Conceptual Evolution of Cell Signaling



Abstract
:1. Introduction
2. Components of Cell Signaling
2.1. Ligands or Signals
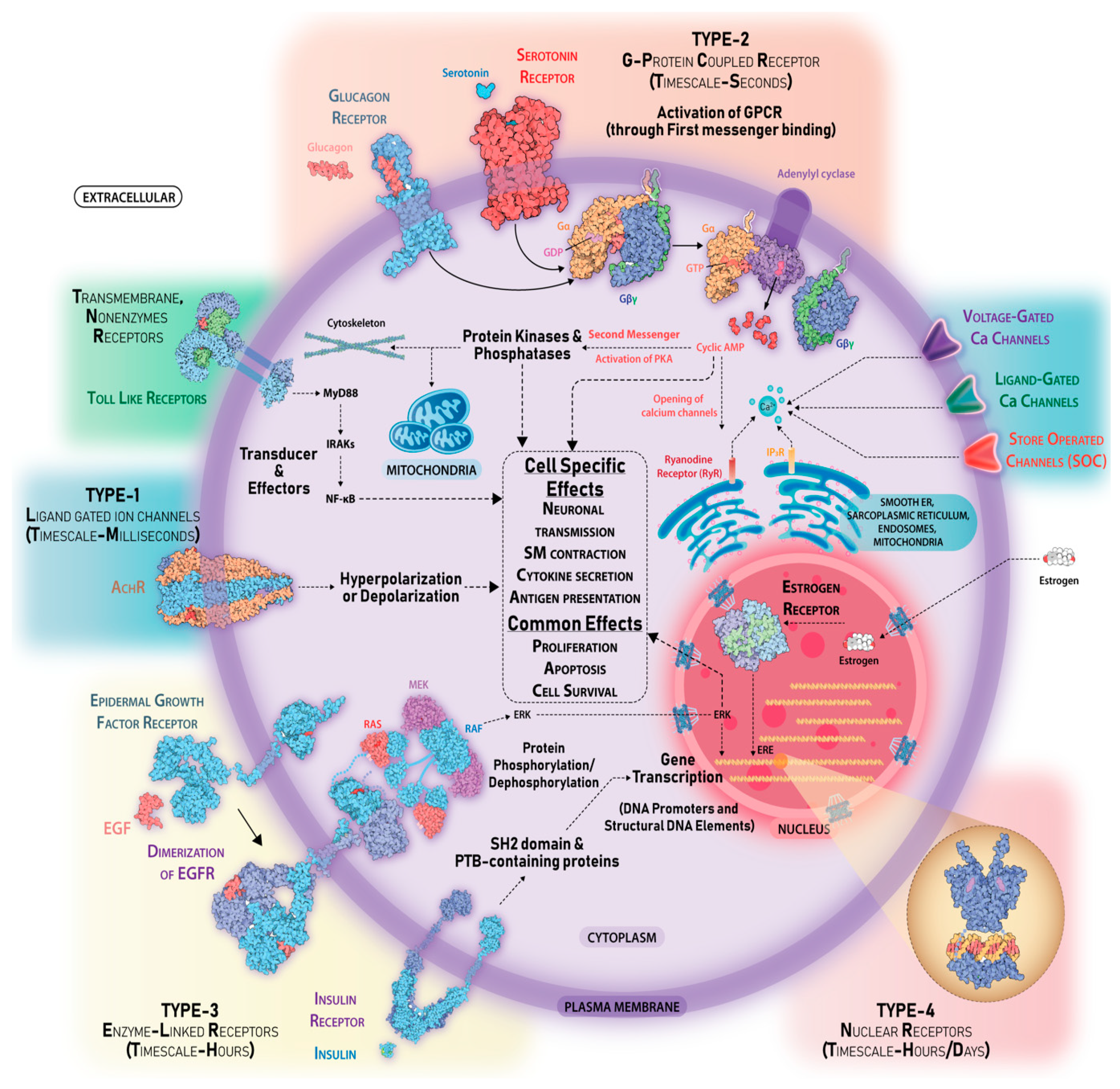
2.2. Receptors
2.3. Specificity in Signaling
2.3.1. Lipids in Signaling
2.3.2. Signaling Domains
2.3.3. Common Signaling Domains Found in Proteins
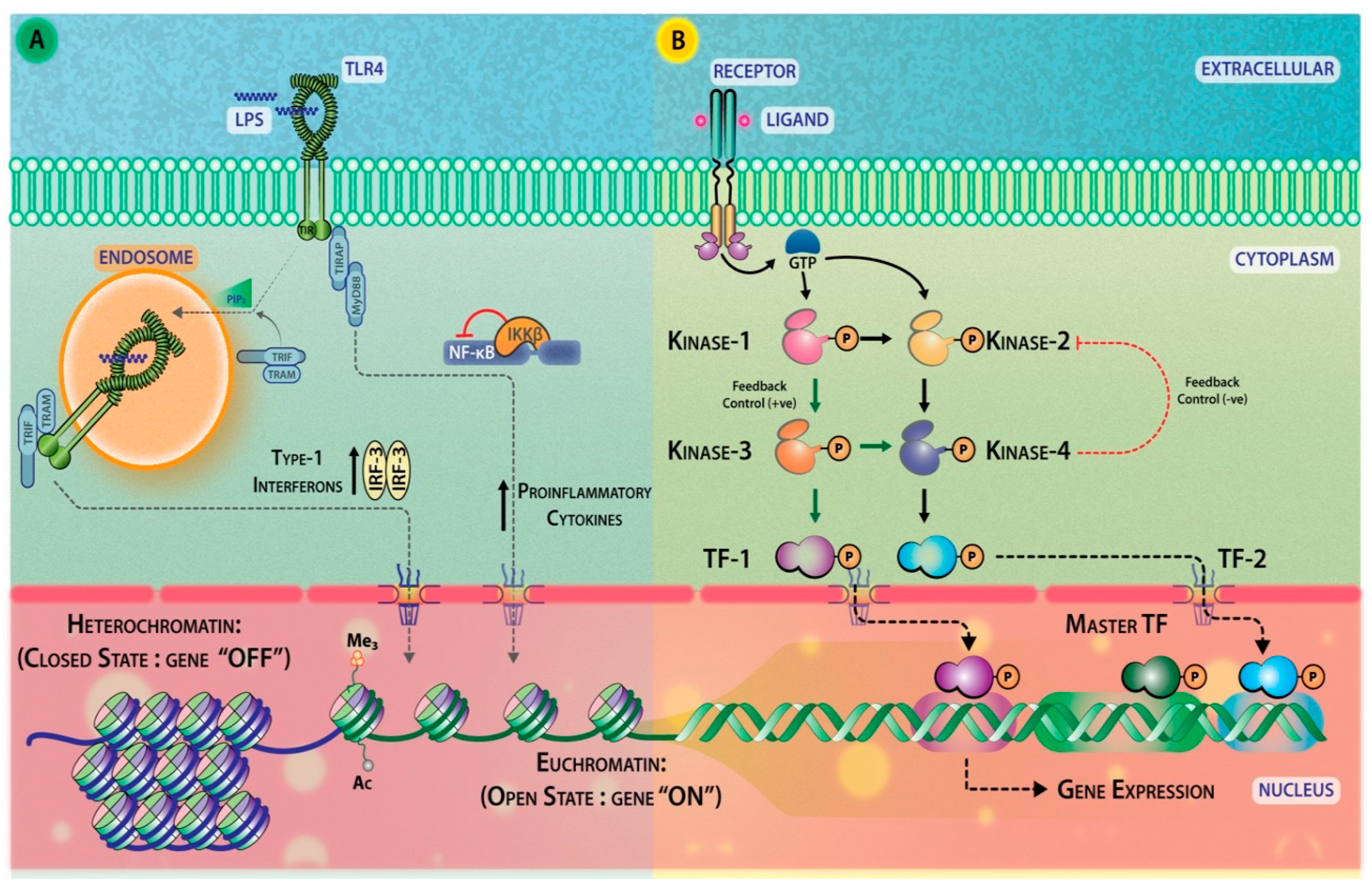
2.4. Signal Transducers
2.5. Second Messengers
2.6. Transcription Factors
3. Directionality of Signaling
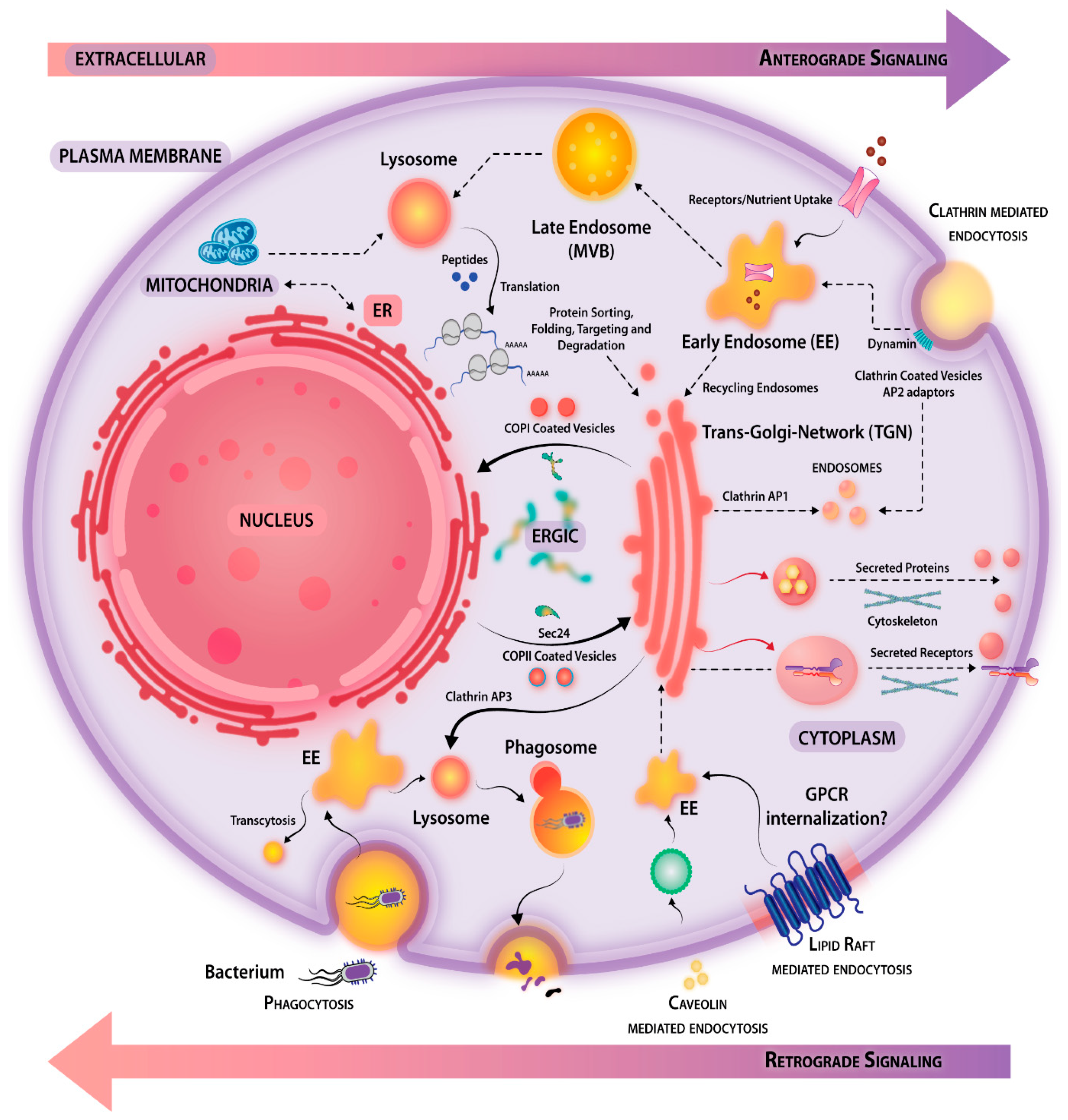
3.1. Retrograde and Anterograde Signaling
3.2. Compartmentalized Signaling
3.3. Cell Adhesion and Membrane Protrusion
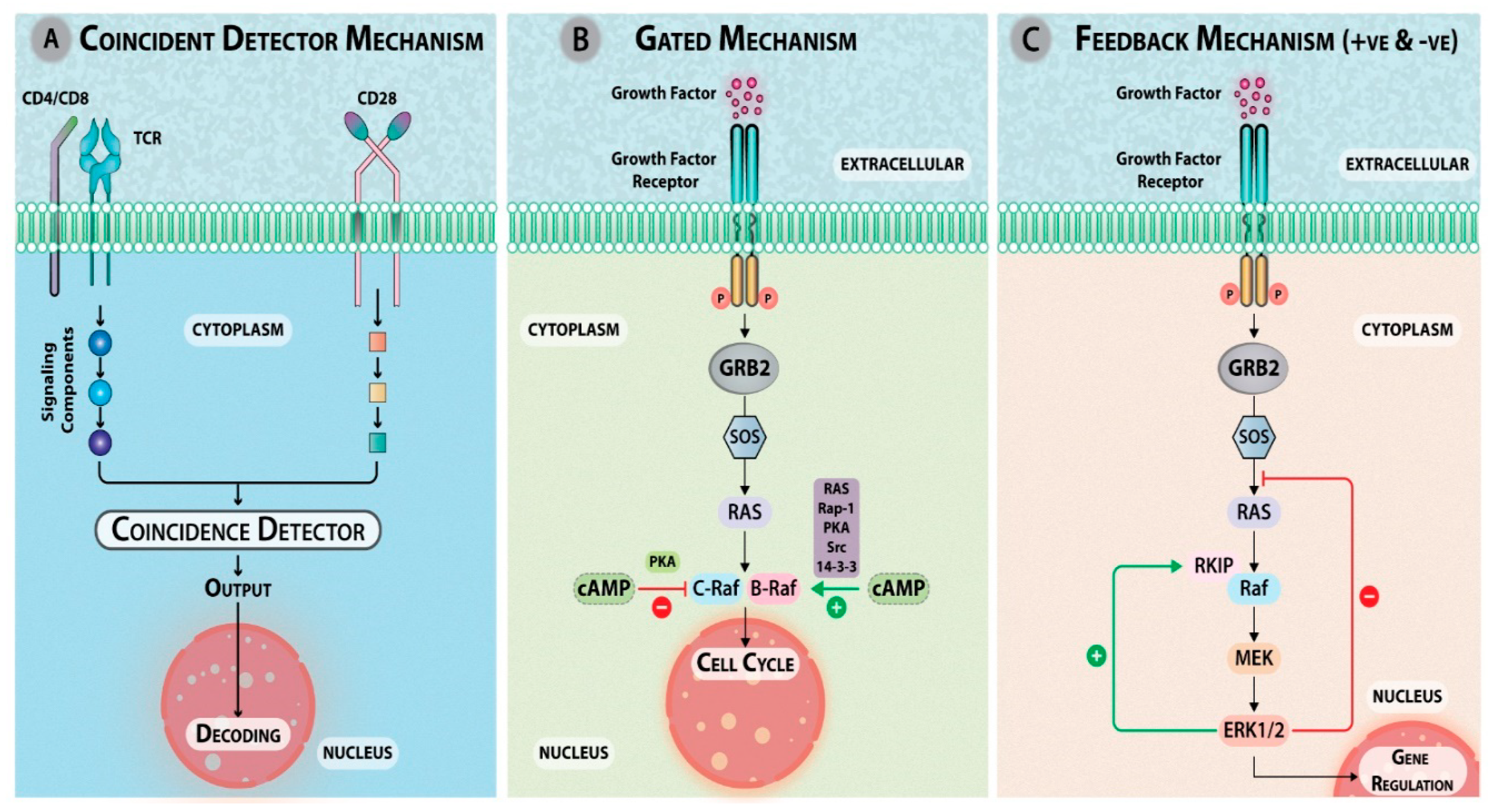
4. Complexity in Signaling
4.1. Interactions between Pathways
4.2. Post-Translational Modifications (PTMs)
4.3. Engagement of Different Signaling Modules
5. Translational Value of Understanding Signal Transduction
6. An Evolutionary Perspective of Signaling
7. Conclusion: Past, Present, and Future of Cell Signaling
7.1. Major Discoveries in the Field of Signal Transduction
7.2. Current Methods in Cell Signaling
Mathematical Modeling, Systems Biology, and –OMICS
7.3. Future of Cell Signaling
Author Contributions
Conflicts of Interest
References
- Darwin, C. The power of movement in plants; D. Appleton and Company: New York, NY, USA, 1897. [Google Scholar]
- Langley, J.N. Observations on the physiological action of extracts of the supra-renal bodies. J. Physiol. 1901, 27, 237–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starling, E.H. Discussion on the therapeutic value of hormones. Proc. R. Soc. Med. 1914, 7, 29–31. [Google Scholar] [PubMed]
- Levi-Montalcini, R.; Hamburger, V. Selective growth stimulating effects of mouse sarcoma on the sensory and sympathetic nervous system of the chick embryo. J. Exp. Zool. 1951, 116, 321–361. [Google Scholar] [CrossRef] [PubMed]
- Hokin, M.R.; Hokin, L.E. Enzyme secretion and the incorporation of P32 into phospholipides of pancreas slices. J. Biol. Chem. 1953, 203, 967–977. [Google Scholar] [CrossRef]
- Krebs, E.G.; Fischer, E.H. The phosphorylase b to a converting enzyme of rabbit skeletal muscle. Biochim. Biophys. Acta 1956, 20, 150–157. [Google Scholar] [CrossRef]
- Sutherland, E.W.; Rall, T.W. The properties of an adenine ribonucleotide produced with cellular particles, ATP, Mg++, and epinephrine or glucagon. J. Am. Chem. Soc. 1957, 79, 3608. [Google Scholar] [CrossRef]
- Cohen, S. Isolation of a mouse submaxillary gland protein accelerating incisor eruption and eyelid opening in the new-born animal. J. Biol. Chem. 1962, 237, 1555–1562. [Google Scholar]
- Ross, E.M.; Gilman, A.G. Resolution of some components of adenylate cyclase necessary for catalytic activity. J. Biol. Chem. 1977, 252, 6966–6969. [Google Scholar]
- Carswell, E.A.; Old, L.J.; Kassel, R.; Green, S.; Fiore, N.; Williamson, B. An endotoxin-induced serum factor that causes necrosis of tumors. Proc. Natl. Acad. Sci. USA 1975, 72, 3666–3670. [Google Scholar] [CrossRef]
- Hunter, T.; Sefton, B.M. Transforming gene product of Rous sarcoma virus phosphorylates tyrosine. Proc. Natl. Acad. Sci. USA 1980, 77, 1311–1315. [Google Scholar] [CrossRef] [Green Version]
- Hildebrand, E. What does Halobacterium tell us about photoreception? Biophys. Struct. Mech. 1977, 3, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Gough, N.R. Neuroprotective Mitochondrial Glutamate Receptors. Sci. Signal. 2012, 5, ec272. [Google Scholar] [CrossRef]
- Psarra, A.G.; Sekeris, C.E. Steroid and thyroid hormone receptors in mitochondria. IUBMB Life 2008, 60, 210–223. [Google Scholar] [CrossRef] [PubMed]
- Tesei, A.; Cortesi, M.; Zamagni, A.; Arienti, C.; Pignatta, S.; Zanoni, M.; Paolillo, M.; Curti, D.; Rui, M.; Rossi, D. Sigma Receptors as Endoplasmic Reticulum Stress “Gatekeepers” and their Modulators as Emerging New Weapons in the Fight Against Cancer. Front. Pharmacol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Brown, W.J.; Farquhar, M.G. The mannose-6-phosphate receptor for lysosomal enzymes is concentrated in cis Golgi cisternae. Cell 1984, 36, 295–307. [Google Scholar] [CrossRef]
- Gao, J.; Wang, Y.; Cai, M.; Pan, Y.; Xu, H.; Jiang, J.; Ji, H.; Wang, H. Mechanistic insights into EGFR membrane clustering revealed by super-resolution imaging. Nanoscale 2015, 7, 2511–2519. [Google Scholar] [CrossRef] [PubMed]
- Kupfer, A.; Dennert, G.; Singer, S.J. The reorientation of the Golgi apparatus and the microtubule-organizing center in the cytotoxic effector cell is a prerequisite in the lysis of bound target cells. J. Mol. Cell. Immunol. JMCI 1985, 2, 37–49. [Google Scholar] [PubMed]
- Chen, I.-J.; Chen, H.-L.; Demetriou, M. Lateral compartmentalization of T cell receptor versus CD45 by galectin-N-glycan binding and microfilaments coordinate basal and activation signaling. J. Biol. Chem. 2007, 282, 35361–35372. [Google Scholar] [CrossRef] [PubMed]
- Kusumi, A.; Tsunoyama, T.A.; Hirosawa, K.M.; Kasai, R.S.; Fujiwara, T.K. Tracking single molecules at work in living cells. Nat. Chem. Biol. 2014, 10, 524. [Google Scholar] [CrossRef]
- Sherman, E.; Barr, V.; Manley, S.; Patterson, G.; Balagopalan, L.; Akpan, I.; Regan, C.K.; Merrill, R.K.; Sommers, C.L.; Lippincott-Schwartz, J. Functional nanoscale organization of signaling molecules downstream of the T cell antigen receptor. Immunity 2011, 35, 705–720. [Google Scholar] [CrossRef]
- Cheng, P.C.; Brown, B.K.; Song, W.; Pierce, S.K. Translocation of the B cell antigen receptor into lipid rafts reveals a novel step in signaling. J. Immunol. 2001, 166, 3693–3701. [Google Scholar] [CrossRef] [PubMed]
- Field, K.A.; Holowka, D.; Baird, B. Fc epsilon RI-mediated recruitment of p53/56lyn to detergent-resistant membrane domains accompanies cellular signaling. Proc. Natl. Acad. Sci. USA 1995, 92, 9201–9205. [Google Scholar] [CrossRef]
- Brown, D.A.; London, E. Functions of lipid rafts in biological membranes. Annu. Rev. Cell Dev. Biol. 1998, 14, 111–136. [Google Scholar] [CrossRef] [PubMed]
- Hansen, G.H.; Immerdal, L.; Thorsen, E.; Niels-Christiansen, L.-L.; Nystrøm, B.T.; Demant, E.J.F.; Danielsen, E.M. Lipid rafts exist as stable cholesterol-independent microdomains in the brush border membrane of enterocytes. J. Biol. Chem. 2001, 276, 32338–32344. [Google Scholar] [CrossRef] [PubMed]
- Varshney, P.; Yadav, V.; Saini, N. Lipid rafts in immune signalling: Current progress and future perspective. Immunology 2016, 149, 13–24. [Google Scholar] [CrossRef]
- Grassmé, H.; Jendrossek, V.; Bock, J.; Riehle, A.; Gulbins, E. Ceramide-rich membrane rafts mediate CD40 clustering. J. Immunol. 2002, 168, 298–307. [Google Scholar] [CrossRef]
- Jin, S.; Zhou, F.; Katirai, F.; Li, P.-L. Lipid raft redox signaling: Molecular mechanisms in health and disease. Antioxid. Redox Signal. 2011, 15, 1043–1083. [Google Scholar] [CrossRef]
- Rajendran, L.; Simons, K. Lipid rafts and membrane dynamics. J. Cell Sci. 2005, 118, 1099–1102. [Google Scholar] [CrossRef] [Green Version]
- Wei, N.; Tsuge, T.; Serino, G.; Dohmae, N.; Takio, K.; Matsui, M.; Deng, X.-W. The COP9 complex is conserved between plants and mammals and is related to the 26S proteasome regulatory complex. Curr. Biol. 1998, 8, 919–924. [Google Scholar] [CrossRef] [Green Version]
- Su, H.; Li, J.; Osinska, H.; Li, F.; Robbins, J.; Liu, J.; Wei, N.; Wang, X. The COP9 signalosome is required for autophagy, proteasome-mediated proteolysis, and cardiomyocyte survival in adult mice. Circ. Hear. Fail. 2013, 6, 1049–1057. [Google Scholar] [CrossRef]
- Huang, E.J.; Reichardt, L.F. Trk receptors: Roles in neuronal signal transduction. Annu. Rev. Biochem. 2003, 72, 609–642. [Google Scholar] [CrossRef] [PubMed]
- Bae, Y.S.; Sung, J.-Y.; Kim, O.-S.; Kim, Y.J.; Hur, K.C.; Kazlauskas, A.; Rhee, S.G. Platelet-derived growth factor-induced H2O2 production requires the activation of phosphatidylinositol 3-kinase. J. Biol. Chem. 2000, 275, 10527–10531. [Google Scholar] [CrossRef] [PubMed]
- Vanhaesebroeck, B.; Stephens, L.; Hawkins, P. PI3K signalling: The path to discovery and understanding. Nat. Rev. Mol. Cell Biol. 2012, 13, 195. [Google Scholar] [CrossRef] [PubMed]
- Funamoto, S.; Meili, R.; Lee, S.; Parry, L.; Firtel, R.A. Spatial and temporal regulation of 3-phosphoinositides by PI 3-kinase and PTEN mediates chemotaxis. Cell 2002, 109, 611–623. [Google Scholar] [CrossRef]
- Wessels, D.; Lusche, D.F.; Kuhl, S.; Heid, P.; Soll, D.R. PTEN plays a role in the suppression of lateral pseudopod formation during Dictyostelium motility and chemotaxis. J. Cell Sci. 2007, 120, 2517–2531. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Tsai, C.-J.; Nussinov, R. Hydrogen bonds and salt bridges across protein-protein interfaces. Protein Eng. 1997, 10, 999–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, C.; Bashton, M.; Kerrison, N.D.; Chothia, C.; Teichmann, S.A. Structure, function and evolution of multidomain proteins. Curr. Opin. Struct. Biol. 2004, 14, 208–216. [Google Scholar] [CrossRef]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A. The Pfam protein families database in 2019. Nucleic Acids Res. 2018, 47, D427–D432. [Google Scholar] [CrossRef]
- Janin, J.; Wodak, S.J. Structural domains in proteins and their role in the dynamics of protein function. Prog. Biophys. Mol. Biol. 1983, 42, 21–78. [Google Scholar] [CrossRef]
- Pawson, T.; Gish, G.D.; Nash, P. SH2 domains, interaction modules and cellular wiring. Trends Cell Biol. 2001, 11, 504–511. [Google Scholar] [CrossRef]
- Mayer, B.J.; Hirai, H.; Sakai, R. Evidence that SH2 domains promote processive phosphorylation by protein-tyrosine kinases. Curr. Biol. 1995, 5, 296–305. [Google Scholar] [CrossRef] [Green Version]
- Park, W.S.; Do Heo, W.; Whalen, J.H.; O’Rourke, N.A.; Bryan, H.M.; Meyer, T.; Teruel, M.N. Comprehensive identification of PIP3-regulated PH domains from C. elegans to H. sapiens by model prediction and live imaging. Mol. Cell 2008, 30, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Weernink, P.A.O.; de Jesús, M.L.; Schmidt, M. Phospholipase D signaling: Orchestration by PIP 2 and small GTPases. Naunyn. Schmiedebergs. Arch. Pharmacol. 2007, 374, 399–411. [Google Scholar] [CrossRef] [PubMed]
- West, M.A.; Bright, N.A.; Robinson, M.S. The role of ADP-ribosylation factor and phospholipase D in adaptor recruitment. J. Cell Biol. 1997, 138, 1239–1254. [Google Scholar] [CrossRef] [PubMed]
- Weber, C.H.; Vincenz, C. The death domain superfamily: A tale of two interfaces? Trends Biochem. Sci. 2001, 26, 475–481. [Google Scholar] [CrossRef]
- Dermody, T.S.; Kirchner, E.; Guglielmi, K.M.; Stehle, T. Immunoglobulin superfamily virus receptors and the evolution of adaptive immunity. PLoS Pathog. 2009, 5, e1000481. [Google Scholar] [CrossRef] [PubMed]
- Trzaskowski, B.; Latek, D.; Yuan, S.; Ghoshdastider, U.; Debinski, A.; Filipek, S. Action of molecular switches in GPCRs-theoretical and experimental studies. Curr. Med. Chem. 2012, 19, 1090–1109. [Google Scholar] [CrossRef]
- Hanoune, J.; Defer, N. Regulation and role of adenylyl cyclase isoforms. Annu. Rev. Pharmacol. Toxicol. 2001, 41, 145–174. [Google Scholar] [CrossRef]
- Kadamur, G.; Ross, E.M. Mammalian phospholipase C. Annu. Rev. Physiol. 2013, 75, 127–154. [Google Scholar] [CrossRef]
- Siehler, S. Regulation of RhoGEF proteins by G12/13-coupled receptors. Br. J. Pharmacol. 2009, 158, 41–49. [Google Scholar] [CrossRef]
- Dascal, N. Signalling via the G protein-activated K+ channels. Cell. Signal. 1997, 9, 551–573. [Google Scholar] [CrossRef]
- Zamponi, G.W.; Currie, K.P.M. Regulation of CaV2 calcium channels by G protein coupled receptors. Biochim. Biophys. Acta (BBA)-Biomembr. 2013, 1828, 1629–1643. [Google Scholar] [CrossRef] [PubMed]
- Lyon, A.M.; Tesmer, J.J.G. Structural insights into phospholipase C-β function. Mol. Pharmacol. 2013, 84, 488–500. [Google Scholar] [CrossRef] [PubMed]
- Ribas, C.; Penela, P.; Murga, C.; Salcedo, A.; García-Hoz, C.; Jurado-Pueyo, M.; Aymerich, I.; Mayor, F., Jr. The G protein-coupled receptor kinase (GRK) interactome: Role of GRKs in GPCR regulation and signaling. Biochim. Biophys. Acta (BBA)-Biomembr. 2007, 1768, 913–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, W.; Luttrell, L.M.; Medvedev, A.V.; Pierce, K.L.; Daniel, K.W.; Dixon, T.M.; Lefkowitz, R.J.; Collins, S. Direct binding of activated c-Src to the β3-adrenergic receptor is required for MAP kinase activation. J. Biol. Chem. 2000, 275, 38131–38134. [Google Scholar] [CrossRef] [PubMed]
- Zenonos, K.; Kyprianou, K. RAS signaling pathways, mutations and their role in colorectal cancer. World J. Gastrointest. Oncol. 2013, 5, 97. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M. Rho signalling at a glance. J. Cell Sci. 2004, 117, 5457–5458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, S.L.; Cao, C.; Pylypenko, O.; Rak, A.; Wandinger-Ness, A. Rab GTPases at a glance. J. Cell Sci. 2007, 120, 3905–3910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, C.L.; Bouvet, S. Arfs at a Glance; The Company of Biologists Ltd.: Cambridge, UK, 2014. [Google Scholar]
- Caputo, E.; Wang, E.; Valentino, A.; Crispi, S.; De Giorgi, V.; Fico, A.; Ficili, B.; Capone, M.; Anniciello, A.; Cavalcanti, E. Ran signaling in melanoma: Implications for the development of alternative therapeutic strategies. Cancer Lett. 2015, 357, 286–296. [Google Scholar] [CrossRef]
- Lemmon, M.A.; Schlessinger, J. Cell signaling by receptor tyrosine kinases. Cell 2010, 141, 1117–1134. [Google Scholar] [CrossRef]
- Depoil, D.; Fleire, S.; Treanor, B.L.; Weber, M.; Harwood, N.E.; Marchbank, K.L.; Tybulewicz, V.L.J.; Batista, F.D. CD19 is essential for B cell activation by promoting B cell receptor–antigen microcluster formation in response to membrane-bound ligand. Nat. Immunol. 2008, 9, 63. [Google Scholar] [CrossRef] [PubMed]
- Tolar, P.; Pierce, S.K. A conformation-induced oligomerization model for B cell receptor microclustering and signaling. In Immunological Synapse; Springer: Berlin, Germany, 2010; pp. 155–169. [Google Scholar]
- Armstrong, N.; Jasti, J.; Beich-Frandsen, M.; Gouaux, E. Measurement of conformational changes accompanying desensitization in an ionotropic glutamate receptor. Cell 2006, 127, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Lu, W.; Milstein, A.D.; Nicoll, R.A. The stoichiometry of AMPA receptors and TARPs varies by neuronal cell type. Neuron 2009, 62, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Ellis-Davies, G.C.R. Caged compounds: photorelease technology for control of cellular chemistry and physiology. Nat. Methods 2007, 4, 619. [Google Scholar] [CrossRef] [PubMed]
- Newman, J.P.; Fong, M.; Millard, D.C.; Whitmire, C.J.; Stanley, G.B.; Potter, S.M. Optogenetic feedback control of neural activity. Elife 2015, 4, e07192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wulff, P.; Arenkiel, B.R. Chemical genetics: Receptor–ligand pairs for rapid manipulation of neuronal activity. Curr. Opin. Neurobiol. 2012, 22, 54–60. [Google Scholar] [CrossRef]
- Levitan, I.B. Modulation of ion channels by protein phosphorylation and dephosphorylation. Annu. Rev. Physiol. 1994, 56, 193–212. [Google Scholar] [CrossRef]
- Zaccolo, M. Phosphodiesterases and compartmentalized cAMP signalling in the heart. Eur. J. Cell Biol. 2006, 85, 693–697. [Google Scholar] [CrossRef]
- Landfield, P.W.; Pitler, T.A. Prolonged Ca2+-dependent afterhyperpolarizations in hippocampal neurons of aged rats. Science 1984, 226, 1089–1092. [Google Scholar] [CrossRef]
- Cuthbertson, K.S.R.; Cobbold, P.H. Phorbol ester and sperm activate mouse oocytes by inducing sustained oscillations in cell Ca2+. Nature 1985, 316, 541. [Google Scholar] [CrossRef]
- Swann, K.; Lai, F.A. The sperm phospholipase C-ζ and Ca2+ signalling at fertilization in mammals. Biochem. Soc. Trans. 2016, 44, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.A.; Fogarty, K.E.; Tsien, R.Y.; Fay, F.S. Calcium gradients in single smooth muscle cells revealed by the digital imaging microscope using Fura-2. Nature 1985, 318, 558. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.W.; Pan, Z.; Kim, E.K.; Lee, J.M.; Bhat, M.B.; Parness, J.; Ma, J. A retrograde signal from calsequestrin for the regulation of store-operated Ca2+ entry in skeletal muscle. J. Biol. Chem. 2003, 278, 3286–3292. [Google Scholar] [CrossRef] [PubMed]
- Brailoiu, E.; Patel, S. Modulation of spontaneous transmitter release from the frog neuromuscular junction by interacting intracellular Ca2+ stores: Critical role for nicotinic acid-adenine dinucleotide phosphate (NAADP). Biochem. J. 2003, 373, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Linse, S.; Helmersson, A.; Forsen, S. Calcium binding to calmodulin and its globular domains. J. Biol. Chem. 1991, 266, 8050–8054. [Google Scholar] [PubMed]
- Wang, H.; Zhang, M. The role of Ca2+-stimulated adenylyl cyclases in bidirectional synaptic plasticity and brain function. Rev. Neurosci. 2012, 23, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Frey, B.M.; Reber, B.F.X.; Vishwanath, B.S.; Escher, G.; Frey, F.J. Annexin I modulates cell functions by controlling intracellular calcium release. FASEB J. 1999, 13, 2235–2245. [Google Scholar] [CrossRef] [PubMed]
- Ciani, L.; Boyle, K.A.; Dickins, E.; Sahores, M.; Anane, D.; Lopes, D.M.; Gibb, A.J.; Salinas, P.C. Wnt7a signaling promotes dendritic spine growth and synaptic strength through Ca2+/Calmodulin-dependent protein kinase II. Proc. Natl. Acad. Sci. USA 2011, 108, 10732–10737. [Google Scholar] [CrossRef]
- Manjarrés, I.M.; Rodríguez-García, A.; Alonso, M.T.; García-Sancho, J. The sarco/endoplasmic reticulum Ca2+ ATPase (SERCA) is the third element in capacitative calcium entry. Cell Calcium 2010, 47, 412–418. [Google Scholar] [CrossRef] [Green Version]
- Yakovlev, A.G.; Wang, G.; Stoica, B.A.; Boulares, H.A.; Spoonde, A.Y.; Yoshihara, K.; Smulson, M.E. A role of the Ca2+/Mg2+-dependent endonuclease in apoptosis and its inhibition by poly (ADP-ribose) polymerase. J. Biol. Chem. 2000, 275, 21302–21308. [Google Scholar] [CrossRef]
- Liu, J.; Farmer, J.D., Jr.; Lane, W.S.; Friedman, J.; Weissman, I.; Schreiber, S.L. Calcineurin is a common target of cyclophilin-cyclosporin A and FKBP-FK506 complexes. Cell 1991, 66, 807–815. [Google Scholar] [CrossRef]
- Brown, M.J.; Palmer, C.R.; Castaigne, A.; De Leeuw, P.W.; Mancia, G.; Rosenthal, T.; Ruilope, L.M. Morbidity and mortality in patients randomised to double-blind treatment with a long-acting calcium-channel blocker or diuretic in the International Nifedipine GITS study: Intervention as a Goal in Hypertension Treatment (INSIGHT). Lancet 2000, 356, 366–372. [Google Scholar] [CrossRef]
- Zhang, S.L.; Yu, Y.; Roos, J.; Kozak, J.A.; Deerinck, T.J.; Ellisman, M.H.; Stauderman, K.A.; Cahalan, M.D. STIM1 is a Ca 2+ sensor that activates CRAC channels and migrates from the Ca 2+ store to the plasma membrane. Nature 2005, 437, 902. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Elce, J.S.; Hamos, J.E.; Nixon, R.A. Widespread activation of calcium-activated neutral proteinase (calpain) in the brain in Alzheimer disease: A potential molecular basis for neuronal degeneration. Proc. Natl. Acad. Sci. USA 1993, 90, 2628–2632. [Google Scholar] [CrossRef] [PubMed]
- Means, A.R.; Dedman, J.R. Calmodulin—an intracellular calcium receptor. Nature 1980, 285, 73. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.H. Calretinin: A gene for a novel calcium-binding protein expressed principally in neurons. J. Cell Biol. 1987, 105, 1343–1353. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.L. Gelsolin: Calcium-and polyphosphoinositide-regulated actin-modulating protein. Bioessays 1987, 7, 176–179. [Google Scholar] [CrossRef] [PubMed]
- Kaftan, E.J.; Ehrlich, B.E.; Watras, J. Inositol 1, 4, 5-trisphosphate (InsP3) and calcium interact to increase the dynamic range of InsP3 receptor-dependent calcium signaling. J. Gen. Physiol. 1997, 110, 529–538. [Google Scholar] [CrossRef]
- Gomez, M.; De Castro, E.; Guarin, E.; Sasakura, H.; Kuhara, A.; Mori, I.; Bartfai, T.; Bargmann, C.I.; Nef, P. Ca2+ signaling via the neuronal calcium sensor-1 regulates associative learning and memory in C. elegans. Neuron 2001, 30, 241–248. [Google Scholar] [CrossRef]
- Vane, J.R.; Mitchell, J.A.; Appleton, I.; Tomlinson, A.; Bishop-Bailey, D.; Croxtall, J.; Willoughby, D.A. Inducible isoforms of cyclooxygenase and nitric-oxide synthase in inflammation. Proc. Natl. Acad. Sci. USA 1994, 91, 2046–2050. [Google Scholar] [CrossRef]
- Leslie, C.C. Properties and regulation of cytosolic phospholipase A2. J. Biol. Chem. 1997, 272, 16709–16712. [Google Scholar] [CrossRef] [PubMed]
- Castagna, M.; Takai, Y.; Kaibuchi, K.; Sano, K.; Kikkawa, U.; Nishizuka, Y. Direct activation of calcium-activated, phospholipid-dependent protein kinase by tumor-promoting phorbol esters. J. Biol. Chem. 1982, 257, 7847–7851. [Google Scholar] [PubMed]
- Oancea, E.; Meyer, T. Protein kinase C as a molecular machine for decoding calcium and diacylglycerol signals. Cell 1998, 95, 307–318. [Google Scholar] [CrossRef]
- Wehrens, X.H.T.; Lehnart, S.E.; Huang, F.; Vest, J.A.; Reiken, S.R.; Mohler, P.J.; Sun, J.; Guatimosim, S.; Song, L.-S.; Rosemblit, N. FKBP12. 6 deficiency and defective calcium release channel (ryanodine receptor) function linked to exercise-induced sudden cardiac death. Cell 2003, 113, 829–840. [Google Scholar] [CrossRef]
- Donato, R. S100: A multigenic family of calcium-modulated proteins of the EF-hand type with intracellular and extracellular functional roles. Int. J. Biochem. Cell Biol. 2001, 33, 637–668. [Google Scholar] [CrossRef]
- Brose, N.; Petrenko, A.G.; Sudhof, T.C.; Jahn, R. Synaptotagmin: A calcium sensor on the synaptic vesicle surface. Science 1992, 256, 1021–1025. [Google Scholar] [CrossRef]
- Carrasco, S.; Mérida, I. Diacylglycerol, when simplicity becomes complex. Trends Biochem. Sci. 2007, 32, 27–36. [Google Scholar] [CrossRef]
- Shin, J.; O’Brien, T.F.; Grayson, J.M.; Zhong, X.-P. Differential regulation of primary and memory CD8 T cell immune responses by diacylglycerol kinases. J. Immunol. 2012, 188, 2111–2117. [Google Scholar] [CrossRef]
- Joshi, R.P.; Schmidt, A.M.; Das, J.; Pytel, D.; Riese, M.J.; Lester, M.; Diehl, J.A.; Behrens, E.M.; Kambayashi, T.; Koretzky, G.A. The ζ isoform of diacylglycerol kinase plays a predominant role in regulatory T cell development and TCR-mediated ras signaling. Sci. Signal. 2013, 6, ra102. [Google Scholar] [CrossRef]
- Nishizuka, Y. The molecular heterogeneity of protein kinase C and its implications for cellular regulation. Nature 1988, 334, 661. [Google Scholar] [CrossRef]
- Huang, K.-P. The mechanism of protein kinase C activation. Trends Neurosci. 1989, 12, 425–432. [Google Scholar] [CrossRef]
- Olivier, M.; Gregory, D.J.; Forget, G. Subversion mechanisms by which Leishmania parasites can escape the host immune response: A signaling point of view. Clin. Microbiol. Rev. 2005, 18, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, M.H.; Berg, H. Interleukin-10 expression is induced by increase of intracellular calcium levels in the monocytic cell line U937. Pflügers Arch. 1998, 435, 868–870. [Google Scholar] [CrossRef] [PubMed]
- Salerno, F.; Paolini, N.A.; Stark, R.; von Lindern, M.; Wolkers, M.C. Distinct PKC-mediated posttranscriptional events set cytokine production kinetics in CD8+ T cells. Proc. Natl. Acad. Sci. USA 2017, 114, 9677–9682. [Google Scholar] [CrossRef] [PubMed]
- Celada, A.; Schreiber, R.D. Role of protein kinase C and intracellular calcium mobilization in the induction of macrophage tumoricidal activity by interferon-gamma. J. Immunol. 1986, 137, 2373–2379. [Google Scholar] [PubMed]
- Brivanlou, A.H.; Darnell, J.E. Signal transduction and the control of gene expression. Science 2002, 295, 813–818. [Google Scholar] [CrossRef] [PubMed]
- Whiteside, S.T.; Goodbourn, S. Signal transduction and nuclear targeting: Regulation of transcription factor activity by subcellular localisation. J. Cell Sci. 1993, 104, 949–955. [Google Scholar] [PubMed]
- Kawai, T.; Adachi, O.; Ogawa, T.; Takeda, K.; Akira, S. Unresponsiveness of MyD88-deficient mice to endotoxin. Immunity 1999, 11, 115–122. [Google Scholar] [CrossRef]
- Kagan, J.C.; Su, T.; Horng, T.; Chow, A.; Akira, S.; Medzhitov, R. TRAM couples endocytosis of Toll-like receptor 4 to the induction of interferon-β. Nat. Immunol. 2008, 9, 361. [Google Scholar] [CrossRef]
- Ross, J.L.; Ali, M.Y.; Warshaw, D.M. Cargo transport: Molecular motors navigate a complex cytoskeleton. Curr. Opin. Cell Biol. 2008, 20, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Hirokawa, N.; Noda, Y.; Tanaka, Y.; Niwa, S. Kinesin superfamily motor proteins and intracellular transport. Nat. Rev. Mol. cell Biol. 2009, 10, 682. [Google Scholar] [CrossRef] [PubMed]
- Klecker, T.; Böckler, S.; Westermann, B. Making connections: interorganelle contacts orchestrate mitochondrial behavior. Trends Cell Biol. 2014, 24, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Wong, Y.C.; Ysselstein, D.; Krainc, D. Mitochondria–lysosome contacts regulate mitochondrial fission via RAB7 GTP hydrolysis. Nature 2018, 554, 382. [Google Scholar] [CrossRef] [PubMed]
- Dagda, R.K.; Banerjee, T. Das Role of protein kinase A in regulating mitochondrial function and neuronal development: Implications to neurodegenerative diseases. Rev. Neurosci. 2015, 26, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.D.; Pawson, T. Cell signaling in space and time: Where proteins come together and when they’re apart. Science 2009, 326, 1220–1224. [Google Scholar] [CrossRef]
- Zhao, C.; Wang, Y.; Chan, K.X.; Marchant, D.B.; Franks, P.J.; Randall, D.; Tee, E.E.; Chen, G.; Ramesh, S.; Phua, S.Y. Evolution of chloroplast retrograde signaling facilitates green plant adaptation to land. Proc. Natl. Acad. Sci. USA 2019, 116, 5015–5020. [Google Scholar] [CrossRef] [Green Version]
- Kleine, T.; Voigt, C.; Leister, D. Plastid signalling to the nucleus: Messengers still lost in the mists? Trends Genet. 2009, 25, 185–192. [Google Scholar] [CrossRef]
- Pfannschmidt, T. Plastidial retrograde signalling–a true “plastid factor” or just metabolite signatures? Trends Plant Sci. 2010, 15, 427–435. [Google Scholar] [CrossRef]
- Koussevitzky, S.; Nott, A.; Mockler, T.C.; Hong, F.; Sachetto-Martins, G.; Surpin, M.; Lim, J.; Mittler, R.; Chory, J. Signals from chloroplasts converge to regulate nuclear gene expression. Science 2007, 316, 715–719. [Google Scholar] [CrossRef]
- Butow, R.A.; Avadhani, N.G. Mitochondrial signaling: The retrograde response. Mol. Cell 2004, 14, 1–15. [Google Scholar] [CrossRef]
- Epstein, C.B.; Waddle, J.A.; Hale IV, W.; Davé, V.; Thornton, J.; Macatee, T.L.; Garner, H.R.; Butow, R.A. Genome-wide responses to mitochondrial dysfunction. Mol. Biol. Cell 2001, 12, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.S.; Small, W.C.; Srere, P.A.; Butow, R.A. Intramitochondrial functions regulate nonmitochondrial citrate synthase (CIT2) expression in Saccharomyces cerevisiae. Mol. Cell. Biol. 1991, 11, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Pracheil, T.; Thornton, J.; Liu, Z. Adenosine triphosphate (ATP) is a candidate signaling molecule in the mitochondria-to-nucleus retrograde response pathway. Genes (Basel) 2013, 4, 86–100. [Google Scholar] [CrossRef] [PubMed]
- Sekito, T.; Thornton, J.; Butow, R.A. Mitochondria-to-nuclear signaling is regulated by the subcellular localization of the transcription factors Rtg1p and Rtg3p. Mol. Biol. Cell 2000, 11, 2103–2115. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.J.; Wang, L.; Ma, Y.; Durick, K.; Perkins, G.; Deerinck, T.J.; Ellisman, M.H.; Taylor, S.S. NH2-terminal targeting motifs direct dual specificity A-kinase–anchoring protein 1 (D-AKAP1) to either mitochondria or endoplasmic reticulum. J. Cell Biol. 1999, 145, 951–959. [Google Scholar] [CrossRef] [PubMed]
- Cardamone, M.D.; Tanasa, B.; Cederquist, C.T.; Huang, J.; Mahdaviani, K.; Li, W.; Rosenfeld, M.G.; Liesa, M.; Perissi, V. Mitochondrial retrograde signaling in mammals is mediated by the transcriptional cofactor GPS2 via direct mitochondria-to-nucleus translocation. Mol. Cell 2018, 69, 757–772. [Google Scholar] [CrossRef] [PubMed]
- Helle, S.C.J.; Kanfer, G.; Kolar, K.; Lang, A.; Michel, A.H.; Kornmann, B. Organization and function of membrane contact sites. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2013, 1833, 2526–2541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kornmann, B.; Currie, E.; Collins, S.R.; Schuldiner, M.; Nunnari, J.; Weissman, J.S.; Walter, P. An ER-mitochondria tethering complex revealed by a synthetic biology screen. Science 2009, 325, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Scarpulla, R.C.; Vega, R.B.; Kelly, D.P. Transcriptional integration of mitochondrial biogenesis. Trends Endocrinol. Metab. 2012, 23, 459–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordens, I.; Fernandez-Borja, M.; Marsman, M.; Dusseljee, S.; Janssen, L.; Calafat, J.; Janssen, H.; Wubbolts, R.; Neefjes, J. The Rab7 effector protein RILP controls lysosomal transport by inducing the recruitment of dynein-dynactin motors. Curr. Biol. 2001, 11, 1680–1685. [Google Scholar] [CrossRef] [Green Version]
- Stoorvogel, W.; Strous, G.J.; Geuze, H.J.; Oorschot, V.; Schwartzt, A.L. Late endosomes derive from early endosomes by maturation. Cell 1991, 65, 417–427. [Google Scholar] [CrossRef]
- Rodriguez-Boulan, E.; Macara, I.G. Organization and execution of the epithelial polarity programme. Nat. Rev. Mol. Cell Biol. 2014, 15, 225. [Google Scholar] [CrossRef] [PubMed]
- Ezratty, E.J.; Partridge, M.A.; Gundersen, G.G. Microtubule-induced focal adhesion disassembly is mediated by dynamin and focal adhesion kinase. Nat. Cell Biol. 2005, 7, 581. [Google Scholar] [CrossRef] [PubMed]
- Paul, N.R.; Jacquemet, G.; Caswell, P.T. Endocytic trafficking of integrins in cell migration. Curr. Biol. 2015, 25, R1092–R1105. [Google Scholar] [CrossRef] [PubMed]
- DeMali, K.A.; Barlow, C.A.; Burridge, K. Recruitment of the Arp2/3 complex to vinculin: Coupling membrane protrusion to matrix adhesion. J. Cell Biol. 2002, 159, 881–891. [Google Scholar] [CrossRef] [PubMed]
- Vidal-Quadras, M.; Holst, M.R.; Francis, M.K.; Larsson, E.; Hachimi, M.; Yau, W.-L.; Peränen, J.; Martín-Belmonte, F.; Lundmark, R. Endocytic turnover of Rab8 controls cell polarization. J. Cell Sci. 2017, 130, 1147–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef] [PubMed]
- Elia, N.; Sougrat, R.; Spurlin, T.A.; Hurley, J.H.; Lippincott-Schwartz, J. Dynamics of endosomal sorting complex required for transport (ESCRT) machinery during cytokinesis and its role in abscission. Proc. Natl. Acad. Sci. USA 2011, 108, 4846–4851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henne, W.M.; Buchkovich, N.J.; Emr, S.D. The ESCRT pathway. Dev. Cell 2011, 21, 77–91. [Google Scholar] [CrossRef] [PubMed]
- Babst, M. MVB vesicle formation: ESCRT-dependent, ESCRT-independent and everything in between. Curr. Opin. Cell Biol. 2011, 23, 452–457. [Google Scholar] [CrossRef] [Green Version]
- Leung, K.F.; Dacks, J.B.; Field, M.C. Evolution of the multivesicular body ESCRT machinery; retention across the eukaryotic lineage. Traffic 2008, 9, 1698–1716. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.; Moita, C.; van Niel, G.; Kowal, J.; Vigneron, J.; Benaroch, P.; Manel, N.; Moita, L.F.; Théry, C.; Raposo, G. Analysis of ESCRT functions in exosome biogenesis, composition and secretion highlights the heterogeneity of extracellular vesicles. J. Cell Sci. 2013, 126, 5553–5565. [Google Scholar] [CrossRef] [PubMed]
- Vivier, E.; Malissen, B. Innate and adaptive immunity: Specificities and signaling hierarchies revisited. Nat. Immunol. 2005, 6, 17. [Google Scholar] [CrossRef] [PubMed]
- Impey, S.; Wayman, G.; Wu, Z.; Storm, D.R. Type I adenylyl cyclase functions as a coincidence detector for control of cyclic AMP response element-mediated transcription: synergistic regulation of transcription by Ca2+ and isoproterenol. Mol. Cell. Biol. 1994, 14, 8272–8281. [Google Scholar] [CrossRef] [PubMed]
- Geetha, N.; Mihaly, J.; Stockenhuber, A.; Blasi, F.; Uhrin, P.; Binder, B.R.; Freissmuth, M.; Breuss, J.M. Signal Integration and Coincidence Detection in the Mitogen-activated Protein Kinase/Extracellular Signal-regulated Kinase (ERK) Cascade: Concomitant Activation of Receptor Tyrosine Kinases and of Lrp-1 Leads to Sustained Erk Phosphorylation Via down-regulation of dual specificity phosphatases (DUSP1 and -6). J. Biol. Chem. 2011, 286, 25663–25674. [Google Scholar] [CrossRef] [PubMed]
- Dumaz, N.; Marais, R. Protein kinase A blocks Raf-1 activity by stimulating 14-3-3 binding and blocking Raf-1 interaction with Ras. J. Biol. Chem. 2003, 278, 29819–29823. [Google Scholar] [CrossRef] [PubMed]
- Peraldi, P.; Frödin, M.; Barnier, J.V.; Calleja, V.; Scimeca, J.-C.; Filloux, C.; Calothy, G.; Van Obberghen, E. Regulation of the MAP kinase cascade in PC12 cells: B-Raf activates MEK-1 (MAP kinase or ERK kinase) and is inhibited by cAMP. FEBS Lett. 1995, 357, 290–296. [Google Scholar] [CrossRef] [Green Version]
- Chiaradonna, F.; Balestrieri, C.; Gaglio, D.; Vanoni, M. RAS and PKA pathways in cancer: New insight from transcriptional analysis. Front. Biosci. 2008, 13, 5257–5278. [Google Scholar] [CrossRef]
- Graves, L.M.; Bornfeldt, K.E.; Raines, E.W.; Potts, B.C.; Macdonald, S.G.; Ross, R.; Krebs, E.G. Protein kinase A antagonizes platelet-derived growth factor-induced signaling by mitogen-activated protein kinase in human arterial smooth muscle cells. Proc. Natl. Acad. Sci. USA 1993, 90, 10300–10304. [Google Scholar] [CrossRef]
- Shin, S.-Y.; Rath, O.; Choo, S.-M.; Fee, F.; McFerran, B.; Kolch, W.; Cho, K.-H. Positive-and negative-feedback regulations coordinate the dynamic behavior of the Ras-Raf-MEK-ERK signal transduction pathway. J. Cell Sci. 2009, 122, 425–435. [Google Scholar] [CrossRef]
- Willardson, B.M.; Wilkins, J.F.; Yoshida, T.; Bitensky, M.W. Regulation of phosducin phosphorylation in retinal rods by Ca2+/calmodulin-dependent adenylyl cyclase. Proc. Natl. Acad. Sci. USA 1996, 93, 1475–1479. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, Y.; Inamitsu, T.; Chida, K.; Iemura, S.-I.; Natsume, T.; Maeda, T.; Hakuno, F.; Takahashi, S.-I. Serine phosphorylation by mTORC1 promotes IRS-1 degradation through SCFβ-TRCP E3 ubiquitin ligase. iScience 2018, 5, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Jordan, J.D.; Iyengar, R. Modes of interactions between signaling pathways. Biochem. Pharmacol. 1998, 55, 1347–1352. [Google Scholar] [PubMed]
- Millar, A.H.; Heazlewood, J.L.; Giglione, C.; Holdsworth, M.J.; Bachmair, A.; Schulze, W.X. The scope, functions, and dynamics of posttranslational protein modifications. Annu. Rev. Plant Biol. 2019, 70. [Google Scholar] [CrossRef] [PubMed]
- Enchev, R.I.; Schulman, B.A.; Peter, M. Protein neddylation: Beyond cullin–RING ligases. Nat. Rev. Mol. Cell Biol. 2015, 16, 30. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.H.; Egli, M.; Stewart, P.L. Structural insights into a circadian oscillator. Science 2008, 322, 697–701. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Pigeonneau, N.; Ventroux, M.; Derouiche, A.; Bidnenko, V.; Mijakovic, I.; Noirot-Gros, M.-F. Protein-tyrosine phosphorylation interaction network in Bacillus subtilis reveals new substrates, kinase activators and kinase cross-talk. Front. Microbiol. 2014, 5, 538. [Google Scholar] [CrossRef]
- Vervoorts, J.; Lüscher-Firzlaff, J.; Lüscher, B. The ins and outs of MYC regulation by posttranslational mechanisms. J. Biol. Chem. 2006, 281, 34725–34729. [Google Scholar] [CrossRef]
- Wong, B.R.; Besser, D.; Kim, N.; Arron, J.R.; Vologodskaia, M.; Hanafusa, H.; Choi, Y. TRANCE, a TNF family member, activates Akt/PKB through a signaling complex involving TRAF6 and c-Src. Mol. Cell 1999, 4, 1041–1049. [Google Scholar] [CrossRef]
- Tamaru, T.; Isojima, Y.; Van Der Horst, G.T.J.; Takei, K.; Nagai, K.; Takamatsu, K. Nucleocytoplasmic shuttling and phosphorylation of BMAL1 are regulated by circadian clock in cultured fibroblasts. Genes Cells 2003, 8, 973–983. [Google Scholar] [CrossRef]
- Lee, C.; Etchegaray, J.-P.; Cagampang, F.R.A.; Loudon, A.S.I.; Reppert, S.M. Posttranslational mechanisms regulate the mammalian circadian clock. Cell 2001, 107, 855–867. [Google Scholar] [CrossRef]
- Weng, G.; Bhalla, U.S.; Iyengar, R. Complexity in biological signaling systems. Science 1999, 284, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Hancock, J.F. Ras nanoclusters: Versatile lipid-based signaling platforms. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2015, 1853, 841–849. [Google Scholar] [CrossRef] [PubMed]
- Whitmarsh, A.J.; Cavanagh, J.; Tournier, C.; Yasuda, J.; Davis, R.J. A mammalian scaffold complex that selectively mediates MAP kinase activation. Science 1998, 281, 1671–1674. [Google Scholar] [CrossRef] [PubMed]
- Leadlay, P.F. Combinatorial approaches to polyketide biosynthesis. Curr. Opin. Chem. Biol. 1997, 1, 162–168. [Google Scholar] [CrossRef]
- Rulifson, E.J.; Wu, C.-H.; Nusse, R. Pathway specificity by the bifunctional receptor frizzled is determined by affinity for wingless. Mol. Cell 2000, 6, 117–126. [Google Scholar] [CrossRef]
- Clandinin, T.R.; DeModena, J.A.; Sternberg, P.W. Inositol trisphosphate mediates a RAS-independent response to LET-23 receptor tyrosine kinase activation in C. elegans. Cell 1998, 92, 523–533. [Google Scholar] [CrossRef]
- Goebeler, M.; Gillitzer, R.; Kilian, K.; Utzel, K.; Bröcker, E.-B.; Rapp, U.R.; Ludwig, S. Multiple signaling pathways regulate NF-κB–dependent transcription of the monocyte chemoattractant protein-1 gene in primary endothelial cells. Blood 2001, 97, 46–55. [Google Scholar] [CrossRef]
- Mullen, A.C.; Orlando, D.A.; Newman, J.J.; Lovén, J.; Kumar, R.M.; Bilodeau, S.; Reddy, J.; Guenther, M.G.; DeKoter, R.P.; Young, R.A. Master transcription factors determine cell-type-specific responses to TGF-β signaling. Cell 2011, 147, 565–576. [Google Scholar] [CrossRef]
- Cruciat, C.-M.; Niehrs, C. Secreted and transmembrane wnt inhibitors and activators. Cold Spring Harb. Perspect. Biol. 2013, 5, a015081. [Google Scholar] [CrossRef]
- Inder, K.; Harding, A.; Plowman, S.J.; Philips, M.R.; Parton, R.G.; Hancock, J.F. Activation of the MAPK module from different spatial locations generates distinct system outputs. Mol. Biol. Cell 2008, 19, 4776–4784. [Google Scholar] [CrossRef] [PubMed]
- Sarma, U.; Sareen, A.; Maiti, M.; Kamat, V.; Sudan, R.; Pahari, S.; Srivastava, N.; Roy, S.; Sinha, S.; Ghosh, I. Modeling and experimental analyses reveals signaling plasticity in a bi-modular assembly of CD40 receptor activated kinases. PLoS ONE 2012, 7, e39898. [Google Scholar] [CrossRef] [PubMed]
- Sassone-Corsi, P. The cyclic AMP pathway. Cold Spring Harb. Perspect. Biol. 2012, 4, a011148. [Google Scholar] [CrossRef] [PubMed]
- Carbonetti, N.H. Immunomodulation in the pathogenesis of Bordetella pertussis infection and disease. Curr. Opin. Pharmacol. 2007, 7, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Cowsert, L.M. In vitro and in vivo activity of antisense inhibitors of ras: Potential for clinical development. Anticancer. Drug Des. 1997, 12, 359–371. [Google Scholar] [PubMed]
- De Bono, J.S.; Rowinsky, E.K. Therapeutics targeting signal transduction for patients with colorectal carcinoma. Br. Med. Bull. 2002, 64, 227–254. [Google Scholar] [CrossRef] [Green Version]
- Dudley, D.T.; Pang, L.; Decker, S.J.; Bridges, A.J.; Saltiel, A.R. A synthetic inhibitor of the mitogen-activated protein kinase cascade. Proc. Natl. Acad. Sci. USA 1995, 92, 7686–7689. [Google Scholar] [CrossRef] [PubMed]
- Sebolt-Leopold, J.S.; Dudley, D.T.; Herrera, R.; Van Becelaere, K.; Wiland, A.; Gowan, R.C.; Tecle, H.; Barrett, S.D.; Bridges, A.; Przybranowski, S. Blockade of the MAP kinase pathway suppresses growth of colon tumors in vivo. Nat. Med. 1999, 5, 810. [Google Scholar] [CrossRef]
- Luo, J.; Manning, B.D.; Cantley, L.C. Targeting the PI3K-Akt pathway in human cancer: Rationale and promise. Cancer Cell 2003, 4, 257–262. [Google Scholar] [CrossRef] [Green Version]
- Geiger, T.; Müller, M.; Dean, N.M.; Fabbro, D. Antitumor activity of a PKC-alpha antisense oligonucleotide in combination with standard chemotherapeutic agents against various human tumors transplanted into nude mice. Anticancer. Drug Des. 1998, 13, 35–45. [Google Scholar]
- Leech, S.H.; Olie, R.A.; Gautschi, O.; Simões-Wüst, A.P.; Tschopp, S.; Häner, R.; Hall, J.; Stahel, R.A.; Zangemeister-Wittke, U. Induction of apoptosis in lung-cancer cells following bcl-xL anti-sense treatment. Int. J. Cancer 2000, 86, 570–576. [Google Scholar] [CrossRef]
- Benjamin, D.; Colombi, M.; Moroni, C.; Hall, M.N. Rapamycin passes the torch: A new generation of mTOR inhibitors. Nat. Rev. Drug Discov. 2011, 10, 868. [Google Scholar] [CrossRef] [PubMed]
- Rodrik-Outmezguine, V.S.; Okaniwa, M.; Yao, Z.; Novotny, C.J.; McWhirter, C.; Banaji, A.; Won, H.; Wong, W.; Berger, M.; de Stanchina, E. Overcoming mTOR resistance mutations with a new-generation mTOR inhibitor. Nature 2016, 534, 272. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Wang, E.; Milazzo, J.P.; Wang, Z.; Kinney, J.B.; Vakoc, C.R. Discovery of cancer drug targets by CRISPR-Cas9 screening of protein domains. Nat. Biotechnol. 2015, 33, 661. [Google Scholar] [CrossRef] [PubMed]
- Rozengurt, E.; Soares, H.P.; Sinnet-Smith, J. Suppression of feedback loops mediated by PI3K/mTOR induces multiple overactivation of compensatory pathways: An unintended consequence leading to drug resistance. Mol. Cancer Ther. 2014, 13, 2477–2488. [Google Scholar] [CrossRef] [PubMed]
- Polakis, P. Wnt signaling and cancer. Genes Dev. 2000, 14, 1837–1851. [Google Scholar] [CrossRef] [PubMed]
- Harikumar, K.B.; Kunnumakkara, A.B.; Ochi, N.; Tong, Z.; Deorukhkar, A.; Sung, B.; Kelland, L.; Jamieson, S.; Sutherland, R.; Raynham, T. A novel small-molecule inhibitor of protein kinase D blocks pancreatic cancer growth in vitro and in vivo. Mol. Cancer Ther. 2010, 9, 1136–1146. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.X.; Sharon, E. IGF-1R as an anti-cancer target—trials and tribulations. Chin. J. Cancer 2013, 32, 242. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Yan, L. Next generation of antibody therapy for cancer. Chin. J. Cancer 2011, 30, 293. [Google Scholar] [CrossRef]
- Hurwitz, H.I.; Fehrenbacher, L.; Hainsworth, J.D.; Heim, W.; Berlin, J.; Holmgren, E.; Hambleton, J.; Novotny, W.F.; Kabbinavar, F. Bevacizumab in combination with fluorouracil and leucovorin: An active regimen for first-line metastatic colorectal cancer. J. Clin. Oncol. 2005, 23, 3502–3508. [Google Scholar] [CrossRef] [PubMed]
- Mason, W.P.; Belanger, K.; Nicholas, G.; Vallières, I.; Mathieu, D.; Kavan, P.; Desjardins, A.; Omuro, A.; Reymond, D. A phase II study of the Ras-MAPK signaling pathway inhibitor TLN-4601 in patients with glioblastoma at first progression. J. Neurooncol. 2012, 107, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Prahallad, A.; Heynen, G.J.J.E.; Germano, G.; Willems, S.M.; Evers, B.; Vecchione, L.; Gambino, V.; Lieftink, C.; Beijersbergen, R.L.; Di Nicolantonio, F. PTPN11 is a central node in intrinsic and acquired resistance to targeted cancer drugs. Cell Rep. 2015, 12, 1978–1985. [Google Scholar] [CrossRef] [PubMed]
- Anantharaman, V.; Iyer, L.M.; Aravind, L. Comparative genomics of protists: New insights into the evolution of eukaryotic signal transduction and gene regulation. Annu. Rev. Microbiol. 2007, 61, 453–475. [Google Scholar] [CrossRef] [PubMed]
- Nichols, S.A.; Dirks, W.; Pearse, J.S.; King, N. Early evolution of animal cell signaling and adhesion genes. Proc. Natl. Acad. Sci. USA 2006, 103, 12451–12456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jambou, R.; Zahraoui, A.; Olofsson, B.; Tavitian, A.; Jaureguiberry, G. Small GTP-binding proteins in Plasmodium falciparum. Biol. Cell 1996, 88, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Al-Khedery, B.; Barnwell, J.W.; Galinski, M.R. Stage-specific expression of 14-3-3 in asexual blood-stage Plasmodium. Mol. Biochem. Parasitol. 1999, 102, 117–130. [Google Scholar] [CrossRef]
- Wurtz, N.; Chapus, C.; Desplans, J.; Parzy, D. cAMP-dependent protein kinase from Plasmodium falciparum: An update. Parasitology 2011, 138, 1–25. [Google Scholar] [CrossRef] [PubMed]
- King, N.; Carroll, S.B. A receptor tyrosine kinase from choanoflagellates: Molecular insights into early animal evolution. Proc. Natl. Acad. Sci. USA 2001, 98, 15032–15037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, K.; Ishida, T. Alu-mediated 100-kb deletion in the primate genome: The loss of the agouti signaling protein gene in the lesser apes. Genome Res. 2006, 16, 485–490. [Google Scholar] [CrossRef] [Green Version]
- Moyle, W.R.; Campbell, R.K.; Myers, R.V.; Bernard, M.P.; Han, Y.; Wang, X. Co-evolution of ligand-receptor pairs. Nature 1994, 368, 251. [Google Scholar] [CrossRef] [PubMed]
- Gerhart, J.; Kirschner, M. The theory of facilitated variation. Proc. Natl. Acad. Sci. USA 2007, 104, 8582–8589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laudet, V. Evolution of the nuclear receptor superfamily: Early diversification from an ancestral orphan receptor. J. Mol. Endocrinol. 1997, 19, 207–226. [Google Scholar] [CrossRef] [PubMed]
- Bridgham, J.T.; Carroll, S.M.; Thornton, J.W. Evolution of hormone-receptor complexity by molecular exploitation. Science 2006, 312, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Noce, T.; Ando, H.; Ueda, T.; Kubokawa, K.; Higashinakagawa, T.; Ishii, S. Molecular cloning and nucleotide sequence analysis of the putative cDNA for the precursor molecule of the chicken LH-β subunit. J. Mol. Endocrinol. 1989, 3, 129–137. [Google Scholar] [CrossRef]
- Prabhu, Y.; Eichinger, L. The Dictyostelium repertoire of seven transmembrane domain receptors. Eur. J. Cell Biol. 2006, 85, 937–946. [Google Scholar] [CrossRef]
- Skorokhod, A.; Gamulin, V.; Gundacker, D.; Kavsan, V.; Muller, I.M.; Muller, W.E.G. Origin of insulin receptor-like tyrosine kinases in marine sponges. Biol. Bull. 1999, 197, 198–206. [Google Scholar] [CrossRef]
- Lam, H.-M.; Chiu, J.; Hsieh, M.-H.; Meisel, L.; Oliveira, I.C.; Shin, M.; Coruzzi, G. Glutamate-receptor genes in plants. Nature 1998, 396, 125. [Google Scholar] [CrossRef]
- Kramer, I.M. Signal Transduction; Academic Press: Cambridge, MA, USA, 2002. [Google Scholar]
- Cadigan, K.M.; Nusse, R. Wnt signaling: A common theme in animal development. Genes Dev. 1997, 11, 3286–3305. [Google Scholar] [CrossRef]
- Hombría, J.C.-G.; Brown, S. The fertile field of Drosophila Jak/STAT signalling. Curr. Biol. 2002, 12, R569–R575. [Google Scholar] [CrossRef]
- Ingham, P.W.; McMahon, A.P. Hedgehog signaling in animal development: Paradigms and principles. Genes Dev. 2001, 15, 3059–3087. [Google Scholar] [CrossRef] [PubMed]
- McKenna, N.J.; O’Malley, B.W. Combinatorial control of gene expression by nuclear receptors and coregulators. Cell 2002, 108, 465–474. [Google Scholar] [CrossRef]
- Moon, R.T.; Bowerman, B.; Boutros, M.; Perrimon, N. The promise and perils of Wnt signaling through β-catenin. Science 2002, 296, 1644–1646. [Google Scholar] [CrossRef] [PubMed]
- Mumm, J.S.; Kopan, R. Notch signaling: From the outside in. Dev. Biol. 2000, 228, 151–165. [Google Scholar] [CrossRef] [PubMed]
- Kopp, E.; Medzhitov, R.; Carothers, J.; Xiao, C.; Douglas, I.; Janeway, C.A.; Ghosh, S. ECSIT is an evolutionarily conserved intermediate in the Toll/IL-1 signal transduction pathway. Genes Dev. 1999, 13, 2059–2071. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Chen, Q.; Ying, S.; Xia, X.; Yu, Z.; Lui, Y.; Tranter, G.; Jin, B.; Song, C.; Seymour, L.W. Evolutionarily conserved primary TNF sequences relate to its primitive functions in cell death induction. J. Cell Sci. 2016, 129, 108–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.-M.; Wang, R.-S.; Chen, L.; Aihara, K. Uncovering signal transduction networks from high-throughput data by integer linear programming. Nucleic Acids Res. 2008, 36, e48. [Google Scholar] [CrossRef] [PubMed]
- Dyjack, N.; Azeredo-Tseng, C.; Yildirim, N. Mathematical modeling reveals differential regulation of MAPK activity by phosphatase proteins in the yeast pheromone response pathway. Mol. Biosyst. 2017, 13, 1323–1335. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Xu, Z.; Ni, M.; Liu, S. Generalized aggregation operators for intuitionistic fuzzy sets. Int. J. Intell. Syst. 2010, 25, 1–30. [Google Scholar] [CrossRef]
- Saez-Rodriguez, J.; Alexopoulos, L.G.; Zhang, M.; Morris, M.K.; Lauffenburger, D.A.; Sorger, P.K. Comparing signaling networks between normal and transformed hepatocytes using discrete logical models. Cancer Res. 2011, 71, 5400–5411. [Google Scholar] [CrossRef] [PubMed]
- MacNamara, A.; Terfve, C.; Henriques, D.; Bernabé, B.P.; Saez-Rodriguez, J. State–time spectrum of signal transduction logic models. Phys. Biol. 2012, 9, 45003. [Google Scholar] [CrossRef] [PubMed]
- Alon, U. Network motifs: Theory and experimental approaches. Nat. Rev. Genet. 2007, 8, 450. [Google Scholar] [CrossRef] [PubMed]
- Brandman, O.; Meyer, T. Feedback loops shape cellular signals in space and time. Science 2008, 322, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, J.D.; de Graauw, M.; van de Water, B.; Geiger, T.; Sharan, R. Elucidation of signaling pathways from large-scale phosphoproteomic data using protein interaction networks. Cell Syst. 2016, 3, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, A. Proteomics of skeletal muscle: Focus on insulin resistance and exercise biology. Proteomes 2016, 4, 6. [Google Scholar] [CrossRef] [PubMed]
- Sharma, I.M.; Petchiappan, A.; Chatterji, D. Quorum sensing and biofilm formation in mycobacteria: Role of c-di-GMP and methods to study this second messenger. IUBMB Life 2014, 66, 823–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sardiu, M.E.; Washburn, M.P. Building protein-protein interaction networks with proteomics and informatics tools. J. Biol. Chem. 2011, 286, 23645–23651. [Google Scholar] [CrossRef]
- Ho, Y.; Gruhler, A.; Heilbut, A.; Bader, G.D.; Moore, L.; Adams, S.-L.; Millar, A.; Taylor, P.; Bennett, K.; Boutilier, K. Systematic identification of protein complexes in Saccharomyces cerevisiae by mass spectrometry. Nature 2002, 415, 180. [Google Scholar] [CrossRef]
- Uetz, P.; Giot, L.; Cagney, G.; Mansfield, T.A.; Judson, R.S.; Knight, J.R.; Lockshon, D.; Narayan, V.; Srinivasan, M.; Pochart, P. A comprehensive analysis of protein–protein interactions in Saccharomyces cerevisiae. Nature 2000, 403, 623. [Google Scholar] [CrossRef]
- Marcotte, E.M.; Pellegrini, M.; Ng, H.-L.; Rice, D.W.; Yeates, T.O.; Eisenberg, D. Detecting protein function and protein-protein interactions from genome sequences. Science 1999, 285, 751–753. [Google Scholar] [CrossRef]
- Aloy, P.; Russell, R.B. Interrogating protein interaction networks through structural biology. Proc. Natl. Acad. Sci. USA 2002, 99, 5896–5901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, R.; Yu, H.; Greenbaum, D.; Kluger, Y.; Krogan, N.J.; Chung, S.; Emili, A.; Snyder, M.; Greenblatt, J.F.; Gerstein, M. A Bayesian networks approach for predicting protein-protein interactions from genomic data. Science 2003, 302, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Penn, B.H.; Netter, Z.; Johnson, J.R.; Von Dollen, J.; Jang, G.M.; Johnson, T.; Ohol, Y.M.; Maher, C.; Bell, S.L.; Geiger, K. An MTB-human protein-protein interaction map identifies a switch between host antiviral and antibacterial responses. Mol. Cell 2018, 71, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Rhee, K.Y.; de Carvalho, L.P.S.; Bryk, R.; Ehrt, S.; Marrero, J.; Park, S.W.; Schnappinger, D.; Venugopal, A.; Nathan, C. Central carbon metabolism in Mycobacterium tuberculosis: An unexpected frontier. Trends Microbiol. 2011, 19, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith-Fischman, S.; Honig, B. Structural genomics: Computational methods for structure analysis. Protein Sci. 2003, 12, 1813–1821. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Fan, K.; Wang, W. The number of protein folds and their distribution over families in nature. Proteins Struct. Funct. Bioinforma. 2004, 54, 491–499. [Google Scholar] [CrossRef]
- Eungdamrong, N.J.; Iyengar, R. Computational approaches for modeling regulatory cellular networks. Trends Cell Biol. 2004, 14, 661–669. [Google Scholar] [CrossRef] [Green Version]
- Kaessmann, H.; Zöllner, S.; Nekrutenko, A.; Li, W.-H. Signatures of domain shuffling in the human genome. Genome Res. 2002, 12, 1642–1650. [Google Scholar] [CrossRef]
- Ueda, M.; Shibata, T. Stochastic signal processing and transduction in chemotactic response of eukaryotic cells. Biophys. J. 2007, 93, 11–20. [Google Scholar] [CrossRef]
- Rangamani, P.; Iyengar, R. Modelling cellular signalling systems. Essays Biochem. 2008, 45, 83–94. [Google Scholar] [CrossRef] [Green Version]
- Arviv, O.; Levy, Y. Folding of multidomain proteins: Biophysical consequences of tethering even in apparently independent folding. Proteins Struct. Funct. Bioinforma. 2012, 80, 2780–2798. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.B.; Manno, M.; Rischel, C.; Thórólfsson, M.; Martorana, V. Aggregation of a multidomain protein: A coagulation mechanism governs aggregation of a model IgG1 antibody under weak thermal stress. Protein Sci. 2010, 19, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Borgia, M.B.; Borgia, A.; Best, R.B.; Steward, A.; Nettels, D.; Wunderlich, B.; Schuler, B.; Clarke, J. Single-molecule fluorescence reveals sequence-specific misfolding in multidomain proteins. Nature 2011, 474, 662. [Google Scholar] [CrossRef] [PubMed]
- Uptain, S.M.; Lindquist, S. Prions as protein-based genetic elements. Annu. Rev. Microbiol. 2002, 56, 703–741. [Google Scholar] [CrossRef] [PubMed]
- Shorter, J.; Lindquist, S. Prions as adaptive conduits of memory and inheritance. Nat. Rev. Genet. 2005, 6, 435. [Google Scholar] [CrossRef] [PubMed]
- Halfmann, R.; Alberti, S.; Lindquist, S. Prions, protein homeostasis, and phenotypic diversity. Trends Cell Biol. 2010, 20, 125–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, K.; Choi, Y.-B.; White-Grindley, E.; Majumdar, A.; Kandel, E.R. Aplysia CPEB can form prion-like multimers in sensory neurons that contribute to long-term facilitation. Cell 2010, 140, 421–435. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; He, X.; Zheng, H.; Huang, L.J.; Hou, F.; Yu, Z.; de la Cruz, M.J.; Borkowski, B.; Zhang, X.; Chen, Z.J. Structural basis for the prion-like MAVS filaments in antiviral innate immunity. Elife 2014, 3, e01489. [Google Scholar] [CrossRef]
- Xu, P.; Rizzoni, E.A.; Sul, S.-Y.; Stephanopoulos, G. Improving metabolic pathway efficiency by statistical model-based multivariate regulatory metabolic engineering. ACS Synth. Biol. 2016, 6, 148–158. [Google Scholar] [CrossRef]
- Kiel, C.; Yus, E.; Serrano, L. Engineering signal transduction pathways. Cell 2010, 140, 33–47. [Google Scholar] [CrossRef]
- Ortlund, E.A.; Bridgham, J.T.; Redinbo, M.R.; Thornton, J.W. Crystal structure of an ancient protein: Evolution by conformational epistasis. Science 2007, 317, 1544–1548. [Google Scholar] [CrossRef] [PubMed]
- Needham, E.J.; Parker, B.L.; Burykin, T.; James, D.E.; Humphrey, S.J. Illuminating the dark phosphoproteome. Sci. Signal. 2019, 12, eaau8645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strickfaden, S.C.; Winters, M.J.; Ben-Ari, G.; Lamson, R.E.; Tyers, M.; Pryciak, P.M. A mechanism for cell-cycle regulation of MAP kinase signaling in a yeast differentiation pathway. Cell 2007, 128, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Lauffenburger, D.A. Cell signaling pathways as control modules: Complexity for simplicity? Proc. Natl. Acad. Sci. USA 2000, 97, 5031–5033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purvis, J.E.; Lahav, G. Encoding and decoding cellular information through signaling dynamics. Cell 2013, 152, 945–956. [Google Scholar] [CrossRef] [PubMed]
- Bower, J.M.; Bolouri, H. Computational Modeling of Genetic and Biochemical Networks; MIT press: Cambridge, MA, USA, 2004. [Google Scholar]
- Wang, B.; Buck, M. Customizing cell signaling using engineered genetic logic circuits. Trends Microbiol. 2012, 20, 376–384. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PROTEIN | CELLULAR/PHYSIOLOGICAL FUNCTION | REFERENCE |
|---|---|---|
| Adenylyl cyclase (AC Type-1) | Act as second messengers in regulatory processes in the central nervous system. | [79] |
| Annexins | Annexin I modulates cell functions by controlling intracellular Ca2+ release. | [80] |
| Ca2+/Calmodulin-dependent protein kinase (CaMK) | Wnt7a signaling promotes dendritic spine growth and synaptic strength through Ca2+/Calmodulin-dependent protein kinase II. | [81] |
| Ca2+-ATPase (SERCA) | The sarco/endoplasmic reticulum Ca2+ ATPase (SERCA) is the third element in capacitative calcium entry. | [82] |
| Ca2+-dependent endonucleases | Ca2+/Mg2+-dependent endonuclease are drivers of apoptosis. | [83] |
| Calcineurin | Calcineurin is a common target of cyclophilin-cyclosporin A and FKBP-FK506 complexes. | [84] |
| Calcium channel blockers | Important roles in arterial and pulmonary hypertension. | [85] |
| Calcium Release-Activated Channel (CRAC) | STIM1 is a Ca2+ sensor that activates CRAC channels and migrates from the Ca2+ store to the plasma membrane. | [86] |
| Calpain | Widespread activation of calcium-activated neutral proteinase (calpain) in the brain in Alzheimer disease. | [87] |
| Calmodulin | Calmodulin—an intracellular calcium receptor. | [88] |
| Calretinin | Calretinin: a gene for a novel calcium-binding protein expressed principally in neurons. | [89] |
| Gelsolin | Gelsolin: calcium-and polyphosphoinositide-regulated actin-modulating protein. | [90] |
| InsP3 receptors | Inositol 1, 4, 5-trisphosphate (InsP3) and calcium interact to increase the dynamic range of InsP3 receptor-dependent calcium signaling. | [91] |
| NCS-1 | Ca2+ signaling via the neuronal calcium sensor-1 regulates associative learning and memory in C. elegans. | [92] |
| Nitric oxide synthase (NOS) | Inducible isoforms of cyclooxygenase and nitric oxide synthase in inflammation. | [93] |
| Phospholipase A2 | cPLA2 requires calcium for its activity. | [94] |
| Phosphorylase kinase | Direct activation of calcium-activated, phospholipid-dependent protein kinase by tumor-promoting phorbol esters. | [95] |
| Protein kinase C | Protein kinase C as a molecular machine for decoding calcium and diacylglycerol signals. | [96] |
| Ryanodine receptors | FKBP12.6 deficiency and defective calcium release channel (ryanodine receptor) function linked to exercise-induced sudden cardiac death. | [97] |
| S100 proteins | S100: A multigenic family of calcium-modulated proteins containing EF hand motif having intracellular and extracellular functional roles. | [98] |
| Synaptotagmin | Synaptotagmin: A calcium sensor on the synaptic vesicle surface. | [99] |
| YEAR | DISCOVERY |
|---|---|
| 1600 | Robert Hooke 1653/Antony Van Leuwenhoek 1682 first observed cell-like structures. |
| 1833 | Anselme Payen isolates first enzyme, diastase. |
| 1839 | Matthias Jakob Schleiden and Theodor Schwann proposed the ‘Cell Theory’. |
| 1876 | Franz Christian Boll discovered Rhodopsin. |
| 1878 | W. F. Kühne proposed the theory of Visual transduction, coined the term “enzyme”. |
| 1884 | J. L. W. Thudichum discovered Sphingolipids (SLs) in the brain. |
| 1889 | G. Yeo coined the term Protoplasm. |
| 1894 | F. A. Locke observed that removal of calcium from frog sartorius-muscle preparation could block the transmission of impulses at the neuromuscular junction. |
| 1895 | N. Cybulski first isolated Adrenaline. |
| 1903 | E. A. Schaefer introduced “Chalones” hormone-like substance. |
| 1905 | J. N. Langley proposed the concept of receptive substance and introduced the “Receptor Theory of Drug Action.” |
| 1905 | Ernest Starling discovered Hormones. |
| 1906 | Camillo Golgi and Santiago Ramón Y Cajal were awarded The Nobel Prize in Physiology or Medicine for their work on the organization of the nervous system. |
| 1921 | Otto Loewi discovered Acetylcholine. |
| 1922 | Frederick Banting and Charles Best discovered Insulin. |
| 1923 | Frederick Grant Banting and John James Rickard Macleod were awarded The Nobel Prize in Physiology or Medicine. |
| 1924 | Hans Spemann and Hilde Mangold identified “Spemann’s Organizer” and The Nobel Prize in Physiology or Medicine in 1935 was awarded to Hans Spemann. |
| 1928 | H. Pollack gave first evidence of Calcium signal. |
| 1929 | Walter Bradford Cannon described ‘Fight’ or ‘Flight’ responses. |
| 1935 | Vittorio Erspamer showed an extract from Enterochromaffin cells made the intestines contract. |
| 1936 | Sir Henry Hallett Dale and Otto Loewi were awarded The Nobel Prize in Physiology or Medicine “for their discoveries relating to chemical transmission of nerve impulses.” |
| 1943 | Takeo Kamada and Haruo Kinoshita showed Ca2+ ions upon injection cause a contraction in muscles. |
| 1947 | Carl Cori and Gerty Cori were awarded The Nobel Prize in Physiology or Medicine “for their discovery of the course of the catalytic conversion of glycogen.” Along with Bernardo Houssay “for his discovery of the part played by the hormone of the anterior pituitary lobe in the metabolism of sugar.” |
| 1948 | Maurice M. Rapport, Arda Green, and Irvine Page co-discovered Serotonin. |
| 1948 | Raymond P. Ahlquist identified subtypes of adrenoreceptors. |
| 1948 | Vittorio Erspamer discovered Octopamine in the salivary glands of the octopus. |
| 1953 | Lowell and Mabel Hokin reported the involvement of Inositol-containing phospholipids in cell regulation. |
| 1953 | Betty M. Twarog and Irvine Page first reported serotonin to be present in the mammalian brain. |
| 1953 | G. H. Sloane Stanley reported Phospholipase C (PLC) in the mammalian brain. |
| 1954 | Takashi Hayashi described a special role of Glutamate in electrophysiological processes. |
| 1955 | Edmond H. Fischer and Edwin G. Krebs discovered the role of Phosphorylation. |
| 1957 | Lord Todd was awarded The Nobel Prize in Chemistry “for his work on nucleotides and nucleotide co-enzymes.” |
| 1957 | T. W. Rall and coworkers discovered cAMP. |
| 1958 | Frederick Sanger was awarded The Nobel Prize in Chemistry “for his work on the structure of proteins, especially that of Insulin.” |
| 1958 | George Beadle and Edward Tatum were awarded The Nobel Prize in Physiology or Medicine “for their discovery that genes act by regulating definite chemical events.” |
| 1961 | E. Essner and Alex B. Novikoff discovered Acid phosphatase inside lysosome using electron microscopy. |
| 1961 | Georg von Békésy was awarded The Nobel Prize in Physiology or Medicine “for his discoveries of the physical mechanism of stimulation within the cochlea.” |
| 1962 | Max F. Perutz and John C. Kendrew were awarded The Nobel Prize in Chemistry “for their studies of the structures of globular proteins.” |
| 1962 | R. W. Butcher and E. W Sutherland discovered Phosphodiesterase (PDE) enzyme that removes cAMP. |
| 1963 | Sir John C. Eccles, Alan L. Hodgkin, and Andrew F. Huxley were awarded The Nobel Prize in Physiology or Medicine “for their discoveries concerning the ionic mechanisms involved in excitation and inhibition in the peripheral and central portions of the nerve cell membrane.” |
| 1963 | Z. A. Cohn concluded lysosomes act as cells digestive system to recycle compounds. |
| 1964 | Jennifer Harvey discovered monomeric G-Protein/GTPase in rat sarcoma (Harvey-Ras). |
| 1964 | Richard A. Lockshin and Carroll M. Williams reported on Programmed Cell Death “Endocrine potentiation of the breakdown of the intersegmental muscles of silkmoths.” |
| 1965 | G. Heppner and D. W. Weiss Discovered TLR4 as a receptor for LPS. |
| 1965 | François Jacob, André Lwoff, and Jacques Monod were awarded The Nobel Prize in Physiology or Medicine “for their discoveries concerning genetic control of enzyme and virus synthesis.” |
| 1967 | Ragnar Granit, Haldan K. Hartline, and George Wald were awarded The Nobel Prize in Physiology or Medicine “for their discoveries concerning the primary physiological and chemical visual processes in the eye”. |
| 1967 | W. H. Kirsten and L. A. Mayer discovered Kirsten Murine Sarcoma Virus. |
| 1968 | S. Ebashi and M. Endo discovered Troponin. |
| 1968 | D. A. Walsh et al., showed that cAMP controlled the activity of PKA. |
| 1970 | Jacques Benveniste et al., discovered Platelet-activating factor. |
| 1970 | Luis F. Leloir was awarded The Nobel Prize in Chemistry “for his discovery of sugar nucleotides and their role in the biosynthesis of carbohydrates.” |
| 1970 | Sir Bernard Katz, Ulf von Euler, and Julius Axelrod were awarded The Nobel Prize in Physiology or Medicine “for their discoveries concerning the humoral transmitters in the nerve terminals and the mechanism for their storage, release, and inactivation.” |
| 1971 | J. R. Vane; G. J. Roth et al., unveiled the mechanism of action of Aspirin. |
| 1971 | R. Miledi et al., isolated cholinergic receptor protein of torpedo electric tissue. |
| 1971 | Rodbell et al., showed the obligatory role of guanylnucleotides in Glucagon’s action. |
| 1971 | Earl W. Sutherland was awarded The Nobel Prize in Physiology or Medicine “for his discoveries concerning the mechanisms of the action of hormones.” |
| 1972 | J. F. R. Kerr et al., coined the term ‘Apoptosis.’ |
| 1974 | Russel Ross et al., discovered that factors extracted from platelets could induce quiescent smooth muscle cells to synthesize DNA. |
| 1974 | A. Tissieres et al., demonstrated that temperature stress induces expression of Heat Shock Proteins. |
| 1975 | P. A. Lawrence and P. M. Shelton reported polarity in the developing insect retina. |
| 1975 | R. H. Michell showed that receptor-activated hydrolysis of PIP2 produced a molecule that caused an increase in intracellular calcium mobilization. |
| 1976 | J. F. Borel et al., showed immunosuppressive properties of macrolide Cyclosporine-A. |
| 1977 | Roger Guillemin and Andrew V. Schally were awarded The Nobel Prize in Physiology or Medicine “for their discoveries concerning the peptide hormone production of the brain.” Along with Rosalyn Yalow “for the development of radioimmunoassays of peptide hormones.” |
| 1977 | Y. Takai et al., confirmed the presence of Cyclic-nucleotide independent protein kinase in bovine cerebellum. |
| 1979 | Discovery of p53 signaling. |
| 1979 | H. N. Antoniades et al., purified Platelet Derived Growth Factor (PDGF). |
| 1979 | R. A Weinberg et al., showed that DNA isolated from chemically transformed rodent fibroblasts caused the morphologic transformation of mouse fibroblasts. |
| 1979 | Philip Cohen published seminal studies on protein phosphorylation, Phosphorylase kinase and role of phosphatases PPPLCA and GSK3β in the regulation of glycogen metabolism. |
| 1980 | Erikson et al.; Hunter and Sefton, isolated Protein tyrosine kinase v-Src. |
| 1980 | A. Hershko and A. Varshavsky’s laboratories independently elucidated detailed signaling associated with ubiquitin system; degradation signals (degrons) in short-lived proteins; and in vivo controls of protein fluxes. |
| 1980 | P. J. Novick et al., elucidated intracellular transport pathways in Yeast. |
| 1980 | Wieschaus and Nusslein-Volhard identified embryonic lethal, loss-of-function alleles of Wingless (Wg). |
| 1981 | J. E. Smart et al., discovered viral oncogene Src constituted a mutated tyrosine protein kinase. |
| 1981 | A. Roberts and M. Sporn discovered TGF-β. |
| 1981 | R. H. Michell et al., showed IP3 acts as a secondary messenger capable of traversing through the cytoplasm to the ER, thereby stimulating the release of Ca2+ into the cytoplasm. |
| 1981 | David H. Hubel and Torsten N. Wiesel were awarded The Nobel Prize in Physiology or Medicine “for their discoveries concerning information processing in the visual system.” |
| 1982 | J. A. Cooper et al., reported dimerization of PDGFR. |
| 1982 | P. Walter and G. Blobel identified Signal Recognition Particle (SRP) and later contributed to the understanding of Unfolded Protein Response (UPR). |
| 1982 | Sune K. Bergström, Bengt I. Samuelsson and John R. Vane were awarded The Nobel Prize in Physiology or Medicine 1982 “for their discoveries concerning prostaglandins and related biologically active substances.” |
| 1982 | Y. Barde et al., purified Brain-Derived Neurotrophic Factor (BDNF). |
| 1983 | K. Shimizu et al., discovered neuroblastoma cell line transforming gene product N-Ras. |
| 1983 | Sir J. Black and P. Leff used β blockers for the treatment of Angina pectoris. |
| 1984 | R. A. Cerione et al., demonstrated functional coupling between guinea pig-derived β-2-Adrenergic receptor and human guanine nucleotide-binding regulatory protein (NS) of adenylyl cyclase was sufficient to recapitulate a neurotransmitter responsive system. |
| 1985 | A. N. Hollenberg et al., 1985; S. Green et al., 1986 discovered nuclear superfamily of receptors: Human glucocorticoid and Oestrogen receptor. |
| 1985 | M. S. Brown and J. L. Goldstein were awarded The Nobel Prize in Chemistry “for their discoveries concerning the regulation of cholesterol metabolism.” |
| 1985 | G. Grynkiewicz et al., discovered Ca2+ indicators with greatly improved fluorescence properties. |
| 1985 | Richard O Hynes and J. W. Tamkun co-discovered Integrins. |
| 1985 | Lewis C Cantley and co-workers discovered Phosphoinositide kinase (PI3K). |
| 1986 | P. Russell and P. Nurse discovered Dual specificity phosphatases. |
| 1986 | Stanley Cohen and Rita Levi-Montalcini were awarded The Nobel Prize in Physiology or Medicine “for their discoveries of growth factors.” |
| 1986 | R. Weinberg elaborated the role of Ras, the first human cancer-causing gene and growth suppressor Retinoblastoma (Rb) protein. |
| 1986 | Ranjan Sen and David Baltimore discovered NF-ĸB. |
| 1986 | Tony Pawson and colleagues discovered the SH2 domain and its role in cellular transformation. |
| 1986 | U. Wilden and co-workers discovered arrestin. |
| 1986 | Y. Nishizuka et al., revealed that calcium released by IP3 work with DAG to activate protein kinase C. |
| 1987 | A. G. Gilman found that GTP cofactor acted through binding of “Transducer”, the effector protein that connects receptor and effector. |
| 1987 | S. J. Elledge and R. W. Davis discovered that ribonucleotide reductase (RNRs) are turned on by DNA damage and are regulated by the cell cycle. |
| 1987 | S. P. Staal cloned Akt oncogene and its human homologs AKT1 and AKT2. |
| 1987 | T. Imagawa et al., purified Ryanodine receptor from skeletal muscle sarcoplasmic reticulum. |
| 1987 | W. Lee et al., discovered AP-1 as a TPA-activated transcription factor that drives the expression of Metallothionein genes. |
| 1988 | R. A. Dixon et al., cloned first mammalian - β adrenergic receptor. |
| 1988 | Sir James W. Black, Gertrude B. Elion, and George H. Hitchings were awarded The Nobel Prize in Physiology or Medicine “for their discoveries of important principles for drug treatment.” |
| 1988 | K. J. Kemphues et al., discovered PAR proteins in C. elegans responsible for animal cell polarization. |
| 1988 | S. R. Sprang elucidated the mechanism of Glycogen Phosphorylase and kinase activation. |
| 1989 | E. Pfeuffer et al., discovered Olfactory Adenylyl Cyclase. |
| 1989 | S. G. Rhee et al., found that phospholipase C (PLC) is the phosphodiesterase responsible for hydrolyzing PIP2 into DAG and IP3. |
| 1989 | B. Vogelstein and co-workers discovered TP53 to be the most frequently mutated in most human cancers. |
| 1989 | C. A. Finlay et al., showed that p53 proto-oncogene acts as a suppressor of transformation. |
| 1989 | Napoleone Ferrara discovered Vascular Endothelial Growth Factor (VEGF). |
| 1989 | P. Gardner showed the role of store-operated Ca2+ entry (SOCE), through STIM1 and ORAI1 in the understanding of immune cell activation (Clonal activation, and Tolerance). |
| 1989 | Sidney Altman and Thomas R. Cech were awarded The Nobel Prize in Chemistry 1989 “for their discovery of catalytic properties of RNA.” |
| 1990 | S. D. Wright et al., found that LPS sensing occurs through TLR4 and its coreceptor CD14. |
| 1991 | A. Galione et al., 1991; P. C. Lee et al., 1993 found that adenine nucleotides induced Ca2+ mobilization in the sea urchin. |
| 1991 | Erwin Neher and Bert Sakmann were awarded The Nobel Prize in Physiology or Medicine “for their discoveries concerning the function of single ion channels in cells.” |
| 1991 | B. Vogelstein and K. W. Kinzler discovered another tumor suppressor gene associated with familial adenomatous polyposis (FAP) and known as Adenomatous polyposis coli. |
| 1991 | C. I. Bargmann and H. R. Horvitz identified chemosensory neurons that are important in the olfactory sense in nematode C. elegans. |
| 1991 | F. Mckeon discovered immunosuppressant Tacrolimus/FK506 was a potent inhibitor of FKBP1A and inhibits calcineurin signaling by hindering substrate access. |
| 1991 | Hans Cleaver and co-workers reported the cloning of a T cell-specific transcription factor that they termed TCF1 and elaborated Wnt signaling. |
| 1991 | M. N. Hall and co-workers discovered the target of rapamycin (TOR) and its role in cell growth control in Saccharomyces cerevisiae. |
| 1991 | Richard R. Ernst was awarded The Nobel Prize in Chemistry “for his contributions to the development of the methodology of high resolution nuclear magnetic resonance (NMR) spectroscopy.” |
| 1992 | Edmond H. Fischer and Edwin G. Krebs were awarded The Nobel Prize in Physiology or Medicine “for their discoveries concerning reversible protein phosphorylation as a biological regulatory mechanism.” |
| 1992 | S. Nakanishi et al., 1992; G. Powis et al., 1994, identified A microbial product Wortmannin as an inhibitor of myosin light chain kinase and later as an inhibitor of PI3K. |
| 1992 | Yoshinori Ohsumi and coworkers discovered that autophagy also occurs in Yeast. |
| 1993 | J. P. Oliver et al., found that Drosophila SH2-SH3 adaptor protein is involved in coupling the sevenless tyrosine kinase to an activator of Ras guanine nucleotide exchange, Sos. |
| 1993 | Kazutoshi Mori identified a cellular quality-control system unfolded proteins response (UPR) and identified IRE1 as a core component of the UPR in Yeast. |
| 1993 | N. Marchenko et al., demonstrated a direct role of p53 in mitochondrial apoptosis. |
| 1994 | Alfred G. Gilman and Martin Rodbell were awarded The Nobel Prize in Physiology or Medicine “for their discovery of G-proteins and the role of these proteins in signal transduction in cells.” |
| 1994 | M. Rothe et al., identified TRAF-2 as Signal transducer associated with the cytoplasmic domain of the 75kDa tumor necrosis factor receptor 2. |
| 1997 | Paul D. Boyer, John E. Walker, and Jens C. Skou were jointly awarded The Nobel Prize in Chemistry “for their elucidation of the enzymatic mechanism underlying the synthesis of adenosine triphosphate (ATP).” |
| 1997 | Three groups independently discovered the SOCS1 protein: T. A. Endo et al., as a JAK-binding protein (JAB) as a suppressor of IL-6 signaling, R. Starr et al., based on sequence homology with the STAT3-SH2 domain and T. Naka et al., STAT-induced STAT Inhibitor (SSI). |
| 1998 | Robert F. Furchgott, Louis J. Ignarro and Ferid Murad were awarded The Nobel Prize in Physiology or Medicine “for their discoveries concerning nitric oxide as a signaling molecule in the cardiovascular system.” |
| 1999 | Günter Blobel was awarded The Nobel Prize in Physiology or Medicine “for the discovery that proteins have intrinsic signals that govern their transport and localization in the cell.” |
| 2000 | Arvid Carlsson, Paul Greengard, and Eric R. Kandel were awarded The Nobel Prize in Physiology or Medicine “for their discoveries concerning signal transduction in the nervous system.” |
| 2001 | Leland H. Hartwell, Tim Hunt, and Sir Paul M. Nurse were awarded The Nobel Prize in Physiology or Medicine “for their discoveries of key regulators of the cell cycle.” |
| 2002 | J. Takagi et al., investigated how integrin binding is conveyed to the cell interior. |
| 2002 | K. Hamada et al., 3D structures of the inositol 1,4,5-triphosphate (IP3) were elucidated. |
| 2002 | P. Lassus et al., the apoptosome may act as an amplifier rather than an initiator of caspase activation. |
| 2003 | Peter Agre and Roderick MacKinnon were jointly awarded The Nobel Prize in Chemistry “for the discovery of water channels” and “for structural and mechanistic studies of ion channels” respectively. |
| 2003 | B. D. Manning et al., Identification of tumor suppressor gene product tuberin (tuberous sclerosis complex-2) as a target of the phosphoinositide 3-kinase/Akt pathway. |
| 2003 | M. Yaffe and Coworkers identified BRCT repeats as phosphopeptide-binding modules involved in protein targeting. |
| 2004 | G. Di Paolo et al., provided genetic evidence for a critical role of PI(4,5)P2 synthesis in the physiology of neurotransmission. |
| 2004 | I. Tassiulas et al., Inflammatory responses in macrophages by Syk and ITAM-containing adaptors were reported. |
| 2004 | Richard Axel and Linda B. Buck were awarded The Nobel Prize in Physiology or Medicine “for their discoveries of odorant receptors and the organization of the olfactory system.” |
| 2004 | Aaron Ciechanover, Avram Hershko and Irwin Rose were awarded The Nobel Prize in Chemistry “for the discovery of ubiquitin-mediated protein degradation.” |
| 2005 | Edward S. Boyden accredited with discovering the Optogenetics. |
| 2005 | R. B. Seth et al., Identification and characterization of MAVS, a mitochondrial antiviral signaling protein that activates NF-κB and IRF-3. |
| 2005 | J. Liou et al., identified STIM as a Ca2+ sensor essential for Ca2+-store-depletion-triggered Ca2+ influx. |
| 2005 | J. Ptacek et al., Systems-level macromolecular networks in Yeast were identified by Globalanalysis of protein phosphorylation. |
| 2006 | Roger D. Kornberg was awarded The Nobel Prize in Chemistry “for his studies of the molecular basis of eukaryotic transcription.” |
| 2006 | S. Takamori et al., presented Molecular anatomy of a trafficking organelle. |
| 2007 | M. J. Rust et al., presented the role of ordered phosphorylation in the oscillation of a three-protein circadian clock in Cyanobacteria. |
| 2007 | S. Y. Zhang TLR3 deficiency was found to be associated with susceptibility to herpes simplex encephalitis. |
| 2007 | J. Bilic et al., LRP6-signalosome” is depended on scaffolding protein Dishevelled. |
| 2008 | D. Pincus et al., discussed the evolution of phospho-tyrosine signaling machinery in Premetazoan lineages. |
| 2008 | B. Apsel et al., accredited with the discovery of dual inhibitors of tyrosine and phosphoinositide kinases. |
| 2008 | Osamu Shimomura, Martin Chalfie and Roger Y. Tsien were awarded The Nobel Prize in Chemistry “for the discovery and development of the green fluorescent protein, GFP.” |
| 2009 | E. Meylan et al., reported connections of NF-κB and oncogenic K-Ras in lung tumor development. |
| 2009 | W. L. Yang et al., Membrane recruitment and phosphorylation of ubiquitinated AKT are promoted by TRAF6. |
| 2010 | A. Breitkreutz et al., deciphered A Global Protein Kinase and phosphatase interaction network in Yeast. |
| 2010 | S. J. Heidorn showed that kinase-dead BRAF cooperates with Ras to hyperactivated CRAF leading to enhancement of MEK and ERK signaling. |
| 2010 | J. Oh et al., showed an association of mTORC2 with ribosomes and phosphorylates the nascent Akt peptide. |
| 2011 | B. M. Gardner and P. Walter described that the unfolded proteins themselves bind directly to Ire1 to stimulate their oligomerization and activation. |
| 2011 | B. Gerlach et al., Linear ubiquitination prevents inflammation and regulates immune signaling. |
| 2011 | E. A. Kiss et al., Natural aryl hydrocarbon receptor ligands control organogenesis of intestinal lymphoid follicles. |
| 2012 | Eric Kandel and group discovered piRNAs as epigenetic controllers of memory-related synaptic plasticity. |
| 2012 | T. R. Wilson et al., highlighted the widespread potential for growth-factor-driven resistance to anticancer kinase inhibitors. |
| 2012 | Robert J. Lefkowitz and Brian K. Kobilka were awarded The Nobel Prize in Chemistry “for studies of G-protein-coupled receptors.” |
| 2012 | Sir John B. Gurdon and Shinya Yamanaka were awarded The Nobel Prize in Physiology or Medicine “for the discovery that mature cells can be reprogrammed to become pluripotent.” |
| 2013 | James E. Rothman, Randy W. Schekman and Thomas C. Südhof were awarded The Nobel Prize in Physiology or Medicine “for their discoveries of machinery regulating vesicle traffic, a major transport system in our cells.” |
| 2013 | James Chen and colleagues discovered another second messenger cGAMP and its activation through Cyclic GMP-AMP synthase, a cytosolic DNA sensor that activates the Type-I interferon pathway and confers antiviral immunity. |
| 2013 | J. E. Toettcher et al., employed optogenetics to interrogate the dynamic control of signaling transmission by the Ras/Erk module. |
| 2013 | P. Vizán et al., revealed Receptor dynamics determine attenuation and refractory behavior of the TGF-β pathway. |
| 2014 | J. C. H. Tam et al., highlighted Intracellular sensing of complement C3 activates cell-autonomous immunity. |
| 2014 | M. AlQuraishi et al., used a computational approach to establish a statistical mechanical framework that integrates biophysical and genomic data to assemble cancer networks. |
| 2015 | Tomas Lindahl, Paul Modrich and Aziz Sancar were awarded The Nobel Prize in Chemistry 2015 “for mechanistic studies of DNA repair.” |
| 2015 | R. G. Efremov deciphered architecture and conformational switch mechanism of the ryanodine receptor. |
| 2015 | M. Lazarou et al., highlighted the role of the ubiquitin kinase PINK1 in autophagy receptors to induce mitophagy. |
| 2016 | John Sondek designed a peptide based on Helix-Turn-Helix (HTH) that selectively blocks an important class of Gαq proteins and prevents interaction with its downstream signaling partners. |
| 2016 | Nicolas Doucet and his research team found that activated RTKs terminate downstream signaling via the direct phosphorylation of an evolutionarily conserved Tyr present in most SRC homology (SH) 3 domains. |
| 2017 | Barry V. L. Potter and Andreas H. Guse and their co-workers found that 2′-deoxy-ADPR (dADPR) an endogenous TRPM2 superagonist may act as a cell-signaling molecule. |
| 2017 | R. Ravindran et al., discovered that the amino acid sensor GCN2 controls gut inflammation by inhibiting inflammasome activation. |
| 2017 | R. A. Saxton et al., showed the structural basis for leucine sensing by the Sestrin2-mTORC1 pathway. |
| 2018 | Frances H. Arnold and George P. Smith were awarded The Nobel Prize in Chemistry “for the directed evolution of enzymes” and “for the phage display of peptides and antibodies” respectively. |
| 2018 | Nicolas Doucet identified a discovery of a new molecular switch that controls activated receptor tyrosine kinases (RTKs) through SRC homology (SH) 3 domains. |
| 2018 | John Sondek and Team developed small proteins called peptides that selectively block a certain type of G-protein signaling. |
| 2019 | Jay T. Groves and his team showed that a molecular assembly phase transition and kinetic proofreading modulate Ras activation by Sos. |
| 2019 | Kaisa Lehti et al., found that FGFR4 efficiently phosphorylates several essential proteins of the Hippo tumor suppressor pathway. |
| MAJOR APPROACHES | TECHNIQUES |
|---|---|
| Imaging |
|
| Centrifugation |
|
| Flow cytometry |
|
| Chromatography |
|
| Spectroscopy |
|
| Algorithms |
|
| Transcriptomics and metabolomics characterization |
|
| Protein-protein interactions Protein-nucleic acid interactions |
|
| In vitro and in vivo real-time analysis |
|
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nair, A.; Chauhan, P.; Saha, B.; Kubatzky, K.F. Conceptual Evolution of Cell Signaling. Int. J. Mol. Sci. 2019, 20, 3292. https://doi.org/10.3390/ijms20133292
Nair A, Chauhan P, Saha B, Kubatzky KF. Conceptual Evolution of Cell Signaling. International Journal of Molecular Sciences. 2019; 20(13):3292. https://doi.org/10.3390/ijms20133292
Chicago/Turabian StyleNair, Arathi, Prashant Chauhan, Bhaskar Saha, and Katharina F. Kubatzky. 2019. "Conceptual Evolution of Cell Signaling" International Journal of Molecular Sciences 20, no. 13: 3292. https://doi.org/10.3390/ijms20133292
APA StyleNair, A., Chauhan, P., Saha, B., & Kubatzky, K. F. (2019). Conceptual Evolution of Cell Signaling. International Journal of Molecular Sciences, 20(13), 3292. https://doi.org/10.3390/ijms20133292