.V.; Cavada, B.S.; Mazard, A.-M.; Rougé, P. Interaction of Diocleinae lectins with Glycoproteins Based in Surface Plasmon Resonance. Mem. Inst. Oswaldo Cruz 2002, 97, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Osterne, V.J.S.; Santiago, M.Q.; Pinto-Junior, V.R.; Cajazeiras, J.B.; Correia, J.L.A.; Leitão, C.C.F.; Carneiro, R.F.; Pereira-Junior, F.N.; Vasconcelos, M.A.; Rocha, B.A.M.; et al. Purification, partial characterization, and CNBr-sepharose immobilization of a vasorelaxant glucose/mannose lectin from Canavalia virosa seeds. Appl. Biochem. Biotechnol. 2014, 172, 3342–3353. [Google Scholar] [CrossRef] [PubMed]
- Pinto-Júnior, V.R.; de Santiago, M.Q.; da Silva Osterne, V.J.; Correia, J.L.A.; Pereira-Júnior, F.N.; Cajazeiras, J.B.; de Vasconcelos, M.A.; Teixeira, E.H.; do Nascimento, A.S.F.; Miguel, T.B.A.R.; et al. Purification, partial characterization and immobilization of a mannose-specific lectin from seeds of Dioclea lasiophylla mart. Molecules 2013, 18, 10857–10869. [Google Scholar] [CrossRef]
- Santiago, M.Q.; Leitão, C.C.F.; Pereira, F.N., Jr.; Pinto, V.R., Jr.; Osterne, V.J.S.; Lossio, C.F.; Cajazeiras, J.B.; Silva, H.C.; Arruda, F.V.S.; Pereira, L.P.; et al. Purification, characterization and partial sequence of a pro-inflammatory lectin from seeds of Canavalia oxyphylla Standl. & L. O. Williams. J. Mol. Recognit. 2014, 27, 117–123. [Google Scholar]
- Lossio, C.F.; Moreira, C.G.; Amorim, R.M.F.; Nobre, C.S.; Silva, M.T.L.; Neto, C.C.; Pinto-Junior, V.R.; Silva, I.B.; Campos, J.; Assreuy, A.M.S.; et al. Lectin from Canavalia villosa seeds: A glucose/mannose-specific protein and a new tool for inflammation studies. Int. J. Biol. Macromol. 2017, 105, 272–280. [Google Scholar] [CrossRef]
- Nascimento, K.S.; Cunha, A.I.; Nascimento, K.S.; Cavada, B.S.; Azevedo, A.M.; Aires-Barros, M.R. An overview of lectins purification strategies. J. Mol. Recognit. 2012, 25, 527–541. [Google Scholar] [CrossRef]
- Flanagan, S.D.; Barondes, S.H. Affinity partitioning. A method for purification of proteins using specific polymer-ligands in aqueous polymer two-phase systems. J. Biol. Chem. 1975, 250, 1484–1489. [Google Scholar] [PubMed]
- Soares, P.A.G.; Nascimento, C.O.; Porto, T.S.; Correia, M.T.S.; Porto, A.L.F.; Carneiro-da-Cunha, M.G. Purification of a lectin from Canavalia ensiformis using PEG–citrate aqueous two-phase system. J. Chromatogr. B 2011, 879, 457–460. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, K.S.; Rosa, P.A.J.; Nascimento, K.S.; Cavada, B.S.; Azevedo, A.M.; Aires-Barros, M.R. Partitioning and recovery of Canavalia brasiliensis lectin by aqueous two-phase systems using design of experiments methodology. Sep. Purif. Technol. 2010, 75, 48–54. [Google Scholar] [CrossRef]
- Nascimento, K.S.; Azevedo, A.M.; Cavada, B.S.; Aires-Barros, M.R. Partitioning of Canavalia brasiliensis Lectin in Polyethylene Glycol—Sodium Citrate Aqueous Two-Phase Systems. Sep. Sci. Technol. 2010, 45, 2180–2186. [Google Scholar] [CrossRef]
- Porto, C.S.; Porto, T.S.; Nascimento, K.S.; Teixeira, E.H.; Cavada, B.S.; Lima-Filho, J.L.; Porto, A.L.F. Partition of lectin from Canavalia grandiflora Benth in aqueous two-phase systems using factorial design. Biochem. Eng. J. 2011, 53, 165–171. [Google Scholar] [CrossRef]
- Guo, W. A new matrix for membrane affinity chromatography and its application to the purification of concanavalin A. J. Memb. Sci. 2001, 182, 227–234. [Google Scholar] [CrossRef]
- Assreuy, A.M.; Shibuya, M.D.; Martins, G.J.; De Souza, M.L.; Cavada, B.S.; Moreira, R.A.; Oliveira, J.T.; Ribeiro, R.A.; Flores, C.A. Anti-inflammatory effect of glucose-mannose binding lectins isolated from Brazilian beans. Mediat. Inflamm. 1997, 6, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Assreuy, A.M.S.; Fontenele, S.R.; de Freitas Pires, A.; Fernandes, D.C.; Rodrigues, N.V.F.C.; Bezerra, E.H.S.; Moura, T.R.; do Nascimento, K.S.; Cavada, B.S. Vasodilator effects of Diocleinae lectins from the Canavalia genus. Naunyn-Schmiedeberg’s. Arch. Pharmacol. 2009, 380, 509–521. [Google Scholar] [CrossRef] [PubMed]
- Pinto, N.V.; Santos, C.F.; Cavada, B.S.; do Nascimento, K.S.; Pereira Junior, F.N.; de Freitas Pires, A.; Assreuy, A.M.S. Homologous Canavalia lectins elicit different patterns of antinociceptive responses. Nat. Prod. Commun. 2013, 8, 1621–1624. [Google Scholar]
- Bento, C.A.M.; Cavada, B.S.; Oliveira, J.T.A.; Moreira, R.A.; Barja-Fidalgo, C. Rat paw edema and leukocyte immigration induced by plant lectins. Agents Actions 1993, 38, 48–54. [Google Scholar] [CrossRef]
- Assreuy, A.M.; Martins, G.J.; Moreira, M.E.; Brito, G.A.; Cavada, B.S.; Ribeiro, R.A.; Flores, C.A. Prevention of cyclophosphamide-induced hemorrhagic cystitis by glucose-mannose binding plant lectins. J. Urol. 1999, 161, 1988–1993. [Google Scholar] [CrossRef]
- Alencar, N.M.; Teixeira, E.H.; Assreuy, A.M.; Cavada, B.S.; Flores, C.A.; Ribeiro, R.A. Leguminous lectins as tools for studying the role of sugar residues in leukocyte recruitment. Mediat. Inflamm. 1999, 8, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Bevilacqua, M.P.; Nelson, R.M. Selectins. J. Clin. Investig. 1993, 91, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Simões, R.C.; Rocha, B.A.M.; Bezerra, M.J.B.; Barroso-Neto, I.L.; Pereira-Junior, F.N.; da Mata Moura, R.; do Nascimento, K.S.; Nagano, C.S.; Delatorre, P.; de Freitas Pires, A.; et al. Protein crystal content analysis by mass spectrometry and preliminary X-ray diffraction of a lectin from Canavalia grandiflora seeds with modulatory role in inflammation. Rapid Commun. Mass Spectrom. 2012, 26, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Osterne, V.J.S.; Silva-Filho, J.C.; Santiago, M.Q.; Pinto-Junior, V.R.; Almeida, A.C.; Barreto, A.A.G.C.; Wolin, I.A.V.; Nascimento, A.P.M.; Amorim, R.M.F.; Rocha, B.A.M.; et al. Structural characterization of a lectin from Canavalia virosa seeds with inflammatory and cytotoxic activities. Int. J. Biol. Macromol. 2017, 94, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, J.G.; Bitencourt, F.S.; Mota, M.R.L.; Silvestre, P.P.; Aguiar, C.N.; Benevides, R.G.; Nascimento, K.S.; de Moura, T.R.; Dal-Secco, D.; Assreuy, A.M.S.; et al. Pharmacological analysis of the neutrophil migration induced by D. rostrata lectin: Involvement of cytokines and nitric oxide. Toxicon 2009, 54, 736–744. [Google Scholar] [CrossRef] [PubMed]
- Rangel, T.B.A.; Assreuy, A.M.S.; de Freitas Pires, A.; de Carvalho, A.U.; Benevides, R.G.; da Conceição Simões, R.; da Silva, H.C.; Bezerra, M.J.B.; do Nascimento, A.S.F.; do Nascimento, K.S.; et al. Crystallization and characterization of an inflammatory lectin purified from the seeds of Dioclea wilsonii. Molecules 2011, 16, 5087–5103. [Google Scholar] [CrossRef] [PubMed]
- Rangel, T.B.A.; Rocha, B.A.M.; Bezerra, G.A.; Assreuy, A.M.S.; de Freitas Pires, A.; do Nascimento, A.S.F.; Bezerra, M.J.B.; do Nascimento, K.S.; Nagano, C.S.; Sampaio, A.H.; et al. Crystal structure of a pro-inflammatory lectin from the seeds of Dioclea wilsonii Standl. Biochimie 2012, 94, 525–532. [Google Scholar] [CrossRef]
- Delatorre, P.; Rocha, B.A.M.; Simões, R.C.; Pereira-Júnior, F.N.; Silva, H.C.; Bezerra, E.H.S.; Bezerra, M.J.B.; Marinho, E.S.; Gadelha, C.A.A.; Santi-Gadelha, T.; et al. Mass spectrometry and X-ray diffraction analysis of two crystal types of Dioclea virgata lectin: An antinociceptive protein candidate to structure/function analysis. Appl. Biochem. Biotechnol. 2011, 164, 741–754. [Google Scholar] [CrossRef]
- Pinto-Junior, V.R.; Correia, J.L.A.; Pereira, R.I.; Pereira-Junior, F.N.; Santiago, M.Q.; Osterne, V.J.S.; Madeira, J.C.; Cajazeiras, J.B.; Nagano, C.S.; Delatorre, P.; et al. Purification and molecular characterization of a novel mannose-specific lectin from Dioclea reflexa hook seeds with inflammatory activity. J. Mol. Recognit. 2016, 29, 134–141. [Google Scholar] [CrossRef]
- Rocha, B.A.M.; Delatorre, P.; Oliveira, T.M.; Benevides, R.G.; Pires, A.F.; Sousa, A.A.S.; Souza, L.A.G.; Assreuy, A.M.S.; Debray, H.; de Azevedo, W.F., Jr.; et al. Structural basis for both pro- and anti-inflammatory response induced by mannose-specific legume lectin from Cymbosema roseum. Biochimie 2011, 93, 806–816. [Google Scholar] [CrossRef] [PubMed]
- Nunes, B.S.; Rensonnet, N.S.; Dal-Secco, D.; Vieira, S.M.; Cavada, B.S.; Teixeira, E.H.; Moura, T.R.; Teixeira, C.S.; Clemente-Napimoga, J.T.; Cunha, F.Q.; et al. Lectin extracted from Canavalia grandiflora seeds presents potential anti-inflammatory and analgesic effects. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2009, 379, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Pinto-Junior, V.R.; Osterne, V.J.S.; Santiago, M.Q.; Lossio, C.F.; Nagano, C.S.; Rocha, C.R.C.; Nascimento, J.C.F.; Nascimento, F.L.F.; Silva, I.B.; Oliveira, A.S.; et al. Molecular modeling, docking and dynamics simulations of the Dioclea lasiophylla Mart. Ex Benth seed lectin: An edematogenic and hypernociceptive protein. Biochimie 2017, 135, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Holanda, F.R.; Coelho-de-Sousa, A.N.; Assreuy, A.M.S.; Leal-Cardoso, J.H.; Pires, A.F.; do Nascimento, K.S.; Teixeira, C.S.; Cavada, B.S.; Santos, C.F. Antinociceptive activity of lectins from Diocleinae seeds on acetic acid-induced writhing test in mice. Protein Pept. Lett. 2009, 16, 1088–1092. [Google Scholar] [CrossRef] [PubMed]
- De Freitas Pires, A.; Assreuy, A.M.S.; Lopes, É.A.B.; Celedônio, N.R.; Soares, C.E.A.; Rodrigues, N.V.F.C.; Sousa, P.L.; Benevides, R.G.; Nagano, C.S.; Cavada, B.S.; et al. Opioid-like antinociceptive effects of oral administration of a lectin purified from the seeds of Canavalia brasiliensis. Fundam. Clin. Pharmacol. 2013, 27, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Kleha, J.F.; Devesly, P.; Johns, A. The effects of lectins on the release of EDRF from rabbit aorta. Br. J. Pharmacol. 1991, 104, 287–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Almeida Gadelha, C.A.; Moreno, F.B.M.B.; Santi-Gadelha, T.; Cajazeiras, J.B.; da Rocha, B.A.M.; Assreuy, A.M.S.; Lima Mota, M.R.; Pinto, N.V.; Passos Meireles, A.V.; Borges, J.C.; et al. Native crystal structure of a nitric oxide-releasing lectin from the seeds of Canavalia maritima. J. Struct. Biol. 2005, 152, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Bezerra, E.H.S.; Rocha, B.A.M.; Nagano, C.S.; de Arruda Bezerra, G.; de Moura, T.R.; Bezerra, M.J.B.; Benevides, R.G.; Sampaio, A.H.; Assreuy, A.M.S.; Delatorre, P.; et al. Structural analysis of ConBr reveals molecular correlation between the carbohydrate recognition domain and endothelial NO synthase activation. Biochem. Biophys. Res. Commun. 2011, 408, 566–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, B.A.M.; Barroso-Neto, I.L.; Teixeira, C.S.; Santiago, M.Q.; Pires, A.F.; Souza, L.A.G.; Nascimento, K.S.; Sampaio, A.H.; Delatorre, P.; Assreuy, A.M.S.; et al. CRLI induces vascular smooth muscle relaxation and suggests a dual mechanism of eNOS activation by legume lectins via muscarinic receptors and shear stress. Arch. Biochem. Biophys. 2015, 565, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Do Nascimento, A.S.F.; Gondim, A.C.S.; Cajazeiras, J.B.; Correia, J.L.A.; de Freitas Pires, A.; do Nascimento, K.S.; da Silva, A.L.C.; Nagano, C.S.; Assreuy, A.M.S.; Cavada, B.S. Purification and partial characterization of a novel lectin from Dioclea lasiocarpa Mart seeds with vasodilator effects. J. Mol. Recognit. 2012, 25, 657–664. [Google Scholar] [CrossRef]
- Pinto-Junior, V.R.; Osterne, V.J.S.; Santiago, M.Q.; Correia, J.L.A.; Pereira-Junior, F.N.; Leal, R.B.; Pereira, M.G.; Chicas, L.S.; Nagano, C.S.; Rocha, B.A.M.; et al. Structural studies of a vasorelaxant lectin from Dioclea reflexa Hook seeds: Crystal structure, molecular docking and dynamics. Int. J. Biol. Macromol. 2017, 98, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Barroso-Neto, I.L.; Delatorre, P.; Teixeira, C.S.; Correia, J.L.A.; Cajazeiras, J.B.; Pereira, R.I.; Nascimento, K.S.; Laranjeira, E.P.P.; Pires, A.F.; Assreuy, A.M.S.; et al. Structural analysis of a Dioclea sclerocarpa lectin: Study on the vasorelaxant properties of Dioclea lectins. Int. J. Biol. Macromol. 2016, 82, 464–470. [Google Scholar] [CrossRef] [PubMed]
- Bezerra, M.J.B.; Rodrigues, N.V.F.C.; de Freitas Pires, A.; Bezerra, G.A.; Nobre, C.B.; de Lima Alencar, K.L.; Soares, P.M.G.; do Nascimento, K.S.; Nagano, C.S.; Martins, J.L.; et al. Crystal structure of Dioclea violacea lectin and a comparative study of vasorelaxant properties with Dioclea rostrata lectin. Int. J. Biochem. Cell Biol. 2013, 45, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Fleming, I.; Busse, R. Molecular mechanisms involved in the regulation of the endothelial nitric oxide synthase. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2003, 284, R1–R12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Nóbrega, R.B.; Rocha, B.A.M.; Gadelha, C.A.A.; Santi-Gadelha, T.; Pires, A.F.; Assreuy, A.M.S.; Nascimento, K.S.; Nagano, C.S.; Sampaio, A.H.; Cavada, B.S.; et al. Structure of Dioclea virgata lectin: Relations between carbohydrate binding site and nitric oxide production. Biochimie 2012, 94, 900–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Mejía, E.G.; Prisecaru, V.I. Lectins as bioactive plant proteins: A potential in cancer treatment. Crit. Rev. Food Sci. Nutr. 2005, 45, 425–445. [Google Scholar] [CrossRef] [PubMed]
- Yau, T.; Dan, X.; Ng, C.C.W.; Ng, T.B. Lectins with potential for anti-cancer therapy. Molecules 2015, 20, 3791–3810. [Google Scholar] [CrossRef] [PubMed]
- Cavada, B.S.; Silva, M.T.L.; Osterne, V.J.S.; Pinto-Junior, V.R.; Nascimento, A.P.M.; Wolin, I.A.V.; Heinrich, I.A.; Nobre, C.A.S.; Moreira, C.G.; Lossio, C.F.; et al. Canavalia bonariensis lectin: Molecular bases of glycoconjugates interaction and antiglioma potential. Int. J. Biol. Macromol. 2018, 106, 369–378. [Google Scholar] [CrossRef]
- Nascimento, A.P.M.; Knaut, J.L.; Rieger, D.K.; Wolin, I.A.V.; Heinrich, I.A.; Mann, J.; Juarez, A.V.; Sosa, L.D.V.; de Paul, A.L.; Moreira, C.G.; et al. Anti-glioma properties of DVL, a lectin purified from Dioclea violacea. Int. J. Biol. Macromol. 2018, 120, 566–577. [Google Scholar] [CrossRef]
- Nascimento, K.S.; Santiago, M.Q.; Pinto-Junior, V.R.; Osterne, V.J.S.; Martins, F.W.V.; Nascimento, A.P.M.; Wolin, I.A.V.; Heinrich, I.A.; Martins, M.G.Q.; Silva, M.T.L.; et al. Structural analysis of Dioclea lasiocarpa lectin: A C6 cells apoptosis-inducing protein. Int. J. Biochem. Cell Biol. 2017, 92, 79–89. [Google Scholar] [CrossRef]
- Liu, B.; Li, C.-Y.; Bian, H.-J.; Min, M.-W.; Chen, L.-F.; Bao, J.-K. Antiproliferative activity and apoptosis-inducing mechanism of Concanavalin A on human melanoma A375 cells. Arch. Biochem. Biophys. 2009, 482, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Gondim, A.C.S.; Romero-Canelón, I.; Sousa, E.H.S.; Blindauer, C.A.; Butler, J.S.; Romero, M.J.; Sanchez-Cano, C.; Sousa, B.L.; Chaves, R.P.; Nagano, C.S.; et al. The potent anti-cancer activity of Dioclea lasiocarpa lectin. J. Inorg. Biochem. 2017, 175, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Cavalcante, T.T.A.; Anderson Matias da Rocha, B.; Alves Carneiro, V.; Vassiliepe Sousa Arruda, F.; Fernandes do Nascimento, A.S.; Cardoso Sá, N.; do Nascimento, K.S.; Sousa Cavada, B.; Holanda Teixeira, E. Effect of lectins from Diocleinae subtribe against oral Streptococci. Molecules 2011, 16, 3530–3543. [Google Scholar] [CrossRef] [PubMed]
- Gomes, B.S.; Siqueira, A.B.S.; de Cássia Carvalho Maia, R.; Giampaoli, V.; Teixeira, E.H.; Arruda, F.V.S.; do Nascimento, K.S.; de Lima, A.N.; Souza-Motta, C.M.; Cavada, B.S.; et al. Antifungal activity of lectins against yeast of vaginal secretion. Braz. J. Microbiol. 2012, 43, 770–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashraf, M.T.; Khan, R.H. Mitogenic lectins. Med. Sci. Monit. 2003, 9, RA265-9. [Google Scholar] [PubMed]
- De Oliveira Silva, F.; das Neves Santos, P.; de Melo, C.M.L.; Teixeira, E.H.; de Sousa Cavada, B.; Arruda, F.V.S.; Cajazeiras, J.B.; Almeida, A.C.; Pereira, V.A.R.; Porto, A.L.F. Immunostimulatory activity of ConBr: A focus on splenocyte proliferation and proliferative cytokine secretion. Cell Tissue Res. 2011, 346, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Tamma, S.M.L.; Kalyanaraman, V.S.; Pahwa, S.; Dominguez, P.; Modesto, R.R. The lectin jacalin induces phosphorylation of ERK and JNK in CD4+ T cells. J. Leukoc. Biol. 2003, 73, 682–688. [Google Scholar] [CrossRef]
- Beckert, W.H.; Sharkey, M.M. Mitogenic activity of the jack bean (Canavalia ensiformis) with rabbit peripheral blood lymphocytes. Int. Arch. Allergy Appl. Immunol. 1970, 39, 337–341. [Google Scholar] [CrossRef]
- De Melo, C.M.L.; Melo, H.; Correia, M.T.S.; Coelho, L.C.B.B.; da Silva, M.B.; Pereira, V.R.A. Mitogenic response and cytokine production induced by cramoll 1,4 lectin in splenocytes of inoculated mice. Scand. J. Immunol. 2011, 73, 112–121. [Google Scholar] [CrossRef]
- Souza, M.A.; Carvalho, F.C.; Ruas, L.P.; Ricci-Azevedo, R.; Roque-Barreira, M.C. The immunomodulatory effect of plant lectins: A review with emphasis on ArtinM properties. Glycoconj. J. 2013, 30, 641–657. [Google Scholar] [CrossRef]
- Da Silva, L.C.N.; Correia, M.T.S. Plant lectins and Toll-like receptors: Implications for therapy of microbial infections. Front. Microbiol. 2014, 5, 20. [Google Scholar] [CrossRef] [PubMed]
- Reis, E.A.G.; Athanazio, D.A.; Cavada, B.S.; Teixeira, E.H.; de Paulo Teixeira Pinto, V.; Carmo, T.M.A.; Reis, A.; Trocolli, G.; Croda, J.; Harn, D.; et al. Potential immunomodulatory effects of plant lectins in Schistosoma mansoni infection. Acta Trop. 2008, 108, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, C.R.; Cavassani, K.A.; Gomes, R.B.; Teixeira, M.J.; Roque-Barreira, M.-C.; Cavada, B.S.; da Silva, J.S.; Barral, A.; Barral-Netto, M. Potential of KM+ lectin in immunization against Leishmania amazonensis infection. Vaccine 2006, 24, 3001–3008. [Google Scholar] [CrossRef] [PubMed]
- Muraille, E.; Pajak, B.; Urbain, J.; Leo, O. Carbohydrate-bearing cell surface receptors involved in innate immunity: Interleukin-12 induction by mitogenic and nonmitogenic lectins. Cell. Immunol. 1999, 191, 1–9. [Google Scholar] [CrossRef] [PubMed]
- De Melo, C.M.L.; de Castro, M.C.A.B.; de Oliveira, A.P.; Gomes, F.O.S.; Pereira, V.R.A.; Correia, M.T.S.; Coelho, L.C.B.B.; Paiva, P.M.G. Immunomodulatory response of Cramoll 1,4 lectin on experimental lymphocytes. Phytother. Res. 2010, 24, 1631–1636. [Google Scholar] [CrossRef]
- Da Silva, L.C.N.; Alves, N.M.P.; de Castro, M.C.A.B.; Pereira, V.R.A.; da Paz, N.V.N.; Coelho, L.C.B.B.; de Figueiredo, R.C.B.Q.; Correia, M.T. Immunomodulatory effects of pCramoll and rCramoll on peritoneal exudate cells (PECs) infected and non-infected with Staphylococcus aureus. Int. J. Biol. Macromol. 2015, 72, 848–854. [Google Scholar] [CrossRef]
- De Oliveira, T.H.C.; Amorin, A.T.; Rezende, I.S.; Santos Barbosa, M.; Martins, H.B.; Brito, A.K.P.; Andrade, E.F.; Gonçalves, G.K.N.; Campos, G.B.; Silva, R.A.A.; et al. Sepsis induced by Staphylococcus aureus: Participation of biomarkers in a murine model. Med. Sci. Monit. 2015, 21, 345–355. [Google Scholar]
- Scherer, W.J.; Udin, S.B. Concanavalin A reduces habituation in the tectum of the frog. Brain Res. 1994, 667, 209–215. [Google Scholar] [CrossRef]
- Lin, S.S.; Levitan, I.B. Concanavalin A: A tool to investigate neuronal plasticity. Trends Neurosci. 1991, 14, 273–277. [Google Scholar] [CrossRef]
- Kirner, A.; Deutsch, S.; Weiler, E.; Polak, E.H.; Apfelbach, R. Concanavalin A application to the olfactory epithelium reveals different sensory neuron populations for the odour pair d- and l-carvone. Behav. Brain Res. 2003, 138, 201–206. [Google Scholar] [CrossRef]
- Suzuki, T.; Okumura-Noji, K. NMDA receptor subunits epsilon 1 (NR2A) and epsilon 2 (NR2B) are substrates for Fyn in the postsynaptic density fraction isolated from the rat brain. Biochem. Biophys. Res. Commun. 1995, 216, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.A.; Gurd, J.W.; Bissoon, N.; Tricaud, N.; Molnar, E.; Zamze, S.E.; Dwek, R.A.; McIlhinney, R.A.; Wing, D.R. Identification of lectin-purified neural glycoproteins, GPs 180, 116, and 110, with NMDA and AMPA receptor subunits: Conservation of glycosylation at the synapse. J. Neurochem. 1998, 70, 2594–2605. [Google Scholar] [CrossRef] [PubMed]
- Yankelevitch-Yahav, R.; Franko, M.; Huly, A.; Doron, R. The Forced Swim Test as a Model of Depressive-like Behavior. J. Vis. Exp. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barauna, S.C.; Kaster, M.P.; Heckert, B.T.; do Nascimento, K.S.; Rossi, F.M.; Teixeira, E.H.; Cavada, B.S.; Rodrigues, A.L.S.; Leal, R.B. Antidepressant-like effect of lectin from Canavalia brasiliensis (ConBr) administered centrally in mice. Pharmacol. Biochem. Behav. 2006, 85, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Rieger, D.K.; Costa, A.P.; Budni, J.; Moretti, M.; Barbosa, S.G.R.; Nascimento, K.S.; Teixeira, E.H.; Cavada, B.S.; Rodrigues, A.L.S.; Leal, R.B. Antidepressant-like effect of Canavalia brasiliensis (ConBr) lectin in mice: Evidence for the involvement of the glutamatergic system. Pharmacol. Biochem. Behav. 2014, 122, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Russi, M.A.; Vandresen-Filho, S.; Rieger, D.K.; Costa, A.P.; Lopes, M.W.; Cunha, R.M.S.; Teixeira, E.H.; Nascimento, K.S.; Cavada, B.S.; Tasca, C.I.; et al. ConBr, a lectin from Canavalia brasiliensis seeds, protects against quinolinic acid-induced seizures in mice. Neurochem. Res. 2012, 37, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Macedo, M.L.R.; Oliveira, C.F.R.; Oliveira, C.T. Insecticidal activity of plant lectins and potential application in crop protection. Molecules 2015, 20, 2014–2033. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Montaño, E.A.; Vega-Castro, N.A. Plant Lectins with Insecticidal and Insectistatic Activities. In Insecticides—Agriculture and Toxicology; IntechOpen: London, UK, 2018. [Google Scholar] [Green Version]
- Sauvion, N.; Nardon, C.; Febvay, G.; Gatehouse, A.M.R.; Rahbé, Y. Binding of the insecticidal lectin Concanavalin A in pea aphid, Acyrthosiphon pisum (Harris) and induced effects on the structure of midgut epithelial cells. J. Insect Physiol. 2004, 50, 1137–1150. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.T.; Kunz, D.; Silva, C.P.; Macedo, M.L.R. Entomotoxic properties of Dioclea violacea lectin and its effects on digestive enzymes of Anagasta kuehniella (Lepidoptera). J. Insect Physiol. 2015, 81, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Rahbé, Y.; Sauvion, N.; Febvay, G.; Peumans, W.J.; Gatehouse, A.M.R. Toxicity of lectins and processing of ingested proteins in the pea aphid Acyrthosiphon pisum. Entomol. Exp. Appl. 1995, 76, 143–155. [Google Scholar] [CrossRef]
- Grangeiro, T.B. Clonagem, Sequenciamento e Expressão do Gene da Lectina (ConBr) de Sementes de Canavalia brasiliensis. Ph.D. Thesis, Federal University of Ceara, Fortaleza, Brazil, 1996. [Google Scholar]
- Cunningham, B.A.; Wang, J.L.; Waxdal, M.J.; Edelman, G.M. The covalent and three-dimensional structure of concanavalin A. II. Amino acid sequence of cyanogen bromide fragment F3. J. Biol. Chem. 1975, 250, 1503–1512. [Google Scholar]
- Perez, G.; Perez, C.; Sousa-Cavada, B.; Moreira, R.; Richardson, M. Comparison of the amino acid sequences of the lectins from seeds of Dioclea lehmanni and Canavalia maritima. Phytochemistry 1991, 30, 2619–2621. [Google Scholar] [CrossRef]
- Grangeiro, T.B.; Schriefer, A.; Calvete, J.J.; Raida, M.; Urbanke, C.; Barral-Netto, M.; Cavada, B.S. Molecular cloning and characterization of ConBr, the lectin of Canavalia brasiliensis seeds. Eur. J. Biochem. 1997, 248, 43–48. [Google Scholar] [CrossRef]
- Bezerra, G.A.; Viertlmayr, R.; Moura, T.R.; Delatorre, P.; Rocha, B.A.M.; do Nascimento, K.S.; Figueiredo, J.G.; Bezerra, I.G.; Teixeira, C.S.; Simões, R.C.; et al. Structural studies of an anti-inflammatory lectin from Canavalia boliviana seeds in complex with dimannosides. PLoS ONE 2014, 9, e97015. [Google Scholar] [CrossRef]
- Silva, M.; Silva, S.; Nascimento, K.; Nagano, C.; Cavada, B. Physico-chemical characterization and partial sequence of a lectin from Canavalia bonariensis Lindl seeds. BMC Proc. 2014, 8, P227. [Google Scholar] [CrossRef]
- Barroso-Neto, I.L.; Rocha, B.A.M.; Simoes, R.C.; Bezerra, M.J.B.; Pereira-Junior, F.N.; Osterne, V.J.S.; Nascimento, K.S.; Nagano, C.S.; Delatorre, P.; Sampaio, A.H.; et al. Crystal Structure of Canavalia grandiflora Seed Lectin Complexed with X-Man. 2014. Available online: https://www.rcsb.org/structure/4L8Q (accessed on 20 December 2018). [CrossRef]
- De Oliveira, T.M.; Delatorre, P.; da Rocha, B.A.M.; de Souza, E.P.; Nascimento, K.S.; Bezerra, G.A.; Moura, T.R.; Benevides, R.G.; Bezerra, E.H.S.; Moreno, F.B.M.B.; et al. Crystal structure of Dioclea rostrata lectin: Insights into understanding the pH-dependent dimer-tetramer equilibrium and the structural basis for carbohydrate recognition in Diocleinae lectins. J. Struct. Biol. 2008, 164, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Calvete, J.J.; Thole, H.H.; Raida, M.; Urbanke, C.; Romero, A.; Grangeiro, T.B.; Ramos, M.V.; Almeida da Rocha, I.M.; Guimarães, F.N.; Cavada, B.S. Molecular characterization and crystallization of Diocleinae lectins. Biochim. Biophys. Acta 1999, 1430, 367–375. [Google Scholar] [CrossRef]
- De Souza, G.A.; Oliveira, P.S.L.; Trapani, S.; Santos, A.C.O.; Rosa, J.C.; Laure, H.J.; Faça, V.M.; Correia, M.T.S.; Tavares, G.A.; Oliva, G.; et al. Amino acid sequence and tertiary structure of Cratylia mollis seed lectin. Glycobiology 2003, 13, 961–972. [Google Scholar] [CrossRef]
- Souza Teixeira, C.; da Silva, H.C.; de Moura, T.R.; Pereira-Junior, F.N.; do Nascimento, K.S.; Shiniti Nagano, C.; Holanda Sampaio, A.; Delatorre, P.; Matias Rocha, B.A.; Sousa Cavada, B. Crystal structure of the lectin of Camptosema pedicellatum: Implications of a conservative substitution at the hydrophobic subsite. J. Biochem. 2012, 152, 87–98. [Google Scholar] [CrossRef]
- Correia, J.L.A.; do Nascimento, A.S.F.; Cajazeiras, J.B.; Gondim, A.C.S.; Pereira, R.I.; de Sousa, B.L.; da Silva, A.L.C.; Garcia, W.; Teixeira, E.H.; do Nascimento, K.S.; et al. Molecular characterization and tandem mass spectrometry of the lectin extracted from the seeds of Dioclea sclerocarpa Ducke. Molecules 2011, 16, 9077–9089. [Google Scholar] [CrossRef]
- Herman, E.M.; Shannon, L.M.; Chrispeels, M.J. Concanavalin A is synthesized as a glycoprotein precursor. Planta 1985, 165, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Chrispeels, M.J.; Hartl, P.M.; Sturm, A.; Faye, L. Characterization of the endoplasmic reticulum-associated precursor of concanavalin A. Partial amino acid sequence and lectin activity. J. Biol. Chem. 1986, 261, 10021–10024. [Google Scholar] [PubMed]
- Bowles, D.J.; Marcus, S.E.; Pappin, D.J.; Findlay, J.B.; Eliopoulos, E.; Maycox, P.R.; Burgess, J. Posttranslational processing of concanavalin A precursors in jackbean cotyledons. J. Cell Biol. 1986, 102, 1284–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faye, L.; Chrispeels, M.J. Transport and processing of the glycosylated precursor of Concanavalin A in jack-bean. Planta 1987, 170, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Bowles, D.J.; Pappin, D.J. Traffic and assembly of concanavalin A. Trends Biochem. Sci. 1988, 13, 60–64. [Google Scholar] [CrossRef]
- Hardman, K.D.; Ainsworth, C.F. Structure of concanavalin A at 2.4-A resolution. Biochemistry 1972, 11, 4910–4919. [Google Scholar] [CrossRef] [PubMed]
- Argos, P.; Tsukihara, T.; Rossmann, M.G. A structural comparison of concanavalin A and tomato bushy stunt virus protein. J. Mol. Evol. 1980, 15, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Chelvanayagam, G.; Heringa, J.; Argos, P. Anatomy and evolution of proteins displaying the viral capsid jellyroll topology. J. Mol. Biol. 1992, 228, 220–242. [Google Scholar] [CrossRef]
- Loris, R.; Hamelryck, T.; Bouckaert, J.; Wyns, L. Legume lectin structure. Biochim. Biophys. Acta 1998, 1383, 9–36. [Google Scholar] [CrossRef]
- Silva, M.T.L.; Osterne, V.J.S.; Nobre, C.A.S.; Chaves, R.P.; Silva, I.B.; Moreira, C.G.; Andrade, M.L.L.; Nagano, C.S.; Rocha, C.R.C.; Leal, R.B.; et al. Structural characterization, docking and dynamics simulations of Canavalia bonariensis lectin. J. Drug Des. Res. 2016, 3, 1–8. [Google Scholar]
- Bezerra, G.A.; Oliveira, T.M.; Moreno, F.B.M.B.; de Souza, E.P.; da Rocha, B.A.M.; Benevides, R.G.; Delatorre, P.; de Azevedo, W.F., Jr.; Cavada, B.S. Structural analysis of Canavalia maritima and Canavalia gladiata lectins complexed with different dimannosides: New insights into the understanding of the structure-biological activity relationship in legume lectins. J. Struct. Biol. 2007, 160, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Sinha, S.; Gupta, G.; Vijayan, M.; Surolia, A. Subunit assembly of plant lectins. Curr. Opin. Struct. Biol. 2007, 17, 498–505. [Google Scholar] [CrossRef] [PubMed]
- Brinda, K.V.; Mitra, N.; Surolia, A.; Vishveshwara, S. Determinants of quaternary association in legume lectins. Protein Sci. 2004, 13, 1735–1749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamp, R.M.; Calvete, J.J.; Choli-Papadopoulou, T. Methods in Proteome and Protein Analysis; Springer Science & Business Media: Berlin, Germany, 2013; ISBN 9783662087220. [Google Scholar]
- Agrawal, B.B.; Goldstein, I.J. Protein-carbohydrate interaction. VII. Physical and chemical studies on concanavalin A, the hemagglutinin of the jack bean. Arch. Biochem. Biophys. 1968, 124, 218–229. [Google Scholar] [CrossRef]
- Kalb, A.J.; Joseph Kalb, A.; Lustig, A. The molecular weight of concanavalin A. Biochim. Biophys. Acta (BBA) Protein Struct. 1968, 168, 366–367. [Google Scholar] [CrossRef]
- Wah, D.A.; Romero, A.; del Sol, F.G.; Cavada, B.S.; Ramos, M.V.; Grangeiro, T.B.; Sampaio, A.H.; Calvete, J.J. Crystal structure of native and Cd/Cd-substituted Dioclea guianensis seed lectin. A novel manganese-binding site and structural basis of dimer-tetramer association. J. Mol. Biol. 2001, 310, 885–894. [Google Scholar] [CrossRef]
- Senear, D.F.; Teller, D.C. Thermodynamics of concanavalin A dimer-tetramer self-association: Sedimentation equilibrium studies. Biochemistry 1981, 20, 3076–3083. [Google Scholar] [CrossRef]
- Del Sol, F.G.; Cavada, B.S.; Calvete, J.J. Crystal structures of Cratylia floribunda seed lectin at acidic and basic pHs. Insights into the structural basis of the pH-dependent dimer–tetramer transition. J. Struct. Biol. 2007, 158, 1–9. [Google Scholar] [CrossRef]
- Zamora-Caballero, S.; Pérez, A.; Sanz, L.; Bravo, J.; Calvete, J.J. Quaternary structure of Dioclea grandiflora lectin assessed by equilibrium sedimentation and crystallographic analysis of recombinant mutants. FEBS Lett. 2015, 589, 2290–2296. [Google Scholar] [CrossRef]
- Suseelan, K.N.; Bhagwath, A.; Pandey, R.; Gopalakrishna, T. Characterization of Con C, a lectin from Canavalia cathartica Thouars seeds. Food Chem. 2007, 104, 528–535. [Google Scholar] [CrossRef]
- Leal, R.B.; Pinto-Junior, V.R.; Osterne, V.J.S.; Wolin, I.A.V.; Nascimento, A.P.M.; Neco, A.H.B.; Araripe, D.A.; Welter, P.G.; Neto, C.C.; Correia, J.L.A.; et al. Crystal structure of DlyL, a mannose-specific lectin from Dioclea lasiophylla Mart. Ex Benth seeds that display cytotoxic effects against C6 glioma cells. Int. J. Biol. Macromol. 2018, 114, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Ghazarian, H.; Idoni, B.; Oppenheimer, S.B. A glycobiology review: Carbohydrates, lectins and implications in cancer therapeutics. Acta Histochem. 2011, 113, 236–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Q.; Zhao, L.; Song, Q.; Wang, Z.; Qiu, X.; Zhang, W.; Zhao, M.; Zhao, G.; Liu, W.; Liu, H.; et al. Hybrid- and complex-type N-glycans are not essential for Newcastle disease virus infection and fusion of host cells. Glycobiology 2011, 22, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Nagae, M.; Yamaguchi, Y. Function and 3D structure of the N-glycans on glycoproteins. Int. J. Mol. Sci. 2012, 13, 8398–8429. [Google Scholar] [CrossRef] [PubMed]
- Bieberich, E. Synthesis, Processing, and Function of N-glycans in N-glycoproteins. In Advances in Neurobiology; Springer: New York, NY, USA, 2014; pp. 47–70. [Google Scholar]
- Pinho, S.S.; Reis, C.A. Glycosylation in cancer: Mechanisms and clinical implications. Nat. Rev. Cancer 2015, 15, 540–555. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Swoger, J.; Greene, S.; Saito, T.; Hurh, J.; Sweeley, C.; Leestma, J.; Mkrdichian, E.; Cerullo, L.; Nishikawa, A.; et al. Beta1,6-N-acetylglucosamine-bearing N-glycans in human gliomas: Implications for a role in regulating invasivity. Cancer Res. 2000, 60, 134–142. [Google Scholar]
- Stanley, P.; Taniguchi, N.; Aebi, M. N-Glycans. In Essentials of Glycobiology; Varki, A., Cummings, R.D., Esko, J.D., Stanley, P., Hart, G.W., Aebi, M., Darvill, A.G., Kinoshita, T., Packer, N.H., Prestegard, J.H., et al., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2017. [Google Scholar]
- Stowell, S.R.; Ju, T.; Cummings, R.D. Protein glycosylation in cancer. Annu. Rev. Pathol. 2015, 10, 473–510. [Google Scholar] [CrossRef]
- Sirois, S.; Touaibia, M.; Chou, K.-C.; Roy, R. Glycosylation of HIV-1 gp120 V3 loop: Towards the rational design of a synthetic carbohydrate vaccine. Curr. Med. Chem. 2007, 14, 3232–3242. [Google Scholar] [CrossRef]
- Qi, Y.; Jo, S.; Im, W. Roles of glycans in interactions between gp120 and HIV broadly neutralizing antibodies. Glycobiology 2016, 26, 251–260. [Google Scholar] [CrossRef]
- Rudd, P.M. Glycosylation and the Immune System. Science 2001, 291, 2370–2376. [Google Scholar] [CrossRef]
- Schwarz, F.; Aebi, M. Mechanisms and principles of N-linked protein glycosylation. Curr. Opin. Struct. Biol. 2011, 21, 576–582. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Isaji, T.; Xu, Q.; Kariya, Y.; Gu, W.; Fukuda, T.; Du, Y. Potential roles of N-glycosylation in cell adhesion. Glycoconj. J. 2012, 29, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Mandal, D.K.; Brewer, C.F. Differences in the binding affinities of dimeric concanavalin A (including acetyl and succinyl derivatives) and tetrameric concanavalin A with large oligomannose-type glycopeptides. Biochemistry 1993, 32, 5116–5120. [Google Scholar] [CrossRef] [PubMed]
- Dam, T.K.; Roy, R.; Das, S.K.; Oscarson, S.; Brewer, C.F. Binding of multivalent carbohydrates to Concanavalin A and Dioclea grandiflora lectin: Thermodynamic analysis of the “multivalency effect”. J. Biol. Chem. 2000, 275, 14223–14230. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lectin | Initial | Edema Peak | Edema (mL) | Duration | Indomethacin | L-NAME a | Carbohydrate b |
|---|---|---|---|---|---|---|---|
| ConA | 1 h | 1 h | 0.29 | 24 h | NT c | NT | Partially |
| ConBr | 30 min | 30 min | 0.92 | 48 h | Partially | Partially | Partially |
| CGL | 30 min | 30 min | 0.68 | 32 h | Partially | Partially | Partially |
| ConM | 30 min | 30 min | 0.56 | 5 h | Partially | No | Blocked |
| ConGF | 30 min | 30 min | 0.41 | 24 h | NT | NT | NT |
| CoxyL | 30 min | 2 h | 1.10 | 5 h | NT | NT | NT |
| DWL | 30 min | 5 h | 0.80 | 24 h | Partially | Partially | No |
| DlyL | 30 min | 1 h | 1.06 | 24 h | Partially | Partially | No |
| CRL1 | 3 h | 4h | 0.47 | 24 h | NT | NT | Partially |
| Lectin | Endothelium-Dependent | Relaxation | Indomethacin | L-NAME a | Carbohydrate b |
|---|---|---|---|---|---|
| ConBr | Yes | 74% | Partially | Partially | Partially |
| CGL | Yes | 108% | Partially | Blocked | Partially |
| ConGF | Yes | 25% | NT c | Blocked | NT |
| ConM | Yes | 110% | NT | Blocked | NT |
| ConA | Yes | 85% | NT | NT | NT |
| ConV | Yes | 77% | NT | NT | Partially |
| CRL1 | Yes | 96% | No | Blocked | NT |
| DLL | Yes | 81% | NT | Partially | Partially |
| DrfL | Yes | 32% | NT | Blocked | Partially |
| DSL | Yes | 36% | NT | Blocked | Blocked |
| DRL | Yes | 96% | Partially | Blocked | Partially |
| DVL | Yes | 43% | No | Partially | Partially |
| DvirL | Yes | 70% | NT | Blocked | NT |
| Lectin | CC 50 (µg/mL) | Confidence Interval |
|---|---|---|
| ConA | 56.02 | 38.01–80.03 |
| CaBo | 230.50 | 59.84–88.8 |
| ConV | 58.8 | 52.45–140.3 |
| DVL | 58.84 | 51.14–67.70 |
| DLL | 70.51 | 58.32–85.25 |
| Lectin | PDB ID | Uniprot ID |
|---|---|---|
| ConA | 1JBC | P02866 |
| ConV | 5F5Q | C0HJY1 |
| CGL | 1WUV | P14894 |
| ConM | 2CWM | P81364 |
| ConBr | 1AZD | P55915 |
| CboL | 4K20 | A0A023GPI8 |
| CaBo | 5U3E | P58906 |
| ConGF | 4L8Q | A0A067XG71 |
| DvirL | 3RRD | P58907 |
| CRL | 3A0K | P86184 |
| DLL | 5UUY | - |
| DlyL | 6CJ9 | C0HK27 |
| DRL | 2ZBJ | P58908 |
| DguiL | 1H9W | P81637 |
| DGL | 1DGL | P08902 |
| Cramoll | 1MVQ | P83721 |
| CFL | 2D3P | P81517 |
| DVL | 2GDF | I1SB09 |
| CPL | 3U4X | J9PBR3 |
| DSL | 4NOT | B3EWJ2 |
| DWL | 3SH3 | P86624 |
| DrfL | 5TG3 | C0HK81 |
| DlehL | - | - |
| Lectin | Cα RMSD |
|---|---|
| ConV | 0.260 |
| CGL | 0.346 |
| ConM | 0.304 |
| ConBr | 0.310 |
| CboL | 0.299 |
| CaBo | 0.335 |
| ConGF | 0.467 |
| DvirL | 0.322 |
| CRL | 0.467 |
| DLL | 0.290 |
| DlyL | 0.283 |
| DRL | 0.287 |
| DguiL | 0.276 |
| DGL | 0.368 |
| Cramoll | 0.331 |
| CFL | 0.330 |
| DVL | 0.369 |
| CPL | 0.270 |
| DSL | 0.280 |
| DWL | 0.351 |
| DrfL | 0.312 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cavada, B.S.; Pinto-Junior, V.R.; Osterne, V.J.S.; Nascimento, K.S. ConA-Like Lectins: High Similarity Proteins as Models to Study Structure/Biological Activities Relationships. Int. J. Mol. Sci. 2019, 20, 30. https://doi.org/10.3390/ijms20010030
Cavada BS, Pinto-Junior VR, Osterne VJS, Nascimento KS. ConA-Like Lectins: High Similarity Proteins as Models to Study Structure/Biological Activities Relationships. International Journal of Molecular Sciences. 2019; 20(1):30. https://doi.org/10.3390/ijms20010030
Chicago/Turabian StyleCavada, Benildo S., Vanir R. Pinto-Junior, Vinicius J. S. Osterne, and Kyria S. Nascimento. 2019. "ConA-Like Lectins: High Similarity Proteins as Models to Study Structure/Biological Activities Relationships" International Journal of Molecular Sciences 20, no. 1: 30. https://doi.org/10.3390/ijms20010030
APA StyleCavada, B. S., Pinto-Junior, V. R., Osterne, V. J. S., & Nascimento, K. S. (2019). ConA-Like Lectins: High Similarity Proteins as Models to Study Structure/Biological Activities Relationships. International Journal of Molecular Sciences, 20(1), 30. https://doi.org/10.3390/ijms20010030