Molecular Advances on Phytases in Barley and Wheat

Abstract
:1. Introduction
2. Plant and Microbial Phytases
3. Mature Grain Phytase Activity
4. Classes of Phytases in Barley and Wheat
5. Biochemical Properties and Storage of the PAPhys
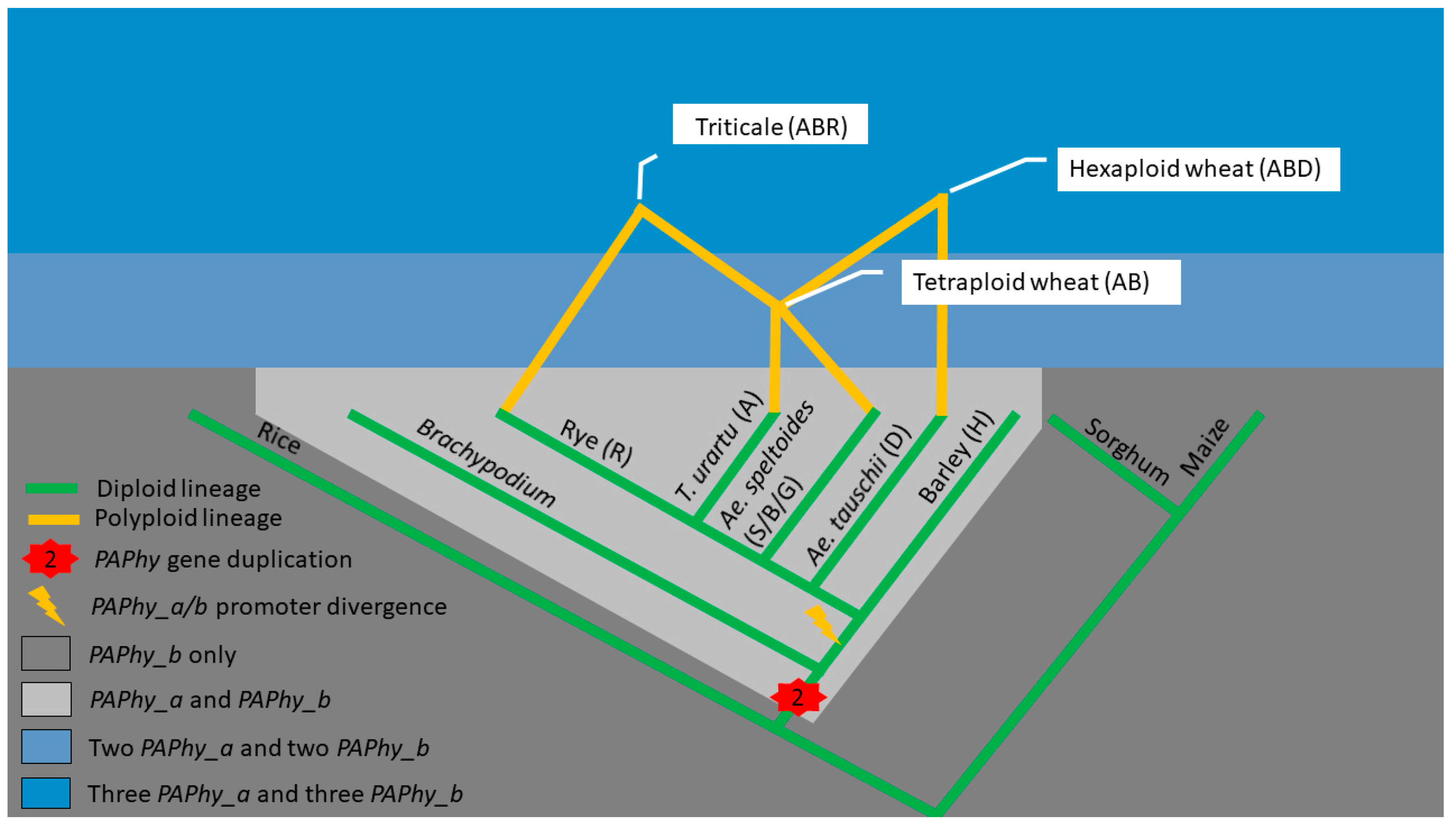
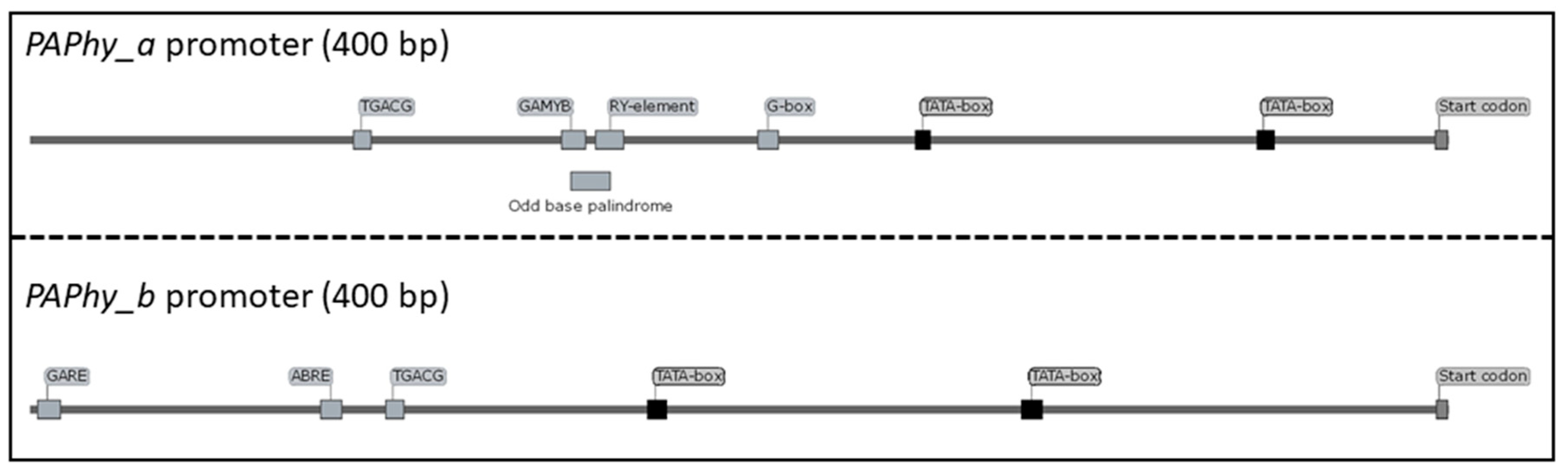
6. PAPhy Genetics
7. Applied Potentials of the PAPhys
8. Achieving Higher MGPA with the PAPhys
Funding
Conflicts of Interest
References
- Brinch-Pedersen, H.; Madsen, C.K.; Holme, I.B.; Dionisio, G. Increased understanding of the cereal phytase complement for better mineral bio-availability and resource management. J. Cereal Sci. 2014, 59, 373–381. [Google Scholar] [CrossRef]
- Bethke, P. From Storage Compartment to Lytic Organelle: The Metamorphosis of the Aleurone Protein Storage Vacuole. Ann. Bot. 1998, 82, 399–412. [Google Scholar] [CrossRef] [Green Version]
- Bohn, L.; Josefsen, L.; Meyer, A.S.; Rasmussen, S.K. Quantitative Analysis of Phytate Globoids Isolated from Wheat Bran and Characterization of Their Sequential Dephosphorylation by Wheat Phytase. J. Agric. Food Chem. 2007, 55, 7547–7552. [Google Scholar] [CrossRef] [PubMed]
- Eeckhout, W.; De Paepe, M. Total phosphorus, phytate-phosphorus and phytase activity in plant feedstuffs. Anim. Feed Sci. Technol. 1994, 47, 19–29. [Google Scholar] [CrossRef]
- Steiner, T.; Mosenthin, R.; Zimmermann, B.; Greiner, R.; Roth, S. Distribution of phytase activity, total phosphorus and phytate phosphorus in legume seeds, cereals and cereal by-products as influenced by harvest year and cultivar. Anim. Feed Sci. Technol. 2007, 133, 320–334. [Google Scholar] [CrossRef]
- Viveros, A.; Centeno, C.; Brenes, A.; Canales, R.; Lozano, A. Phytase and Acid Phosphatase Activities in Plant Feedstuffs. J. Agric. Food Chem. 2000, 48, 4009–4013. [Google Scholar] [CrossRef] [Green Version]
- Bouis, H.E. Special Issue on Improving Human Nutrition Through Agriculture. Food Nutr. Bull. 2000, 21, 351–578. [Google Scholar]
- De Benoist, B.; Cogswell, M.; Egli, I.; McLean, E. Worldwide prevalence of anaemia 1993–2005. In WHO Global Database of Anaemia; WHO: Geneva, Switzerland, 2008. [Google Scholar]
- Stoltzfus, R.J.; Mullany, L.; Black, R.E. Iron deficiency anemia. In Comparative Quantification of Health Risks: Global and Regional Burden of Disease Attribution to Selected Major Risk Factors; Ezzati, M., Lopez, A.D., Rodgers, A., Murray, C.J., Eds.; World Health Organization: Geneva, Switzerland, 2004; ISBN 9241580313. [Google Scholar]
- Brown, K.H.; Rivera, J.A.; Bhutta, Z.A.; Gibson, R.S.; King, J.C.; Lonnerdal, B. Chapter 2: Assessment of the Risk of Zinc Deficiency in Populations. Food Nutr. Bull. 2004, 25, S130. [Google Scholar]
- Yakoob, M.Y.; Theodoratou, E.; Jabeen, A.; Imdad, A.; Eisele, T.P.; Ferguson, J.; Jhass, A.; Rudan, I.; Campbell, H.; Black, R.E.; et al. Preventive zinc supplementation in developing countries: impact on mortality and morbidity due to diarrhea, pneumonia and malaria. BMC Public Health 2011, 11, S23. [Google Scholar] [CrossRef] [PubMed]
- Schindler, D.W.; Carpenter, S.R.; Chapra, S.C.; Hecky, R.E.; Orihel, D.M. Reducing Phosphorus to Curb Lake Eutrophication is a Success. Environ. Sci. Technol. 2016, 50, 8923–8929. [Google Scholar] [CrossRef] [PubMed]
- Cordell, D.; Drangert, J.-O.; White, S. The story of phosphorus: Global food security and food for thought. Glob. Environ. Chang. 2009, 19, 292–305. [Google Scholar] [CrossRef]
- Lei, X.G.; Porres, J.M.; Mullaney, E.J.; Brinch-Pedersen, H. Phytase: Source, Structure and Application. In Industrial Enzymes: Structure, Function and Applications; Polaina, J., MacCabe, A., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 505–530. ISBN 978-1-4020-5376-4. [Google Scholar]
- Dionisio, G.; Holm, P.B.; Brinch-Pedersen, H. Wheat (Triticum aestivum L.) and barley (Hordeum vulgare L.) multiple inositol polyphosphate phosphatases (MINPPs) are phytases expressed during grain filling and germination. Plant Biotechnol. J. 2007, 5, 325–338. [Google Scholar] [CrossRef]
- Dionisio, G.; Madsen, C.K.; Holm, P.B.; Welinder, K.G.; Jørgensen, M.; Stoger, E.; Arcalis, E.; Brinch-Pedersen, H. Cloning and characterization of purple acid phosphatase phytases from wheat, barley, maize, and rice. Plant Physiol. 2011, 156. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.H.J.; Phillippy, B.Q. Substrate selectivity in Aspergillus ficuum phytase and acid phosphatases using myo-inositol phosphates. J. Agric. Food Chem. 1994, 42, 423–425. [Google Scholar] [CrossRef]
- Holme, I.B.; Wendt, T.; Gil-Humanes, J.; Deleuran, L.C.; Starker, C.G.; Voytas, D.F.; Brinch-Pedersen, H. Evaluation of the mature grain phytase candidate HvPAPhy_a gene in barley (Hordeum vulgare L.) using CRISPR/Cas9 and TALENs. Plant Mol. Biol. 2017, 95, 111–121. [Google Scholar] [CrossRef]
- Swanson, S.J.; Jones, R.L. Gibberellic Acid Induces Vacuolar Acidification in Barley Aleurone. Plant Cell 1996, 8, 2211, LP-2221. [Google Scholar] [CrossRef] [PubMed]
- Greiner, R.; Jany, K.-D.; Larsson Alminger, M. Identification and Properties of myo -Inositol Hexakisphosphate Phosphohydrolases (Phytases) from Barley (Hordeum vulgare). J. Cereal Sci. 2000, 31, 127–139. [Google Scholar] [CrossRef]
- Jiang, L.; Phillips, T.E.; Hamm, C.A.; Drozdowicz, Y.M.; Rea, P.A.; Maeshima, M.; Rogers, S.W.; Rogers, J.C. The protein storage vacuole. J. Cell Biol. 2001, 155, 991, LP-1002. [Google Scholar] [CrossRef]
- Madsen, C.K.; Dionisio, G.; Holme, I.B.; Holm, P.B.; Brinch-Pedersen, H. High mature grain phytase activity in the Triticeae has evolved by duplication followed by neofunctionalization of the purple acid phosphatase phytase (PAPhy) gene. J. Exp. Bot. 2013, 64. [Google Scholar] [CrossRef]
- IWGSC A chromosome-based draft sequence of the hexaploid bread wheat (Triticum aestivum) genome. Science 2014, 345, 1251788. [CrossRef]
- IWGSC IWGSC-Blast. Available online: https://urgi.versailles.inra.fr/blast_iwgsc/blast.php (accessed on 17 May 2019).
- Madsen, C.K.; Petersen, G.; Seberg, O.; Brinch-Pedersen, H. Evolution and diversity of PAPhy_a phytase in the genepool of wheat (Triticum aestivum L., Poaceae). Genet. Resour. Crop Evol. 2017, 64. [Google Scholar] [CrossRef]
- Guo, W.; Yang, H.; Liu, Y.; Gao, Y.; Ni, Z.; Peng, H.; Xin, M.; Hu, Z.; Sun, Q.; Yao, Y. The wheat transcription factor TaGAMyb recruits histone acetyltransferase and activates the expression of a high-molecular-weight glutenin subunit gene. Plant J. 2015, 84, 347–359. [Google Scholar] [CrossRef]
- De Pater, S.; Katagiri, F.; Kijne, J.; Chua, N.-H. bZIP proteins bind to a palindromic sequence without an ACGT core located in a seed-specific element of the pea lectin promoter. Plant J. 1994, 6, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.; Knudsen, S. The nitrogen response of a barley C-hordein promoter is controlled by positive and negative regulation of the GCN4 and endosperm box. Plant J. 1993, 4, 343–355. [Google Scholar] [CrossRef]
- Bäumlein, H.; Nagy†, I.; Villarroel, R.; Inzé, D.; Wobus, U. Cis-analysis of a seed protein gene promoter: the conservative RY repeat CATGCATG within the legumin box is essential for tissue-specific expression of a legumin gene. Plant J. 1992, 2, 233–239. [Google Scholar]
- Holme, I.B.; Dionisio, G.; Brinch-Pedersen, H.; Wendt, T.; Madsen, C.K.; Vincze, E.; Holm, P.B. Cisgenic barley with improved phytase activity. Plant Biotechnol. J. 2012, 10. [Google Scholar] [CrossRef]
- Holme, I.B.; Dionisio, G.; Madsen, C.K.; Brinch-Pedersen, H. Barley HvPAPhy_a as transgene provides high and stable phytase activities in mature barley straw and in grains. Plant Biotechnol. J. 2017, 15. [Google Scholar] [CrossRef]
- Pointillart, A.; Fourdin, A.; Fontaine, N. Importance of Cereal Phytase Activity for Phytate Phosphorus Utilization by Growing Pigs Fed Diets Containing Triticale or Corn. J. Nutr. 1987, 117, 907–913. [Google Scholar] [CrossRef]
- Sandberg, A.-S.; Andersson, H. Effect of Dietary Phytase on the Digestion of Phytate in the Stomach and Small Intestine of Humans. J. Nutr. 1988, 118, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Türk, M.; Carlsson, N.-G.; Sandberg, A.-S. Reduction in the Levels of Phytate During Wholemeal Bread Making; Effect of Yeast and Wheat Phytases. J. Cereal Sci. 1996, 23, 257–264. [Google Scholar] [CrossRef]
- Koréissi-Dembélé, Y.; Fanou-Fogny, N.; Moretti, D.; Schuth, S.; Dossa, R.A.M.; Egli, I.; Zimmermann, M.B.; Brouwer, I.D. Dephytinisation with intrinsic wheat phytase and iron fortification significantly increase iron absorption from fonio (Digitaria exilis) meals in West African women. PLoS ONE 2013, 8, e70613. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Wu, S.; Tiwari, V.; Sehgal, S.; Raupp, J.; Wilson, D.; Abbasov, M.; Gill, B.; Poland, J. Genomic Analysis Confirms Population Structure and Identifies Inter-Lineage Hybrids in Aegilops tauschii. Front. Plant Sci. 2019, 10, 9. [Google Scholar] [CrossRef]
- Feuillet, C.; Langridge, P.; Waugh, R. Cereal breeding takes a walk on the wild side. Trends Genet. 2008, 24, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Brinch-Pedersen, H.; Madsen, C.K.; Dionisio, G.; Holm, P.B. High expression cereal phytase gene. WO 2012/146597 Al. 2012. Available online: https://patents.google.com/patent/WO2012146597A1/en (accessed on 17 May 2019).
- Scholey, D.; Burton, E.; Morgan, N.; Sanni, C.; Madsen, C.K.; Dionisio, G.; Brinch-Pedersen, H. P and Ca digestibility is increased in broiler diets supplemented with the high-phytase HIGHPHY wheat. Animal 2017, 11. [Google Scholar] [CrossRef]
- Yin, K.; Gao, C.; Qiu, J.-L. Progress and prospects in plant genome editing. Nat. Plants 2017, 3, 17107. [Google Scholar] [CrossRef] [PubMed]
- Bekalu, Z.E.; Madsen, C.K.; Dionisio, G.; Brinch-Pedersen, H. Aspergillus ficuum phytase activity is inhibited by cereal grain components. PLoS ONE 2017, 12. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Cereal | n | Total P (% of Dry Matter) | Total IP6 P (% of Dry Matter) | Percent IP6 out of Total P | Phytase Activity (FTU/kg) * |
|---|---|---|---|---|---|
| Wheat 1 | 13 | 0.33 ± 0.02 | 0.22 ± 0.02 | 67 ± 4.8 | 1193 ± 223 |
| Wheat 2 | 18 | 0.40 ± 0.04 | 0.29 ± 0.04 | 73 ± 8.1 | 2886 ± 645 |
| Wheat 3 | 30 | 0.29 ± 0.03 | 0.23 ± 0.03 | 79 ± 0.07 | 1637 ± 275 |
| Barley 1 | 9 | 0.37 ± 0.02 | 0.22 ± 0.01 | 60 ± 2.4 | 582 ± 178 |
| Barley 2 | 15 | 0.42 ± 0.4 | 0.26 ± 0.03 | 63 ± 3.5 | 2323 ± 648 |
| Barley 3 | 21 | 0.31 ± 0.03 | 0.19 ± 0.02 | 61 ± 0.04 | 1016 ± 330 |
| Rye 1 | 2 | 0.36 (0.35-0.36) | 0.22 (0.20-0.23) | 61 ± (56 -66) | 5130 (4132-6127) |
| Rye 2 | 13 | 0.36 ± 0.02 | 0.24 ± 0.2 | 67 ± 5.0 | 6016 ± 1578 |
| Rye 3 | 6 | 0.34 ± 0.03 | 0.20 ± 0.01 | 59 ± 0.02 | 5147 ± 649 |
| Triticale 1 | 6 | 0.37 ± 0.02 | 0.25 ± 0.02 | 67 ± 3.7 | 1688 ± 227 |
| Triticale 2 | 12 | 0.40 ± 0.03 | 0.28 ± 0.03 | 70 ± 5.4 | 2799 ± 501 |
| Oats 1 | 6 | 0.36 ± 0.03 | 0.21 ± 0.04 | 59 ± 11 | 42 ± 50 |
| Oats 2 | 6 | 0.37 ± 0.01 | 0.25 ± 0.02 | 67 ± 5.4 | 496 ± 35 |
| Oats 3 | 9 | 0.29 ± 0.02 | 0.17 ± 0.03 | 59 ± 0.07 | 84 ± 39 |
| Maize 1 | 11 | 0.28 ± 0.03 | 0.19 ± 0.03 | 68 ± 5.9 | 15 ± 18 |
| Maize 3 | 7 | 0.32 ± 0.01 | 0.18 ± 0.01 | 78 ± 0.01 | 70 ± 7 |
| Rice 4 | 1 | 72 |
| Class | Enzyme | Km (µM) | Vmax (µmol/(min × mg)) | Kcat (s−1) | Kcat/Km (s−1M−1) | pH Optimum |
|---|---|---|---|---|---|---|
| PAP Phytases | rTaPAPhy_a1 2 | 35 | 223 | 279 | 796 × 104 | 5.5 |
| rTaPAPhy_b1 2 | 45 | 216 | 270 | 600 × 104 | 5 | |
| rHvPAPhy_a 2 | 36 | 208 | 260 | 722 × 104 | ||
| rHvPAPhy_b 2 | 46 | 202 | 253 | 550 × 104 | ||
| HAP Phytases | rTaPhyIIa2 1 | 246 | 4.5 | |||
| rHvPhyIIb 1 | 334 | 4.5 | ||||
| A. ficuum phytase 3 | 27 | - | 348 | 129 × 105 | 5.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Madsen, C.K.; Brinch-Pedersen, H. Molecular Advances on Phytases in Barley and Wheat. Int. J. Mol. Sci. 2019, 20, 2459. https://doi.org/10.3390/ijms20102459
Madsen CK, Brinch-Pedersen H. Molecular Advances on Phytases in Barley and Wheat. International Journal of Molecular Sciences. 2019; 20(10):2459. https://doi.org/10.3390/ijms20102459
Chicago/Turabian StyleMadsen, Claus Krogh, and Henrik Brinch-Pedersen. 2019. "Molecular Advances on Phytases in Barley and Wheat" International Journal of Molecular Sciences 20, no. 10: 2459. https://doi.org/10.3390/ijms20102459
APA StyleMadsen, C. K., & Brinch-Pedersen, H. (2019). Molecular Advances on Phytases in Barley and Wheat. International Journal of Molecular Sciences, 20(10), 2459. https://doi.org/10.3390/ijms20102459