Modulation of CAT-2B-Mediated l-Arginine Uptake and Nitric Oxide Biosynthesis in HCT116 Cell Line Through Biological Activity of 4′-Geranyloxyferulic Acid Extract from Quinoa Seeds

 ,
,  ,
,  ,
, 


Abstract
:1. Introduction
2. Results
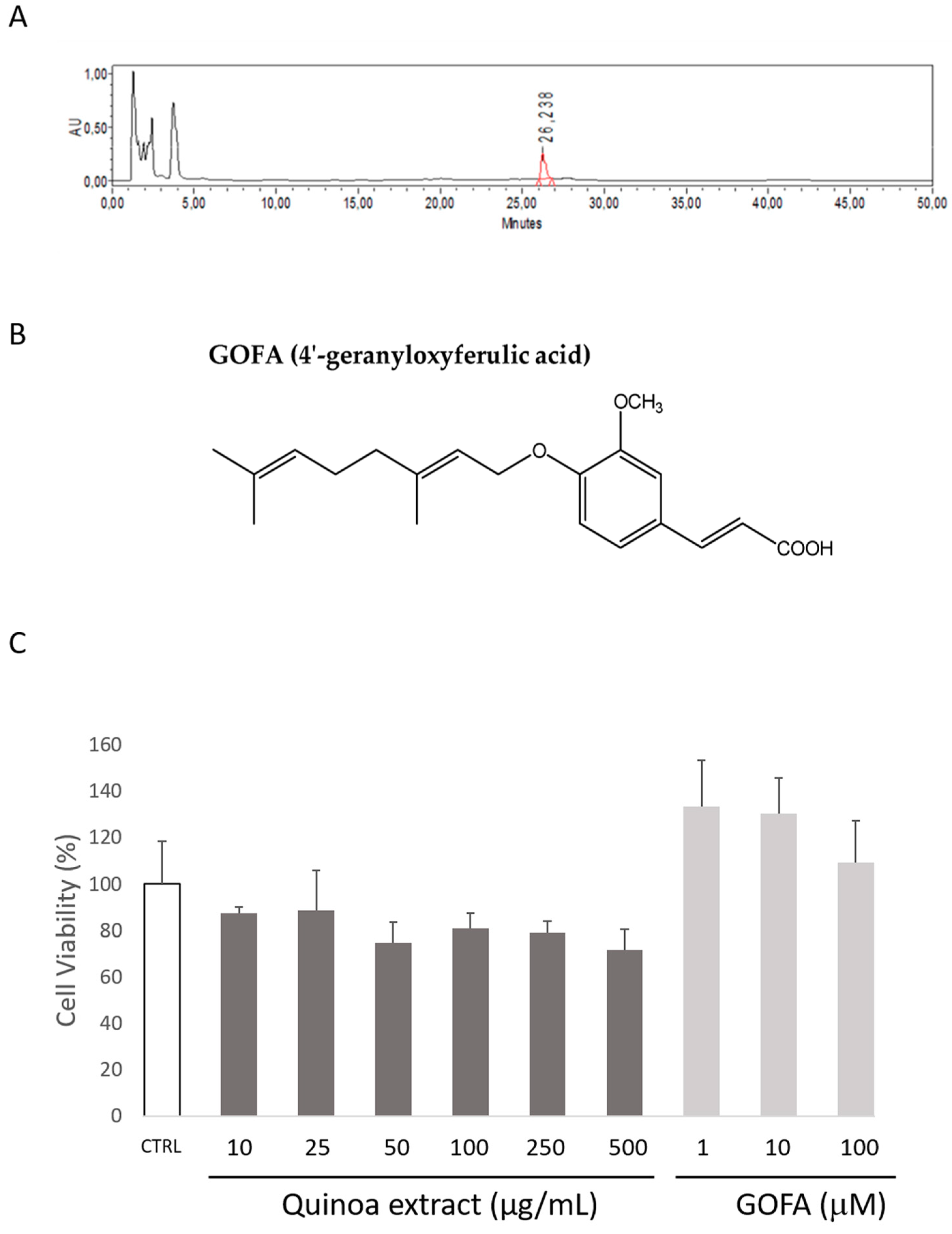
2.1. Quantification of GOFA in Roasted Quinoa Seeds Extracts
2.2. Quinoa and GOFA Effects on Cell Viability
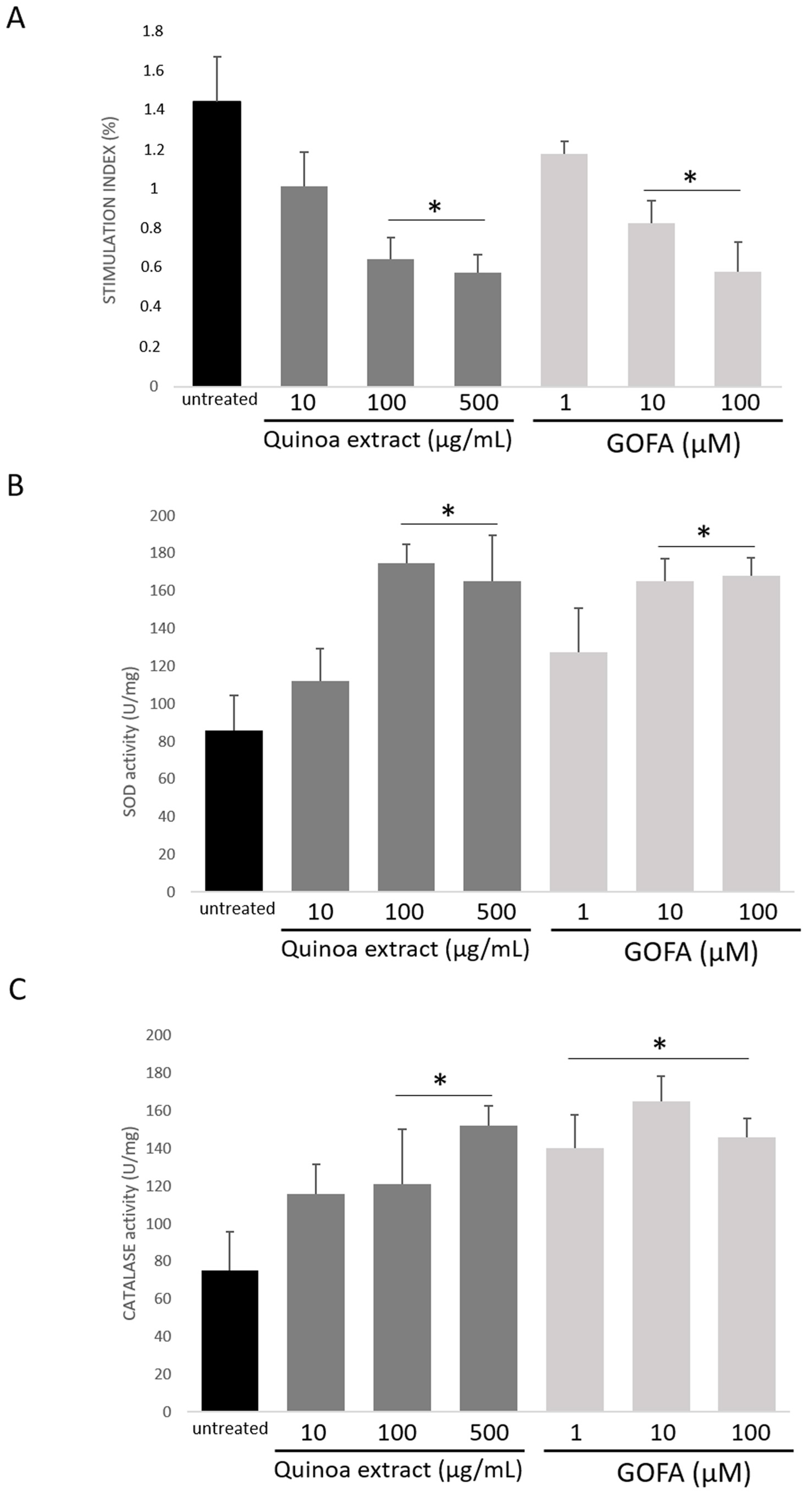
2.3. Quinoa and GOFA Effects on Redox Balance
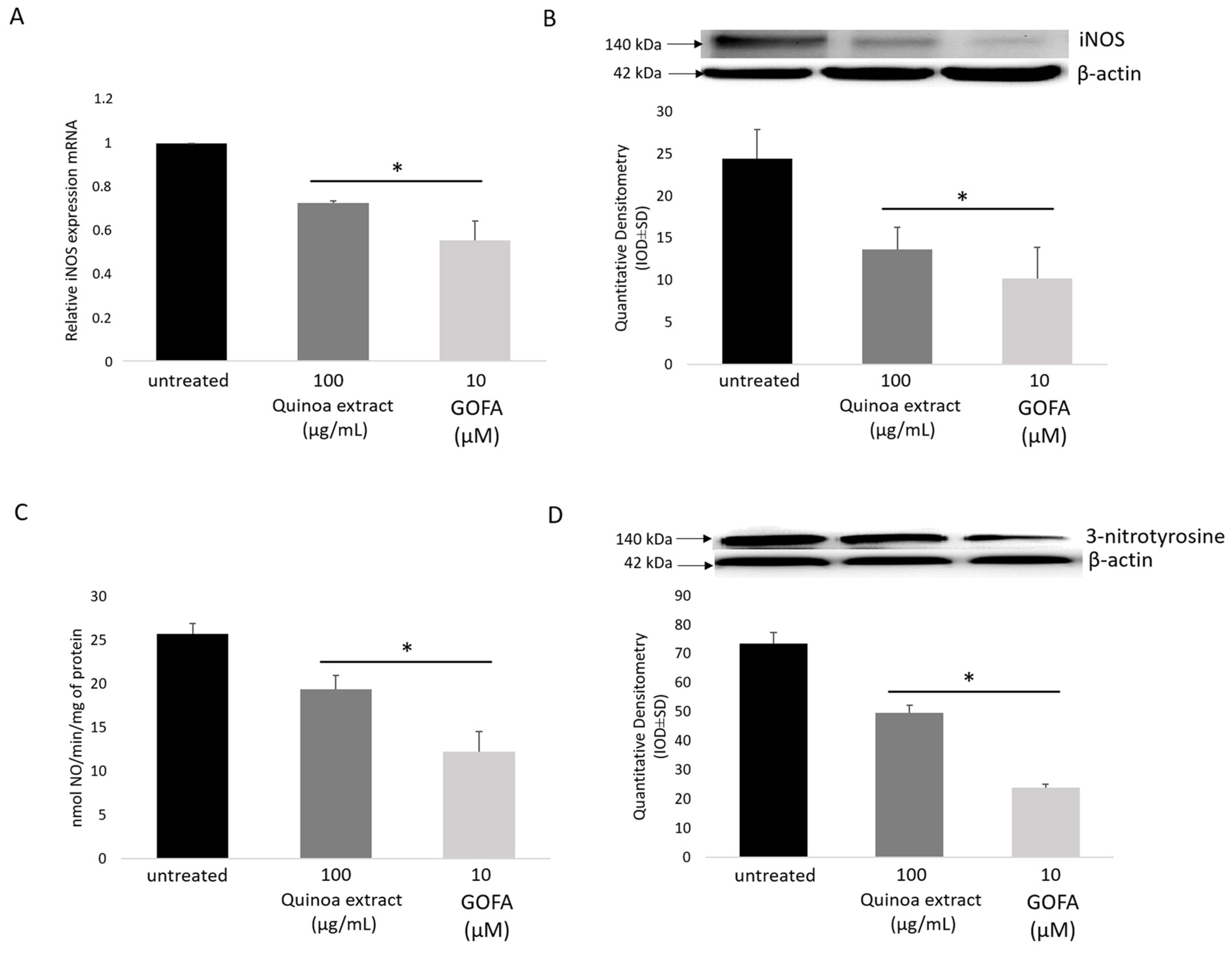
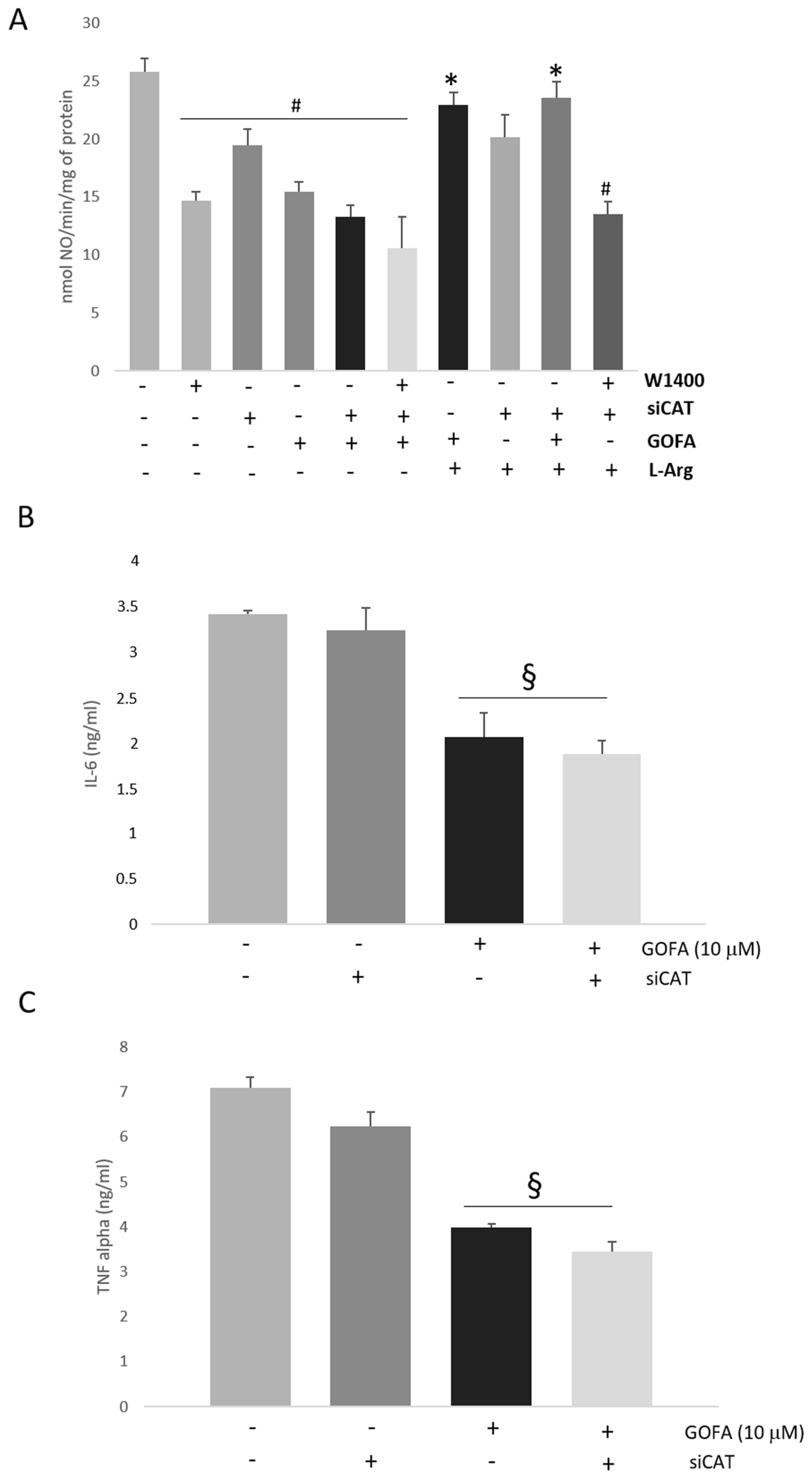
2.4. Quinoa and GOFA Effects on iNOS/NO/3-Nitrotyrosine
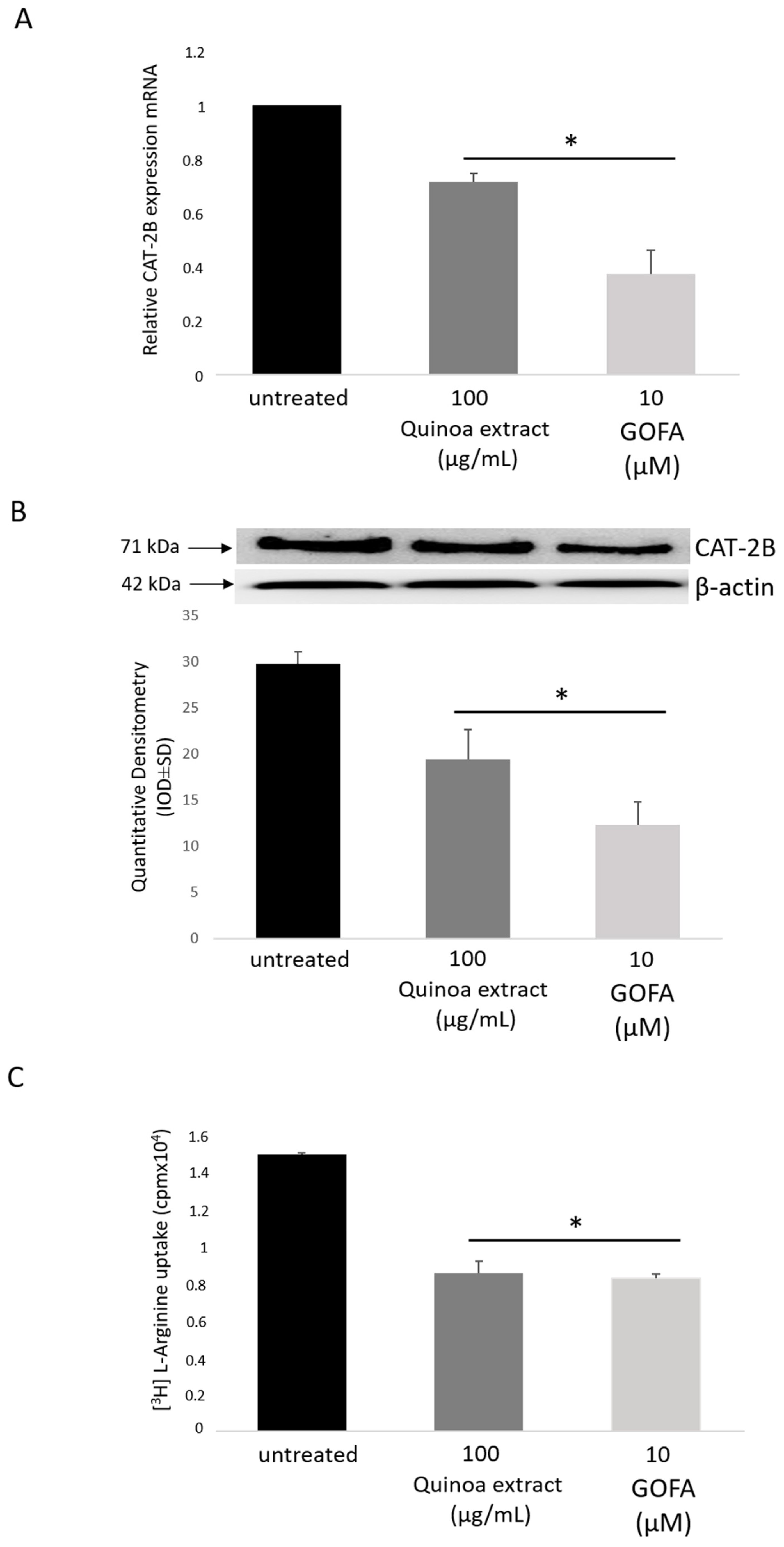
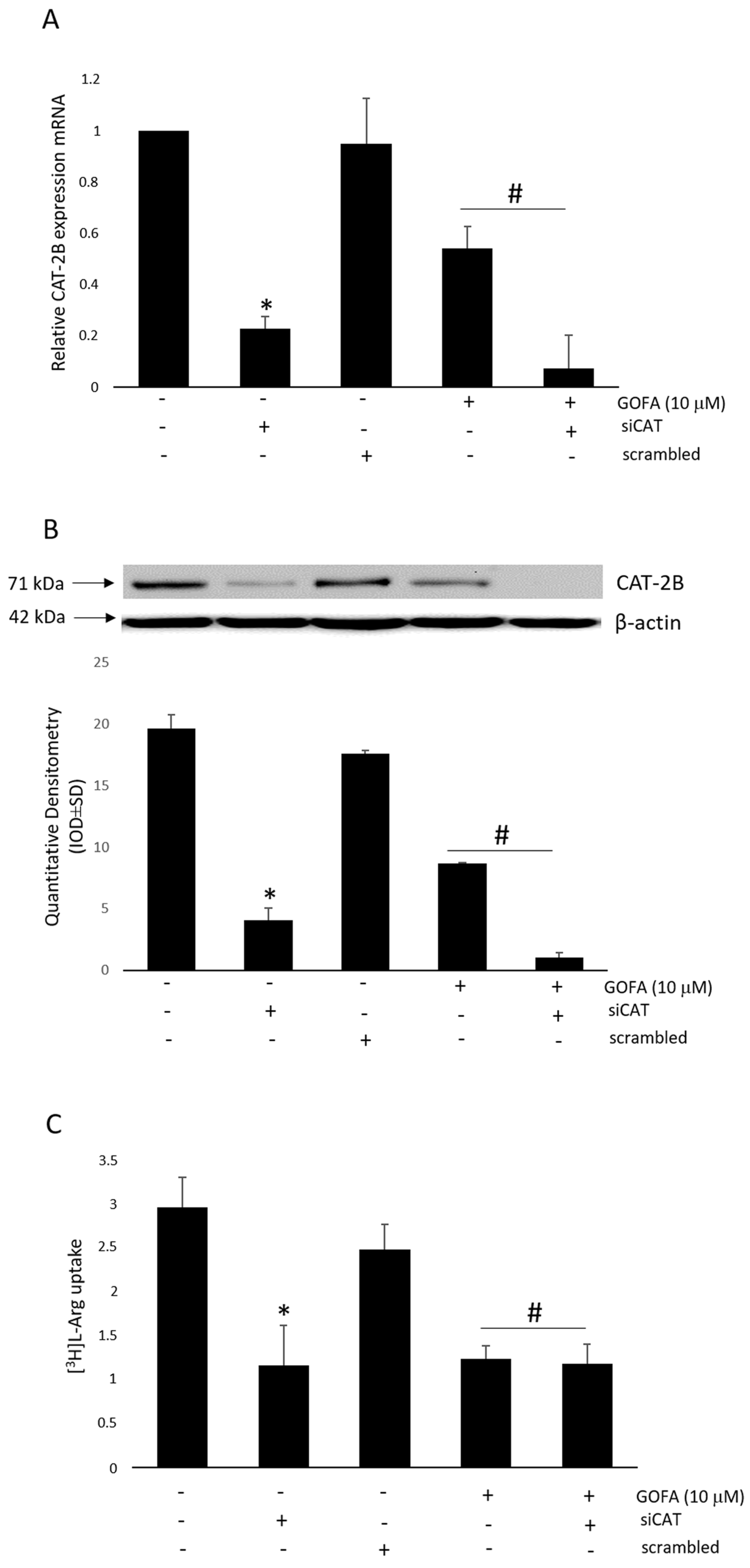
2.5. Effects of GOFA on the Expression of Cationic Amino Acid Transporters in HCT-116 Cells
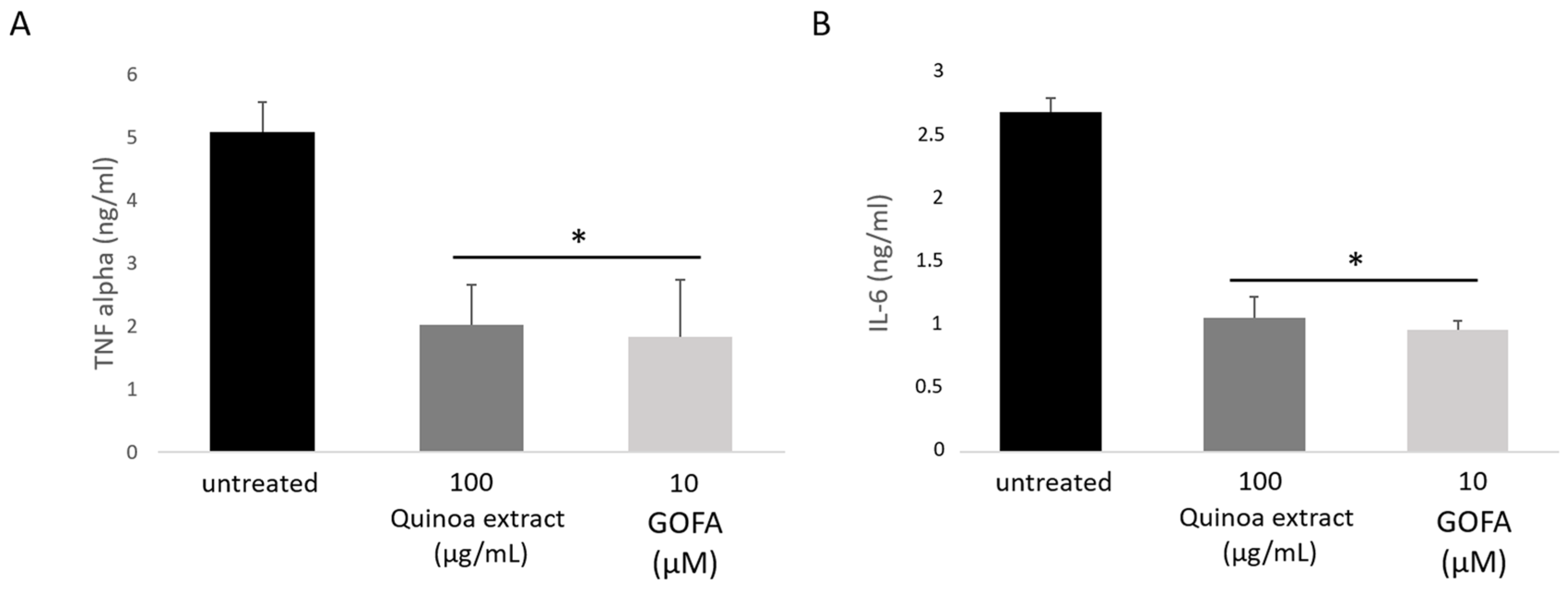
2.6. Quinoa Extract and GOFA Effects on the Pro-Inflammatory Markers
2.7. Effect of CAT-2B Silencing on the l-Arg/iNOS Pathways Levels
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Extraction Procedures and HPLC Analysis
4.3. Cell Culture, Assay for Cell Viability and Cytotoxicity
4.3.1. Cell Culture
4.3.2. MTT
4.3.3. NitroBlue-Tetrazolium Assay
4.4. Antioxidant Enzymes Activity
4.4.1. Superoxide Dismutase Activity Assay
4.4.2. Catalase Activity
4.5. Expression Analysis
4.5.1. Quantitative Real-Time PCR
4.5.2. Western Blot Analysis
4.6. Analysis of the l-Arg/iNOS/NO System
4.6.1. NOS Activity
4.6.2. Measurement of NO Release
4.6.3. l-Arg Uptake Assay
4.7. siRNA Transfection
4.8. Cytokine’s Levels Measurement
4.9. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, H.; Khor, T.O.; Shu, L.; Su, Z.Y.; Fuentes, F.; Lee, J.H.; Kong, A.N. Plants vs. cancer: A review on natural phytochemicals in preventing and treating cancers and their druggability. Anticancer Agents Med. Chem. 2012, 12, 1281–1305. [Google Scholar] [CrossRef] [PubMed]
- Nobili, S.; Lippi, D.; Witort, E.; Donnini, M.; Bausi, L.; Mini, E.; Capaccioli, S. Natural compounds for cancer treatment and prevention. Pharmacol. Res. 2009, 59, 365–378. [Google Scholar] [CrossRef] [PubMed]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miar, A.; Hevia, D.; Muñoz-Cimadevilla, H.; Astudillo, A.; Velasco, J.; Sainz, R.M.; Mayo, J.C. Manganese superoxide dismutase (SOD2/MnSOD)/catalase and SOD2/GPx1 ratios as biomarkers for tumor progression and metastasis in prostate, colon, and lung cancer. Free Radic. Biol. Med. 2015, 85, 45–55. [Google Scholar] [CrossRef]
- Speranza, L.; Franceschelli, S.; Pesce, M.; Vinciguerra, I.; De Lutiis, M.A.; Grilli, A.; Felaco, M.; Patruno, A. Phosphodiesterase type-5 inhibitor and oxidative stress. Int. J. Immunopathol. Pharmacol. 2005, 21, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Chen, L.; Ma, K.; Zhao, Y.; Liu, X.; Wang, Y.; Liu, M.; Liang, S.; Zhu, H.; Xu, N. Polarization of macrophages in the tumor microenvironment is influenced by EGFR signaling within colon cancer cells. Oncotarget 2016, 7, 75366–75378. [Google Scholar] [CrossRef] [Green Version]
- Patil, M.D.; Bhaumik, J.; Babykutty, S.; Banerjee, U.C.; Fukumura, D. Arginine dependence of tumor cells: Targeting a chink in cancer’s armor. Oncogene 2016, 35, 4957–4972. [Google Scholar] [CrossRef]
- Sans-Fons, M.G.; Yeramian, A.; Pereira-Lopes, S.; Santamaría-Babi, L.F.; Modolell, M.; Lloberas, J.; Celada, A. Arginine transport is impaired in C57Bl/6 mouse macrophages as a result of a deletion in the promoter of Slc7a2 (CAT2), and susceptibility to Leishmania infection is reduced. J. Infect. Dis. 2013, 207, 1684–1693. [Google Scholar] [CrossRef]
- Huang, Q.; Huang, C.; Zhao, Y.; Wang, B.; Ren, J.; Li, N.; Li, J. LPS-stimulated RAW264.7 macrophage CAT-2-mediated l-Arginine uptake and nitric oxide biosynthesis is inhibited by omega fatty acid lipid emulsion. J. Surg. Res. 2013, 179, e211–e217. [Google Scholar] [CrossRef]
- Speranza, L.; Franceschelli, S.; Pesce, M.; Menghini, L.; Patruno, A.; Vinciguerra, I.; De Lutiis, M.A.; Felaco, M.; Felaco, P.; Grilli, A. Anti-inflammatory properties of the plant Verbascum mallophorum. J. Biol. Regul. Homeost. Agents 2009, 23, 189–195. [Google Scholar]
- Morry, J.; Ngamcherdtrakul, W.; Yantasee, W. Oxidative stress in cancer and fibrosis: Opportunity for therapeutic intervention with antioxidant compounds, enzymes, and nanoparticles. Redox. Biol. 2017, 11, 240–253. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Tsao, R. Phytochemicals in quinoa and amaranth grains and their antioxidant, anti-inflammatory and potential health beneficial effects: A review. Mol. Nutr. Food Res. 2017, 61, 1600767. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Zhu, F. Quinoa starch: Structure, properties, and applications. Carbohydr. Polym. 2018, 181, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Nowak, V.; Du, J.; Charrondière, U.R. Assessment of the nutritional composition of quinoa (Chenopodium quinoa Willd). Food Chem. 2016, 193, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Bruyere, C.; Genovese, S.; Lallemand, B.; Ionescu-Motatu, A.; Curini, M.; Kiss, R.; Epifano, F. Growth inhibitory activities of oxyprenylated and non-prenylated naturally occurring phenylpropanoids in cancer cell lines. Bioorg. Med. Chem. Lett. 2011, 21, 4174–4179. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; de Azevedo, M.B.; Durán, N.; Alderete, J.B.; Epifano, F.; Genovese, S.; Tanaka, M.; Tanaka, T.; Curini, M. Colorectal cancer chemoprevention by 2 beta-cyclodextrin inclusion compounds of auraptene and 4′-geranyloxyferulic acid. Int. J. Cancer. 2010, 126, 830–840. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Kochi, T.; Shirakami, Y.; Genovese, S.; Epifano, F.; Fiorito, S.; Mori, T.; Tanaka, T.; Moriwaki, H. A newly synthesized compound, 4′-geranyloxyferulic acid-N(omega)-nitro-l-arginine methyl ester suppresses inflammation-associated colorectal carcinogenesis in male mice. Int. J. Cancer 2014, 135, 774–784. [Google Scholar] [CrossRef]
- Fiorito, S.; Epifano, F.; Palmisano, R.; Genovese, S.; Taddeo, V.A. A re-investigation of the phytochemical composition of the edible herb Amaranthus retroflexus L. J. Pharm. Biomed. Anal. 2017, 143, 183–187. [Google Scholar] [CrossRef]
- Franceschelli, S.; Pesce, M.; Ferrone, A.; Patruno, A.; Pasqualone, L.; Carlucci, G.; Ferrone, V.; Carlucci, M.; de Lutiis, M.A.; Grilli, A.; et al. A Novel Biological Role of α-Mangostin in Modulating Inflammatory Response Through the Activation of SIRT-1 Signaling Pathway. J. Cell. Physiol. 2016, 231, 2439–2451. [Google Scholar] [CrossRef]
- Pesce, M.; Franceschelli, S.; Ferrone, A.; De Lutiis, M.A.; Patruno, A.; Grilli, A.; Felaco, M.; Speranza, L. Verbascoside down-regulates some pro-inflammatory signal transduction pathways by increasing the activity of tyrosine phosphatase SHP-1 in the U937 cell line. J. Cell. Mol. Med. 2015, 19, 1548–1556. [Google Scholar] [CrossRef]
- Franceschelli, S.; Pesce, M.; Ferrone, A.; Gatta, D.M.; Patruno, A.; De Lutiis, M.A.; Quiles, J.L.; Grilli, A.; Felaco, M.; Speranza, L. Biological Effect of Licochalcone C on the Regulation of PI3K/Akt/eNOS and NF-κB/iNOS/NO Signaling Pathways in H9c2 Cells in Response to LPS Stimulation. Int. J. Mol. Sci. 2017, 18, 690. [Google Scholar] [CrossRef] [PubMed]
- MacCallum, C.; Skandarajah, A.; Gibbs, P.; Hayes, I. The Value of Clinical Colorectal Cancer Registries in Colorectal Cancer Research: A Systematic Review. JAMA Surg. 2018, 153, 841–849. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.R.; Chang, D.K. Colorectal cancer in inflammatory bowel disease: The risk, pathogenesis, prevention and diagnosis. World J. Gastroenterol. 2014, 20, 9872–9881. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.; Duong, H.Q. The molecular characteristics of colorectal cancer: Implications for diagnosis and therapy. Oncol. Lett. 2018, 16, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Janakiram, N.B.; Rao, C.V. The role of inflammation in colon cancer. Adv. Exp. Med. Biol. 2014, 816, 25–52. [Google Scholar] [CrossRef] [PubMed]
- Rossi, M.; Mirbagheri, S.; Keshavarzian, A.; Bishehsari, F. Nutraceuticals in Colorectal Cancer: A Mechanistic Approach. Eur. J. Pharmacol. 2018, 833, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Virgili, F.; Marino, M. Regulation of cellular signals from nutritional molecules: A specific role for phytochemicals, beyond antioxidant activity. Free Radic. Biol. Med. 2008, 45, 1205–1216. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.L.; Norhaizan, M.E.; Liew, W.P. Nutrients and Oxidative Stress: Friend or Foe? Oxid. Med. Cell. Longev. 2018, 2018, 9719584. [Google Scholar] [CrossRef]
- Genovese, S.; Fiorito, S.; Epifano, F.; Taddeo, V.A. A Novel Class of Emerging Anticancer Compounds: Oxyprenylated Secondary Metabolites from Plants and Fungi. Curr. Med. Chem. 2015, 22, 3426–3433. [Google Scholar] [CrossRef]
- Chaiswing, L.; St Clair, W.H.; St Clair, D.K. Redox Paradox: A Novel Approach to Therapeutics-sistant Cancer. Antioxid. Redox Signal. 2018, 29, 1237–1272. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget. 2017, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed]
- Pais, R.; Rievaj, J.; Meek, C.; De Costa, G.; Jayamaha, S.; Alexander, R.T.; Reimann, F.; Gribble, F. Role of enteroendocrine L-cells in arginine vasopressin-mediated inhibition of colonic anion secretion. J. Physiol. 2016, 594, 4865–4878. [Google Scholar] [CrossRef] [PubMed]
- Leung, K.T.; Chan, K.Y.; Ma, T.P.; Yu, J.W.; Tong, J.H.; Tam, Y.H.; Cheung, H.M.; To, K.F.; Lam, H.S.; Lee, K.H.; et al. Dysregulated expression of arginine metabolic enzymes in human intestinal tissues of necrotizing enterocolitis and response of CaCO2 cells to bacterial components. J. Nutr. Biochem. 2016, 29, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Taddeo, V.A.; Genovese, S.; Carlucci, G.; Ferrone, V.; Patruno, A.; Ferrone, A.; de Medina, P.; Fiorito, S.; Epifano, F. Quantitative profiling of 4′-geranyloxyferulic acid and its 7 conjugate with l-nitroarginine methyl ester in mononuclear cells by high-performance 8 liquid chromatography with fluorescence detection. J. Pharm. Biomed. Anal. 2017, 133, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Pesce, M.; Ferrone, A.; Rizzuto, A.; Tatangelo, R.; Iezzi, I.; Ladu, S.; Franceschelli, S.; Speranza, L.; Patruno, A.; Felaco, M.; et al. The SHP-1 expression is associated with cytokines and psychopathological status in unmedicated first episode schizophrenia patients. Brain Behav. Immun. 2014, 41, 251–260. [Google Scholar] [CrossRef]
- Franceschelli, S.; Gatta, D.M.; Pesce, M.; Ferrone, A.; Patruno, A.; de Lutiis, M.A.; Grilli, A.; Felaco, M.; Croce, F.; Speranza, L. New Approach in Translational Medicine: Effects of Electrolyzed Reduced Water (ERW) on NF-κB/iNOS Pathway in U937 Cell Line under Altered Redox State. Int. J. Mol. Sci. 2016, 17, 1461. [Google Scholar] [CrossRef]
- Patruno, A.; Franceschelli, S.; Pesce, M.; Maccallini, C.; Fantacuzzi, M.; Speranza, L.; Ferrone, A.; De Lutiis, M.A.; Ricciotti, E.; Amoroso, R.; et al. Novel aminobenzyL-Acetamidine derivative modulate the differential regulation of NOSs in LPS induced inflammatory response: Role of PI3K/Akt pathway. Biochim. Biophys. Acta Gen. Subj. 2012, 1820, 2095–2104. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Entry | Linearity Range (μg/mL) | Calibration Curve | Weighting Factor | Determination Coefficient (r2) | Rt (min) * |
|---|---|---|---|---|---|
| GOFA | 1–100 | Y = 24142x + 3054 | 1/x2 | 0.9993 | 41.77 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Franceschelli, S.; Gatta, D.M.P.; Pesce, M.; Ferrone, A.; Quiles, J.L.; Genovese, S.; Epifano, F.; Fiorito, S.; Taddeo, V.A.; Patruno, A.; et al. Modulation of CAT-2B-Mediated l-Arginine Uptake and Nitric Oxide Biosynthesis in HCT116 Cell Line Through Biological Activity of 4′-Geranyloxyferulic Acid Extract from Quinoa Seeds. Int. J. Mol. Sci. 2019, 20, 3262. https://doi.org/10.3390/ijms20133262
Franceschelli S, Gatta DMP, Pesce M, Ferrone A, Quiles JL, Genovese S, Epifano F, Fiorito S, Taddeo VA, Patruno A, et al. Modulation of CAT-2B-Mediated l-Arginine Uptake and Nitric Oxide Biosynthesis in HCT116 Cell Line Through Biological Activity of 4′-Geranyloxyferulic Acid Extract from Quinoa Seeds. International Journal of Molecular Sciences. 2019; 20(13):3262. https://doi.org/10.3390/ijms20133262
Chicago/Turabian StyleFranceschelli, Sara, Daniela Maria Pia Gatta, Mirko Pesce, Alessio Ferrone, José Luis Quiles, Salvatore Genovese, Francesco Epifano, Serena Fiorito, Vito Alessandro Taddeo, Antonia Patruno, and et al. 2019. "Modulation of CAT-2B-Mediated l-Arginine Uptake and Nitric Oxide Biosynthesis in HCT116 Cell Line Through Biological Activity of 4′-Geranyloxyferulic Acid Extract from Quinoa Seeds" International Journal of Molecular Sciences 20, no. 13: 3262. https://doi.org/10.3390/ijms20133262
APA StyleFranceschelli, S., Gatta, D. M. P., Pesce, M., Ferrone, A., Quiles, J. L., Genovese, S., Epifano, F., Fiorito, S., Taddeo, V. A., Patruno, A., Grilli, A., Felaco, M., & Speranza, L. (2019). Modulation of CAT-2B-Mediated l-Arginine Uptake and Nitric Oxide Biosynthesis in HCT116 Cell Line Through Biological Activity of 4′-Geranyloxyferulic Acid Extract from Quinoa Seeds. International Journal of Molecular Sciences, 20(13), 3262. https://doi.org/10.3390/ijms20133262