Interleukin-10 Facilitates Glutamatergic Synaptic Transmission and Homeostatic Plasticity in Cultured Hippocampal Neurons



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
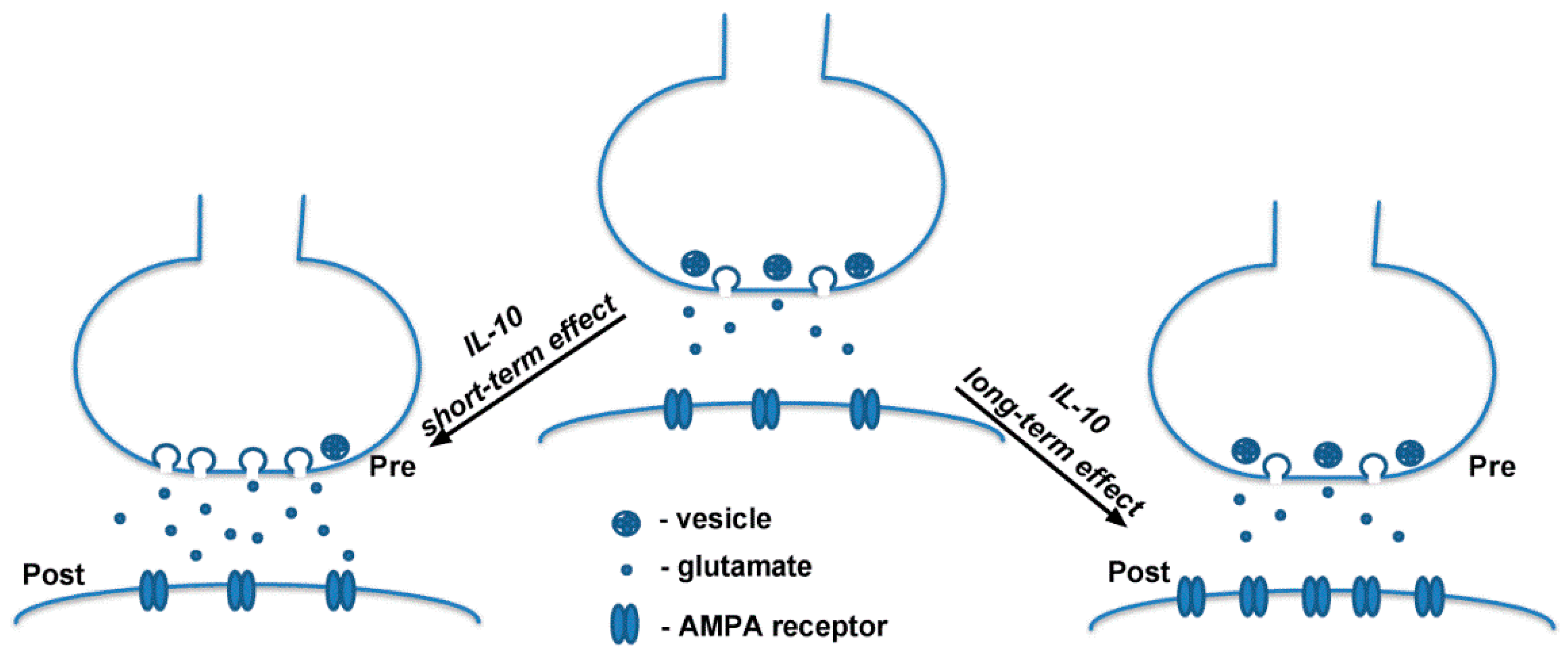
:1. Introduction
2. Results
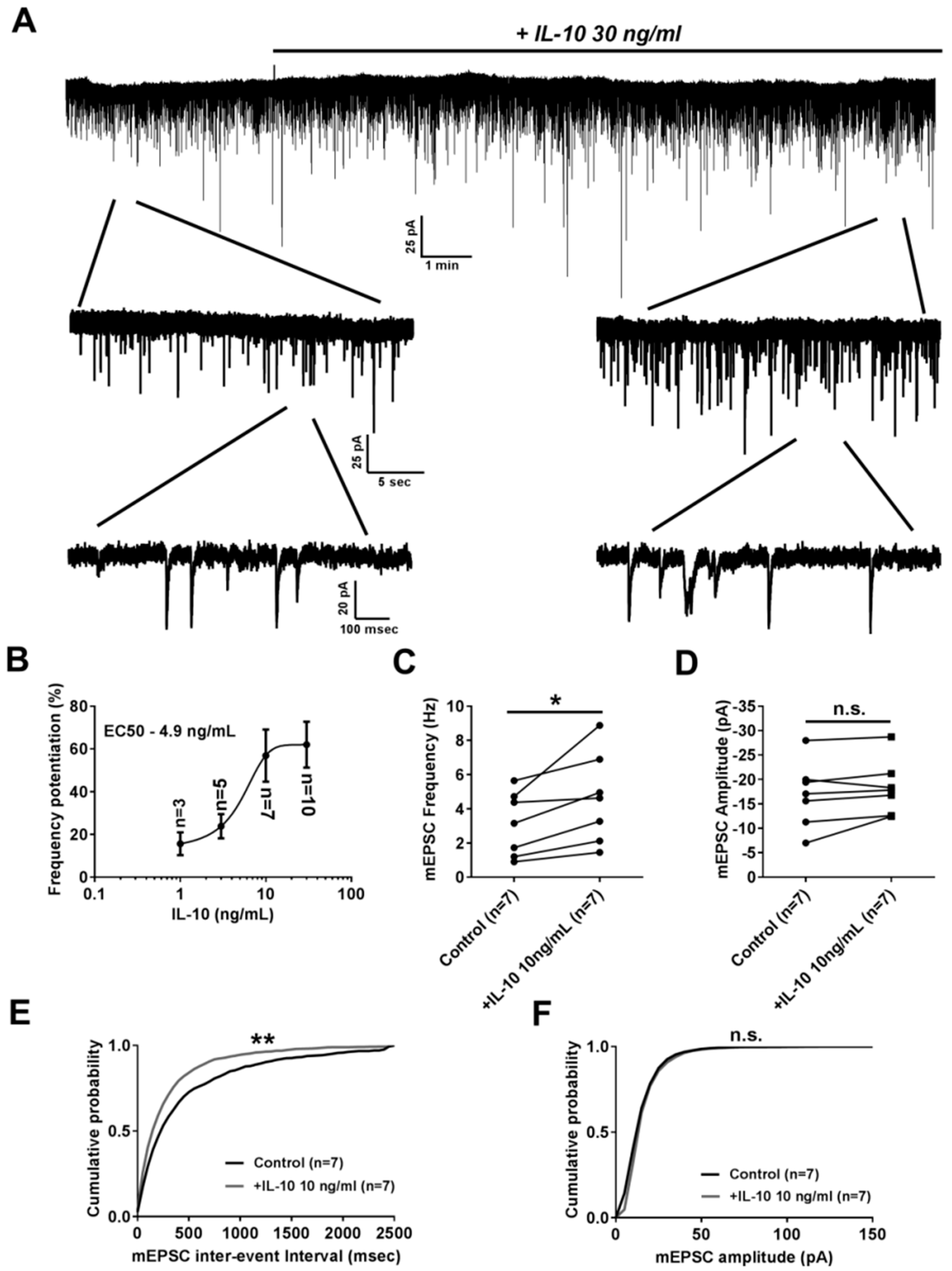
2.1. Effect of Interleukin-10 on Basal Glutamatergic Synaptic Transmission
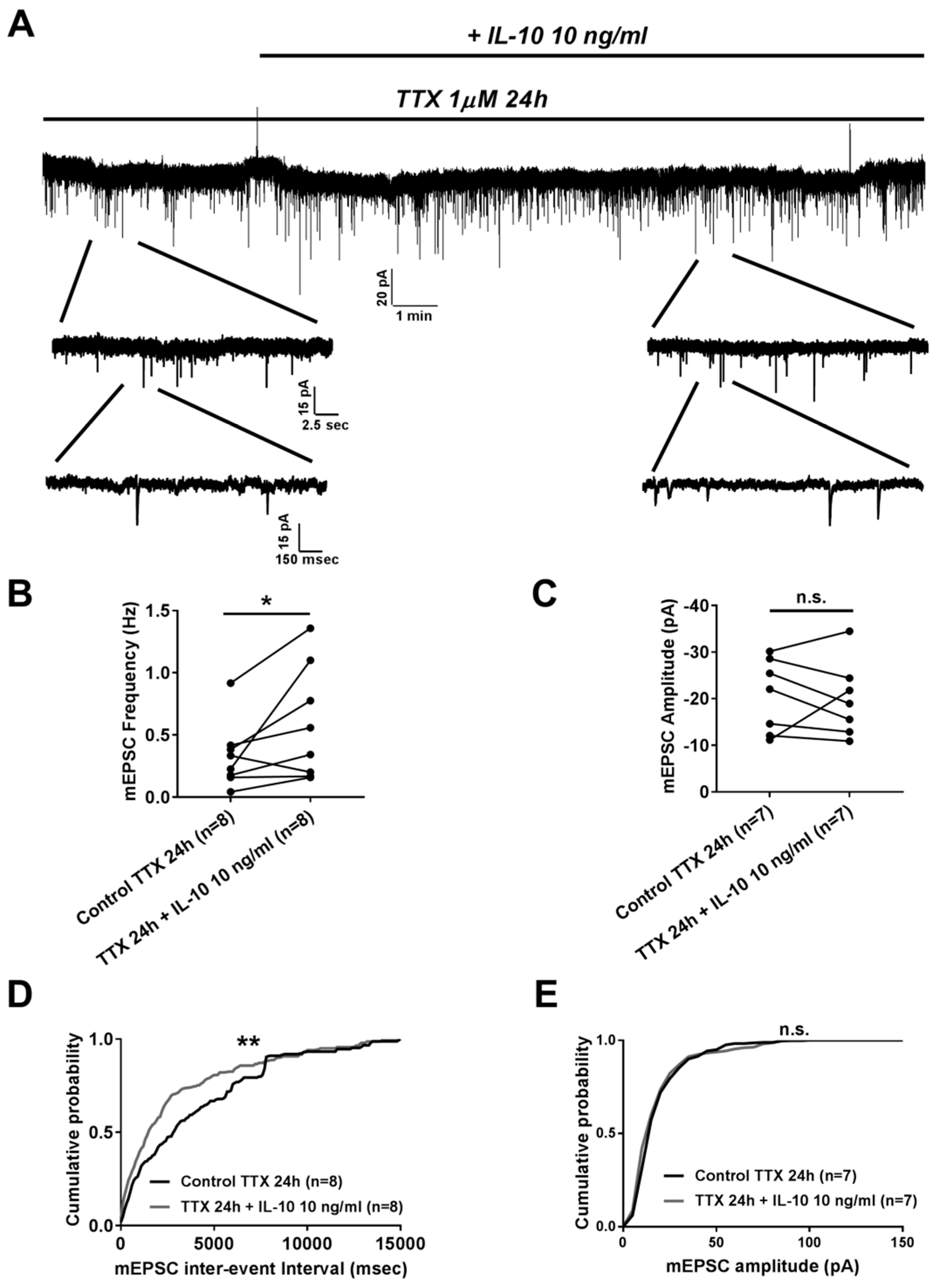
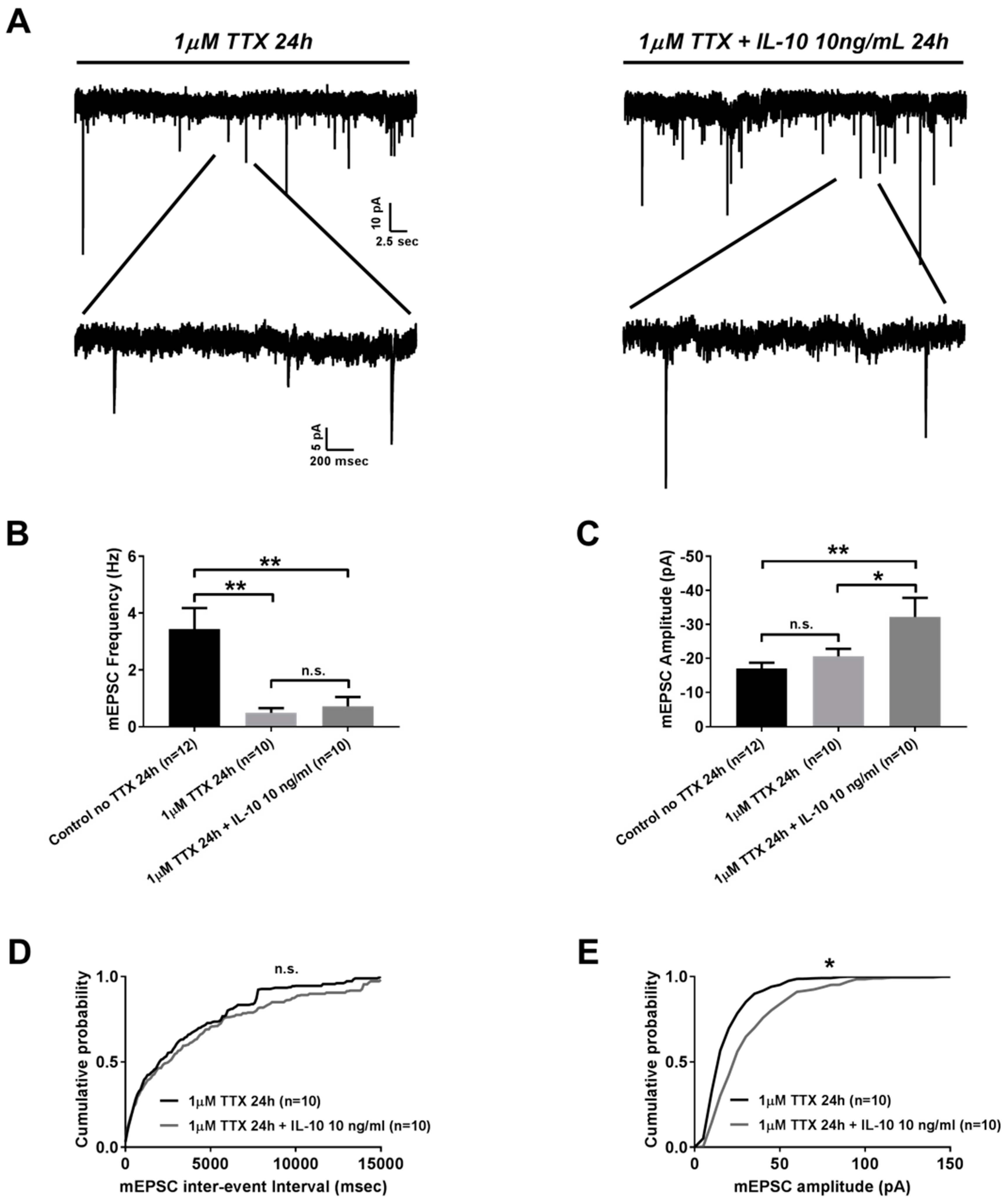
2.2. Effects of Interleukin-10 on Homeostatic Synaptic Plasticity
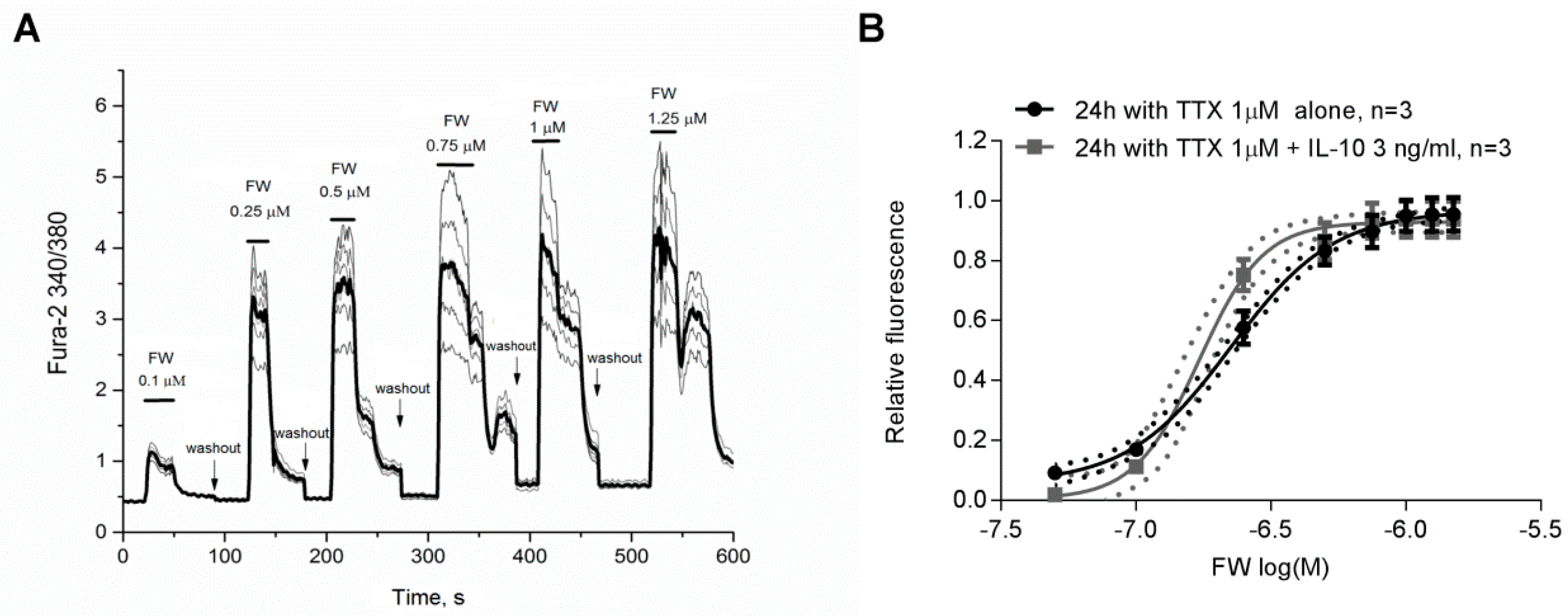
2.3. Effect of IL-10 on AMPA Receptor Agonist 5-Fluorowillardiine-Induced Neuronal Ca2+ Responses at Condition of Homeostatic Synaptic Plasticity
3. Discussion
4. Material and Methods
4.1. Preparation of Primary Hippocampal Culture
4.2. Whole-Cell Patch Clamp Recordings
4.3. Fluorescence Measurements of Neuronal Ca2+ Signals
4.4. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Kwilasz, A.J.; Grace, P.M.; Serbedzija, P.; Maier, S.F.; Watkins, L.R. The therapeutic potential of interleukin-10 in neuroimmune diseases. Neuropharmacology 2015, 96, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.M.; Stillings, S.A.; Leclerc, J.L.; Phillips, H.; Edwards, N.J.; Robicsek, S.A.; Hoh, B.L.; Blackburn, S.; Dore, S. Role of Interleukin-10 in Acute Brain Injuries. Front. Neurol. 2017, 8, 244. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Chen, X.; Liu, Z.; Peng, Y.P.; Qiu, Y.H. Interleukin-10 Protection against Lipopolysaccharide-Induced Neuro-Inflammation and Neurotoxicity in Ventral Mesencephalic Cultures. Int. J. Mol. Sci. 2015, 17, 25. [Google Scholar] [CrossRef] [PubMed]
- Tukhovskaya, E.A.; Turovsky, E.A.; Turovskaya, M.V.; Levin, S.G.; Murashev, A.N.; Zinchenko, V.P.; Godukhin, O.V. Anti-inflammatory cytokine interleukin-10 increases resistance to brain ischemia through modulation of ischemia-induced intracellular Ca(2)(+) response. Neurosci. Lett. 2014, 571, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Levin, S.G.; Godukhin, O.V. Anti-inflammatory cytokines, TGF-beta1 and IL-10, exert anti-hypoxic action and abolish posthypoxic hyperexcitability in hippocampal slice neurons: Comparative aspects. Exp. Neurol 2011, 232, 329–332. [Google Scholar] [CrossRef] [PubMed]
- Joniec-Maciejak, I.; Ciesielska, A.; Wawer, A.; Sznejder-Pacholek, A.; Schwenkgrub, J.; Cudna, A.; Hadaczek, P.; Bankiewicz, K.S.; Czlonkowska, A.; Czlonkowski, A. The influence of AAV2-mediated gene transfer of human IL-10 on neurodegeneration and immune response in a murine model of Parkinson’s disease. Pharm. Rep. 2014, 66, 660–669. [Google Scholar] [CrossRef]
- Kiyota, T.; Ingraham, K.L.; Swan, R.J.; Jacobsen, M.T.; Andrews, S.J.; Ikezu, T. AAV serotype 2/1-mediated gene delivery of anti-inflammatory interleukin-10 enhances neurogenesis and cognitive function in APP+PS1 mice. Gene Ther. 2012, 19, 724–733. [Google Scholar] [CrossRef]
- Grace, P.M.; Loram, L.C.; Christianson, J.P.; Strand, K.A.; Flyer-Adams, J.G.; Penzkover, K.R.; Forsayeth, J.R.; van Dam, A.M.; Mahoney, M.J.; Maier, S.F.; et al. Behavioral assessment of neuropathic pain, fatigue, and anxiety in experimental autoimmune encephalomyelitis (EAE) and attenuation by interleukin-10 gene therapy. Brain Behav. Immun. 2017, 59, 49–54. [Google Scholar] [CrossRef]
- Levin, S.G.; Sirota, N.P.; Nenov, M.N.; Savina, T.A.; Godukhin, O.V. Interleukin-10 and PD150606 modulate expression of AMPA receptor GluA1 and GluA2 subunits under hypoxic conditions. Neuroreport 2018, 29, 84–91. [Google Scholar] [CrossRef]
- Savina, T.A.; Shchipakina, T.G.; Levin, S.G.; Godukhin, O.V. Interleukin–10 prevents the hypoxia–induced decreases in expressions of AMPA receptor subunit GluA1 and alpha subunit of Ca(2+)/calmodulin-dependent protein kinase II in hippocampal neurons. Neurosci. Lett. 2013, 534, 279–284. [Google Scholar] [CrossRef]
- Vezzani, A.; Viviani, B. Neuromodulatory properties of inflammatory cytokines and their impact on neuronal excitability. Neuropharmacology 2015, 96, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Levin, S.G.; Konakov, M.V.; Godukhin, O.V. Role of BK(Ca) Potassium Channels in the Mechanisms of Modulatory Effects of IL-10 on Hypoxia-Induced Changes in Activity of Hippocampal Neurons. Bull. Exp. Biol. Med. 2016, 160, 643–645. [Google Scholar] [CrossRef] [PubMed]
- Turovskaya, M.V.; Turovsky, E.A.; Zinchenko, V.P.; Levin, S.G.; Godukhin, O.V. Interleukin-10 modulates [Ca2+]i response induced by repeated NMDA receptor activation with brief hypoxia through inhibition of InsP(3)-sensitive internal stores in hippocampal neurons. Neurosci. Lett. 2012, 516, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Turovskaya, M.V.; Zinchenko, V.P.; Babaev, A.A.; Epifanova, E.A.; Tarabykin, V.S.; Turovsky, E.A. Mutation in the Sip1 transcription factor leads to a disturbance of the preconditioning of AMPA receptors by episodes of hypoxia in neurons of the cerebral cortex due to changes in their activity and subunit composition. The protective effects of interleukin-10. Arch. Biochem Biophys 2018, 654, 126–135. [Google Scholar] [PubMed]
- Shen, K.F.; Zhu, H.Q.; Wei, X.H.; Wang, J.; Li, Y.Y.; Pang, R.P.; Liu, X.G. Interleukin-10 down-regulates voltage gated sodium channels in rat dorsal root ganglion neurons. Exp. Neurol. 2013, 247, 466–475. [Google Scholar] [CrossRef]
- Suryanarayanan, A.; Carter, J.M.; Landin, J.D.; Morrow, A.L.; Werner, D.F.; Spigelman, I. Role of interleukin-10 (IL–10) in regulation of GABAergic transmission and acute response to ethanol. Neuropharmacology 2016, 107, 181–188. [Google Scholar] [CrossRef]
- He, P.; Liu, Q.; Wu, J.; Shen, Y. Genetic deletion of TNF receptor suppresses excitatory synaptic transmission via reducing AMPA receptor synaptic localization in cortical neurons. FASEB J. 2012, 26, 334–345. [Google Scholar] [CrossRef]
- Kanno, T.; Nagata, T.; Yamamoto, S.; Okamura, H.; Nishizaki, T. Interleukin-18 stimulates synaptically released glutamate and enhances postsynaptic AMPA receptor responses in the CA1 region of mouse hippocampal slices. Brain Res. 2004, 1012, 190–193. [Google Scholar] [CrossRef]
- Cuevas-Olguin, R.; Esquivel-Rendon, E.; Vargas-Mireles, J.; Garcia-Oscos, F.; Miranda-Morales, M.; Salgado, H.; Rose-John, S.; Atzori, M. Interleukin 6 trans-signaling regulates basal synaptic transmission and sensitivity to pentylenetetrazole-induced seizures in mice. Synapse 2017, 71. [Google Scholar] [CrossRef]
- Liu, Q.; Xin, W.; He, P.; Turner, D.; Yin, J.; Gan, Y.; Shi, F.D.; Wu, J. Interleukin-17 inhibits adult hippocampal neurogenesis. Sci. Rep. 2014, 4, 7554. [Google Scholar] [CrossRef]
- Yang, S.; Liu, Z.W.; Wen, L.; Qiao, H.F.; Zhou, W.X.; Zhang, Y.X. Interleukin-1beta enhances NMDA receptor-mediated current but inhibits excitatory synaptic transmission. Brain Res. 2005, 1034, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Kelly, A.; Lynch, A.; Vereker, E.; Nolan, Y.; Queenan, P.; Whittaker, E.; O’Neill, L.A.; Lynch, M.A. The anti-inflammatory cytokine, interleukin (IL)-10, blocks the inhibitory effect of IL-1 beta on long term potentiation. A role for JNK. J. Biol. Chem. 2001, 276, 45564–45572. [Google Scholar] [CrossRef] [PubMed]
- Han, E.B.; Stevens, C.F. Development regulates a switch between post- and presynaptic strengthening in response to activity deprivation. Proc. Natl. Acad. Sci. USA 2009, 106, 10817–10822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, J.J.; Zou, R.X.; He, H.M.; Tang, Y.Q.; Wang, H.L. Effect of Pb Exposure on Synaptic Scaling Through Regulation of AMPA Receptor Surface Trafficking. Toxicol. Sci. 2018, 165, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Stellwagen, D.; Malenka, R.C. Synaptic scaling mediated by glial TNF–alpha. Nature 2006, 440, 1054–1059. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.H.; Park, E.; You, B.; Jung, Y.; Park, A.R.; Park, S.G.; Lee, J.R. Neuronal synapse formation induced by microglia and interleukin 10. PLoS ONE 2013, 8, e81218. [Google Scholar] [CrossRef]
- Almolda, B.; de Labra, C.; Barrera, I.; Gruart, A.; Delgado-Garcia, J.M.; Villacampa, N.; Vilella, A.; Hofer, M.J.; Hidalgo, J.; Campbell, I.L.; et al. Alterations in microglial phenotype and hippocampal neuronal function in transgenic mice with astrocyte-targeted production of interleukin-10. Brain Behav. Immun. 2015, 45, 80–97. [Google Scholar] [CrossRef]
- Ledeboer, A.; Breve, J.J.; Wierinckx, A.; van der Jagt, S.; Bristow, A.F.; Leysen, J.E.; Tilders, F.J.; Van Dam, A.M. Expression and regulation of interleukin-10 and interleukin-10 receptor in rat astroglial and microglial cells. Eur. J. Neurosci. 2002, 16, 1175–1185. [Google Scholar] [CrossRef]
- Norden, D.M.; Fenn, A.M.; Dugan, A.; Godbout, J.P. TGFbeta produced by IL-10 redirected astrocytes attenuates microglial activation. Glia 2014, 62, 881–895. [Google Scholar] [CrossRef]
- De Pitta, M.; Brunel, N.; Volterra, A. Astrocytes: Orchestrating synaptic plasticity? Neuroscience 2016, 323, 43–61. [Google Scholar] [CrossRef] [Green Version]
- Wake, H.; Moorhouse, A.J.; Miyamoto, A.; Nabekura, J. Microglia: Actively surveying and shaping neuronal circuit structure and function. Trends Neurosci. 2013, 36, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Ben Achour, S.; Pascual, O. Glia: The many ways to modulate synaptic plasticity. Neurochem. Int. 2010, 57, 440–445. [Google Scholar] [CrossRef] [PubMed]
- Bai, F.; Xie, C.; Yuan, Y.; Shi, Y.; Zhang, Z. Promoter haplotypes of interleukin-10 gene linked to cortex plasticity in subjects with risk of Alzheimer’s disease. Neuroimage Clin. 2018, 17, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Magalhaes, C.A.; Carvalho, M.D.G.; Sousa, L.P.; Caramelli, P.; Gomes, K.B. Alzheimer’s disease and cytokine IL–10 gene polymorphisms: Is there an association? Arq. Neuropsiquiatr. 2017, 75, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Kosenkov, A.M.; Gaidin, S.G.; Sergeev, A.I.; Teplov, I.Y.; Zinchenko, V.P. Fast changes of NMDA and AMPA receptor activity under acute hyperammonemia in vitro. Neurosci. Lett. 2018, 686, 80–86. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nenov, M.N.; Konakov, M.V.; Teplov, I.Y.; Levin, S.G. Interleukin-10 Facilitates Glutamatergic Synaptic Transmission and Homeostatic Plasticity in Cultured Hippocampal Neurons. Int. J. Mol. Sci. 2019, 20, 3375. https://doi.org/10.3390/ijms20133375
Nenov MN, Konakov MV, Teplov IY, Levin SG. Interleukin-10 Facilitates Glutamatergic Synaptic Transmission and Homeostatic Plasticity in Cultured Hippocampal Neurons. International Journal of Molecular Sciences. 2019; 20(13):3375. https://doi.org/10.3390/ijms20133375
Chicago/Turabian StyleNenov, Miroslav N., Maxim V. Konakov, Ilia Y. Teplov, and Sergey G. Levin. 2019. "Interleukin-10 Facilitates Glutamatergic Synaptic Transmission and Homeostatic Plasticity in Cultured Hippocampal Neurons" International Journal of Molecular Sciences 20, no. 13: 3375. https://doi.org/10.3390/ijms20133375
APA StyleNenov, M. N., Konakov, M. V., Teplov, I. Y., & Levin, S. G. (2019). Interleukin-10 Facilitates Glutamatergic Synaptic Transmission and Homeostatic Plasticity in Cultured Hippocampal Neurons. International Journal of Molecular Sciences, 20(13), 3375. https://doi.org/10.3390/ijms20133375