Bacterial Colonization of the Female Upper Genital Tract


Abstract
:1. The Female Upper Genital Tract—Not as Sterile as Presumed?
2. Upper Genital Tract Microbiota Sampling and Challenges
3. Endometrial Microbiota
4. Bacterial Colonization of Other Sites of the Upper Genital Tract (UGT)
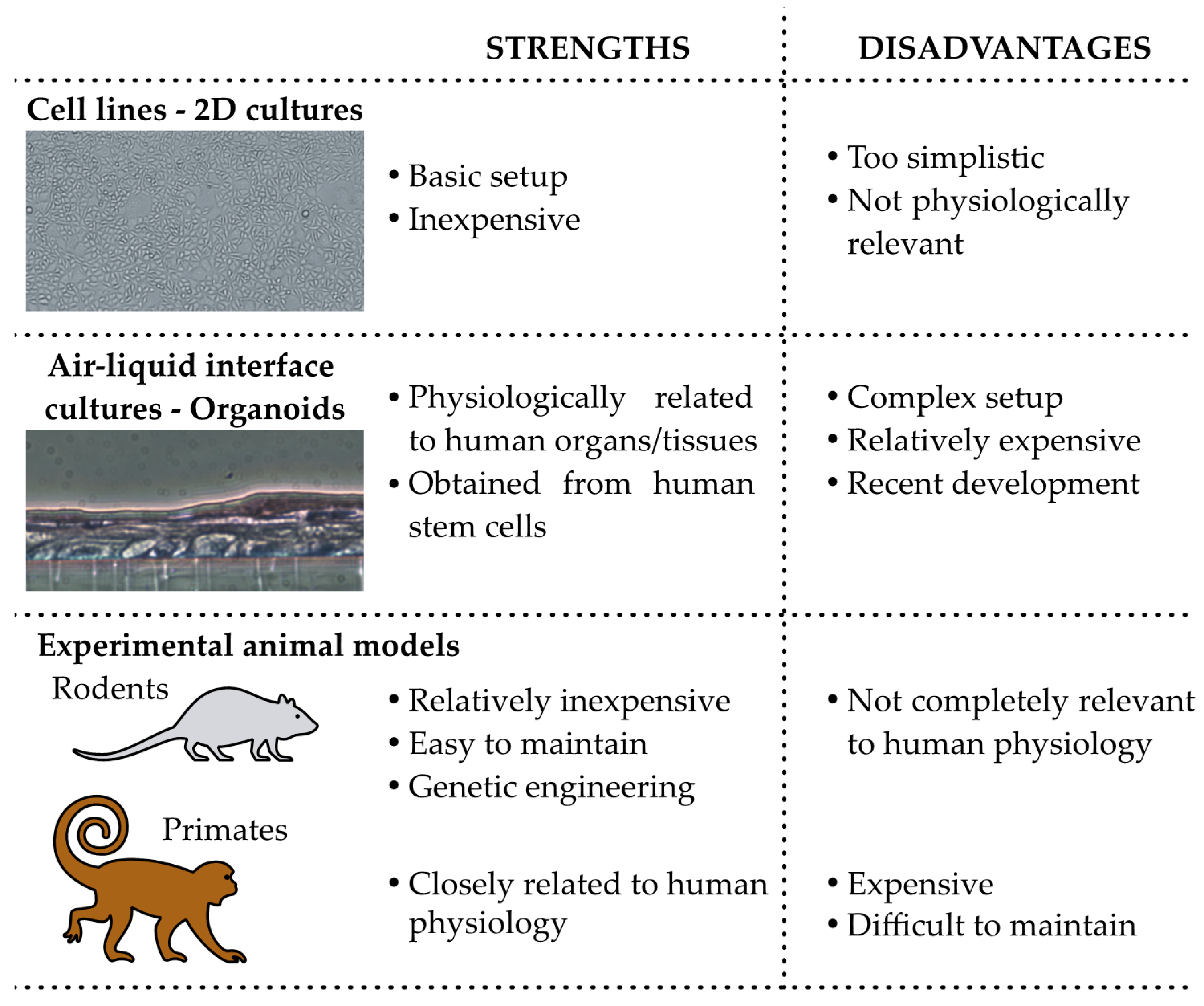
5. Experimental Assessment of the Role of Uterine Microbiota: Animal and in Vitro Models
6. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Rosenberg, E.; Zilber-Rosenberg, I. The hologenome concept of evolution after 10 years. Microbiome 2018, 6, 78. [Google Scholar] [CrossRef] [PubMed]
- Belizário, J.E.; Napolitano, M. Human microbiomes and their roles in dysbiosis, common diseases, and novel therapeutic approaches. Front. Microbiol. 2015, 6, 1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominguez-Bello, M.G.; Godoy-Vitorino, F.; Knight, R.; Blaser, M.J. Role of the microbiome in human development. Gut 2019, 68, 1108–1114. [Google Scholar] [CrossRef] [PubMed]
- Van de Guchte, M.; Blottière, H.M.; Doré, J. Humans as holobionts: Implications for prevention and therapy. Microbiome 2018, 6, 81. [Google Scholar] [CrossRef] [PubMed]
- Younes, J.A.; Lievens, E.; Hummelen, R.; van der Westen, R.; Reid, G.; Petrova, M.I. Women and Their Microbes: The Unexpected Friendship. Trends Microbiol. 2018, 26, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Anahtar, M.N.; Gootenberg, D.B.; Mitchell, C.M.; Kwon, D.S. Cervicovaginal Microbiota and Reproductive Health: The Virtue of Simplicity. Cell Host Microbe 2018, 23, 159–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravel, J.; Gajer, P.; Abdo, Z.; Schneider, G.M.; Koenig, S.S.K.; McCulle, S.L.; Karlebach, S.; Gorle, R.; Russell, J.; Tacket, C.O.; et al. Vaginal microbiome of reproductive-age women. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4680–4687. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.M.; Chase, D.M.; Herbst-Kralovetz, M.M. Uterine Microbiota: Residents, Tourists, or Invaders? Front. Immunol. 2018, 9, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, D.M.; Ma, J.; Prince, A.L.; Antony, K.M.; Seferovic, M.D.; Aagaard, K.M. Maturation of the infant microbiome community structure and function across multiple body sites and in relation to mode of delivery. Nat. Med. 2017, 23, 314–326. [Google Scholar] [CrossRef] [Green Version]
- Mor, A.; Driggers, P.H.; Segars, J.H. Molecular characterization of the human microbiome from a reproductive perspective. Fertil. Steril. 2015, 104, 1344–1350. [Google Scholar] [CrossRef]
- Fox, C.; Eichelberger, K. Maternal microbiome and pregnancy outcomes. Fertil. Steril. 2015, 104, 1358–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capoccia, R.; Greub, G.; Baud, D. Ureaplasma urealyticum, Mycoplasma hominis and adverse pregnancy outcomes. Curr. Opin. Infect. Dis. 2013, 26, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Ammerdorffer, A.; Stojanov, M.; Greub, G.; Baud, D. Chlamydia trachomatis and chlamydia-like bacteria: New enemies of human pregnancies. Curr. Opin. Infect. Dis. 2017, 30, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.A.; Beasley, D.E.; Dunn, R.R.; Archie, E.A. Lactobacilli Dominance and Vaginal pH: Why Is the Human Vaginal Microbiome Unique? Front. Microbiol. 2016, 7, 1936. [Google Scholar] [CrossRef] [PubMed]
- Amabebe, E.; Anumba, D.O.C. The Vaginal Microenvironment: The Physiologic Role of Lactobacilli. Front. Med. 2018, 5, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovachev, S. Defence factors of vaginal lactobacilli. Crit. Rev. Microbiol. 2018, 44, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.-S.; Ehlers, M.M.; Lombaard, H.; Redelinghuys, M.J.; Kock, M.M. Etiology of bacterial vaginosis and polymicrobial biofilm formation. Crit. Rev. Microbiol. 2017, 43, 651–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravel, J.; Brotman, R.M.; Gajer, P.; Ma, B.; Nandy, M.; Fadrosh, D.W.; Sakamoto, J.; Koenig, S.S.; Fu, L.; Zhou, X.; et al. Daily temporal dynamics of vaginal microbiota before, during and after episodes of bacterial vaginosis. Microbiome 2013, 1, 29. [Google Scholar] [CrossRef] [PubMed]
- Goldenberg, R.L.; Culhane, J.F.; Iams, J.D.; Romero, R. Epidemiology and causes of preterm birth. Lancet 2008, 371, 75–84. [Google Scholar] [CrossRef]
- Hansen, L.K.; Becher, N.; Bastholm, S.; Glavind, J.; Ramsing, M.; Kim, C.J.; Romero, R.; Jensen, J.S.; Uldbjerg, N. The cervical mucus plug inhibits, but does not block, the passage of ascending bacteria from the vagina during pregnancy. Acta Obstet. Gynecol. Scand. 2014, 93, 102–108. [Google Scholar] [CrossRef]
- Becher, N.; Waldorf, K.A.; Hein, M.; Uldbjerg, N. The cervical mucus plug: Structured review of the literature. Acta Obstet. Gynecol. Scand. 2009, 88, 502–513. [Google Scholar] [CrossRef]
- Zervomanolakis, I.; Ott, H.W.; Hadziomerovic, D.; Mattle, V.; Seeber, B.E.; Virgolini, I.; Heute, D.; Kissler, S.; Leyendecker, G.; Wildt, L. Physiology of upward transport in the human female genital tract. Ann. N. Y. Acad. Sci. 2007, 1101, 1–20. [Google Scholar] [CrossRef]
- Kunz, G.; Beil, D.; Deininger, H.; Wildt, L.; Leyendecker, G. The dynamics of rapid sperm transport through the female genital tract: Evidence from vaginal sonography of uterine peristalsis and hysterosalpingoscintigraphy. Hum. Reprod. 1996, 11, 627–632. [Google Scholar] [CrossRef]
- Bulletti, C.; de Ziegler, D.; Polli, V.; Diotallevi, L.; Del Ferro, E.; Flamigni, C. Uterine contractility during the menstrual cycle. Hum. Reprod. 2000, 15 (Suppl. 1), 81–89. [Google Scholar] [CrossRef] [Green Version]
- Wildt, L.; Kissler, S.; Licht, P.; Becker, W. Sperm transport in the human female genital tract and its modulation by oxytocin as assessed by hysterosalpingoscintigraphy, hysterotonography, electrohysterography and Doppler sonography. Hum. Reprod. Update 1998, 4, 655–666. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Song, X.; Wei, W.; Zhong, H.; Dai, J.; Lan, Z.; Li, F.; Yu, X.; Feng, Q.; Wang, Z.; et al. The microbiota continuum along the female reproductive tract and its relation to uterine-related diseases. Nat. Commun. 2017, 8, 875. [Google Scholar] [CrossRef]
- Moreno, I.; Codoñer, F.M.; Vilella, F.; Valbuena, D.; Martinez-Blanch, J.F.; Jimenez-Almazán, J.; Alonso, R.; Alamá, P.; Remohí, J.; Pellicer, A.; et al. Evidence that the endometrial microbiota has an effect on implantation success or failure. Am. J. Obstet. Gynecol. 2016, 215, 684–703. [Google Scholar] [CrossRef] [Green Version]
- Franasiak, J.M.; Werner, M.D.; Juneau, C.R.; Tao, X.; Landis, J.; Zhan, Y.; Treff, N.R.; Scott, R.T. Endometrial microbiome at the time of embryo transfer: Next-generation sequencing of the 16S ribosomal subunit. J. Assist. Reprod. Genet. 2016, 33, 129–136. [Google Scholar] [CrossRef]
- Mitchell, C.M.; Haick, A.; Nkwopara, E.; Garcia, R.; Rendi, M.; Agnew, K.; Fredricks, D.N.; Eschenbach, D. Colonization of the upper genital tract by vaginal bacterial species in nonpregnant women. Am. J. Obstet. Gynecol. 2015, 212, 611.e1-9. [Google Scholar] [CrossRef]
- Miles, S.M.; Hardy, B.L.; Merrell, D.S. Investigation of the microbiota of the reproductive tract in women undergoing a total hysterectomy and bilateral salpingo-oopherectomy. Fertil. Steril. 2017, 107, 813–820.e1. [Google Scholar] [CrossRef] [Green Version]
- Moreno, I.; Simon, C. Relevance of assessing the uterine microbiota in infertility. Fertil. Steril. 2018, 110, 337–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, X.; Franasiak, J.M.; Zhan, Y.; Scott, R.T.; Rajchel, J.; Bedard, J.; Newby, R.; Scott, R.T.; Treff, N.R.; Chu, T. Characterizing the endometrial microbiome by analyzing the ultra-low bacteria from embryo transfer catheter tips in IVF cycles: Next generation sequencing (NGS) analysis of the 16S ribosomal gene. Hum. Microbiome J. 2017, 3, 15–21. [Google Scholar] [CrossRef]
- Verstraelen, H.; Vilchez-Vargas, R.; Desimpel, F.; Jauregui, R.; Vankeirsbilck, N.; Weyers, S.; Verhelst, R.; De Sutter, P.; Pieper, D.H.; Van De Wiele, T. Characterisation of the human uterine microbiome in non-pregnant women through deep sequencing of the V1-2 region of the 16S rRNA gene. PeerJ 2016, 4, e1602. [Google Scholar] [CrossRef] [PubMed]
- De Goffau, M.C.; Lager, S.; Salter, S.J.; Wagner, J.; Kronbichler, A.; Charnock-Jones, D.S.; Peacock, S.J.; Smith, G.C.S.; Parkhill, J. Recognizing the reagent microbiome. Nat. Microbiol. 2018, 3, 851–853. [Google Scholar] [CrossRef] [PubMed]
- Salter, S.J.; Cox, M.J.; Turek, E.M.; Calus, S.T.; Cookson, W.O.; Moffatt, M.F.; Turner, P.; Parkhill, J.; Loman, N.J.; Walker, A.W. Reagent and laboratory contamination can critically impact sequence-based microbiome analyses. BMC Biol. 2014, 12, 87. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Hofstaedter, C.E.; Zhao, C.; Mattei, L.; Tanes, C.; Clarke, E.; Lauder, A.; Sherrill-Mix, S.; Chehoud, C.; Kelsen, J.; et al. Optimizing methods and dodging pitfalls in microbiome research. Microbiome 2017, 5, 52. [Google Scholar] [CrossRef] [PubMed]
- Muhleisen, A.L.; Herbst-Kralovetz, M.M. Menopause and the vaginal microbiome. Maturitas 2016, 91, 42–50. [Google Scholar] [CrossRef]
- Walther-António, M.R.S.; Chen, J.; Multinu, F.; Hokenstad, A.; Distad, T.J.; Cheek, E.H.; Keeney, G.L.; Creedon, D.J.; Nelson, H.; Mariani, A.; et al. Potential contribution of the uterine microbiome in the development of endometrial cancer. Genome Med. 2016, 8, 122. [Google Scholar] [CrossRef]
- Strowitzki, T.; Germeyer, A.; Popovici, R.; von Wolff, M. The human endometrium as a fertility-determining factor. Hum. Reprod. Update 2006, 12, 617–630. [Google Scholar] [CrossRef] [Green Version]
- Simón, C.; Martín, J.C.; Pellicer, A. Paracrine regulators of implantation. Best Pract. Res. Clin. Obstet. Gynaecol. 2000, 14, 815–826. [Google Scholar] [CrossRef]
- Fang, R.-L.; Chen, L.-X.; Shu, W.-S.; Yao, S.-Z.; Wang, S.-W.; Chen, Y.-Q. Barcoded sequencing reveals diverse intrauterine microbiomes in patients suffering with endometrial polyps. Am. J. Transl. Res. 2016, 8, 1581. [Google Scholar] [PubMed]
- Kyono, K.; Hashimoto, T.; Kikuchi, S.; Nagai, Y.; Sakuraba, Y. A pilot study and case reports on endometrial microbiota and pregnancy outcome: An analysis using 16S rRNA gene sequencing among IVF patients, and trial therapeutic intervention for dysbiotic endometrium. Reprod. Med. Biol. 2019, 18, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.N.; Fujishita, A.; Masumoto, H.; Muto, H.; Kitajima, M.; Masuzaki, H.; Kitawaki, J. Molecular detection of intrauterine microbial colonization in women with endometriosis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2016, 199, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Pelzer, E.S.; Willner, D.; Buttini, M.; Huygens, F. A role for the endometrial microbiome in dysfunctional menstrual bleeding. Antonie van Leeuwenhoek 2018, 111, 933–943. [Google Scholar] [CrossRef] [PubMed]
- Kyono, K.; Hashimoto, T.; Nagai, Y.; Sakuraba, Y. Analysis of endometrial microbiota by 16S ribosomal RNA gene sequencing among infertile patients: a single-center pilot study. Reprod. Med. Biol. 2018, 17, 297–306. [Google Scholar] [CrossRef] [PubMed]
- DiGiulio, D.B.; Callahan, B.J.; McMurdie, P.J.; Costello, E.K.; Lyell, D.J.; Robaczewska, A.; Sun, C.L.; Goltsman, D.S.A.; Wong, R.J.; Shaw, G.; et al. Temporal and spatial variation of the human microbiota during pregnancy. Proc. Natl. Acad. Sci. USA 2015, 112, 11060–11065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelzer, E.S.; Willner, D.; Buttini, M.; Hafner, L.M.; Theodoropoulos, C.; Huygens, F. The fallopian tube microbiome: Implications for reproductive health. Oncotarget 2018, 9, 21541. [Google Scholar] [CrossRef]
- Zhou, B.; Sun, C.; Huang, J.; Xia, M.; Guo, E.; Li, N.; Lu, H.; Shan, W.; Wu, Y.; Li, Y.; et al. The biodiversity Composition of Microbiome in Ovarian Carcinoma Patients. Sci. Rep. 2019, 9, 1691. [Google Scholar] [CrossRef]
- Banerjee, S.; Tian, T.; Wei, Z.; Shih, N.; Feldman, M.D.; Coukos, G.; Alwine, J.C.; Robertson, E.S. The ovarian cancer oncobiome. Oncotarget 2017, 8, 36225. [Google Scholar] [CrossRef]
- Aagaard, K.; Ma, J.; Antony, K.M.; Ganu, R.; Petrosino, J.; Versalovic, J. The placenta harbors a unique microbiome. Sci. Transl. Med. 2014, 6, 237ra65. [Google Scholar] [CrossRef]
- Doyle, R.M.; Harris, K.; Kamiza, S.; Harjunmaa, U.; Ashorn, U.; Nkhoma, M.; Dewey, K.G.; Maleta, K.; Ashorn, P.; Klein, N. Bacterial communities found in placental tissues are associated with severe chorioamnionitis and adverse birth outcomes. PLoS ONE 2017, 12, e0180167. [Google Scholar] [CrossRef] [PubMed]
- Parnell, L.A.; Briggs, C.M.; Cao, B.; Delannoy-Bruno, O.; Schrieffer, A.E.; Mysorekar, I.U. Microbial communities in placentas from term normal pregnancy exhibit spatially variable profiles. Sci. Rep. 2017, 7, 11200. [Google Scholar] [CrossRef] [PubMed]
- Seferovic, M.D.; Pace, R.M.; Caroll, M.; Belfort, B.; Major, A.M.; Chu, D.M.; Racusin, D.A.; Castro, E.C.C.; Muldrew, K.L.; Versalovic, J.; et al. Visualization of Microbes by 16S in situ Hybridization in Term and Preterm Placentae without Intraamniotic Infection. Am. J. Obstet. Gynecol. 2019, in press. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, M.S.; Bommarito, P.A.; Martin, E.M.; Smeester, L.; Fichorova, R.N.; Onderdonk, A.B.; Kuban, K.C.K.; O’Shea, T.M.; Fry, R.C. Microorganisms in the human placenta are associated with altered CpG methylation of immune and inflammation-related genes. PLoS ONE 2017, 12, e0188664. [Google Scholar] [CrossRef] [PubMed]
- Prince, A.L.; Ma, J.; Kannan, P.S.; Alvarez, M.; Gisslen, T.; Harris, R.A.; Sweeney, E.L.; Knox, C.L.; Lambers, D.S.; Jobe, A.H.; et al. The placental membrane microbiome is altered among subjects with spontaneous preterm birth with and without chorioamnionitis. Am. J. Obstet. Gynecol. 2016, 214, 627.e1–627.e16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leiby, J.S.; McCormick, K.; Sherrill-Mix, S.; Clarke, E.L.; Kessler, L.R.; Taylor, L.J.; Hofstaedter, C.E.; Roche, A.M.; Mattei, L.M.; Bittinger, K.; et al. Lack of detection of a human placenta microbiome in samples from preterm and term deliveries. Microbiome 2018, 6, 196. [Google Scholar] [CrossRef] [PubMed]
- Lauder, A.P.; Roche, A.M.; Sherrill-Mix, S.; Bailey, A.; Laughlin, A.L.; Bittinger, K.; Leite, R.; Elovitz, M.A.; Parry, S.; Bushman, F.D. Comparison of placenta samples with contamination controls does not provide evidence for a distinct placenta microbiota. Microbiome 2016, 4, 29. [Google Scholar] [CrossRef]
- Bushman, F.D. De-Discovery of the Placenta Microbiome. Am. J. Obstet. Gynecol. 2019, 220, 213–214. [Google Scholar] [CrossRef] [Green Version]
- Theis, K.R.; Romero, R.; Winters, A.D.; Greenberg, J.M.; Gomez-Lopez, N.; Alhousseini, A.; Bieda, J.; Maymon, E.; Pacora, P.; Fettweis, J.M.; et al. Does the human placenta delivered at term have a microbiota? Results of cultivation, quantitative real-time PCR, 16S rRNA gene sequencing, and metagenomics. Am. J. Obstet. Gynecol. 2019, 220, 267.e1–267.e39. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Arango, L.F.; Barrett, H.L.; McIntyre, H.D.; Callaway, L.K.; Morrison, M.; Nitert, M.D. Contributions of the maternal oral and gut microbiome to placental microbial colonization in overweight and obese pregnant women. Sci. Rep. 2017, 7, 2860. [Google Scholar] [CrossRef]
- Bassols, J.; Serino, M.; Carreras-Badosa, G.; Burcelin, R.; Blasco-Baque, V.; Lopez-Bermejo, A.; Fernandez-Real, J.-M. Gestational diabetes is associated with changes in placental microbiota and microbiome. Pediatr. Res. 2016, 80, 777–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franklin, C.L.; Ericsson, A.C. Microbiota and reproducibility of rodent models. Lab Anim. 2017, 46, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Gondek, D.C.; Olive, A.J.; Stary, G.; Starnbach, M.N. CD4+ T Cells Are Necessary and Sufficient to Confer Protection against Chlamydia trachomatis Infection in the Murine Upper Genital Tract. J. Immunol. 2012, 189, 2441–2449. [Google Scholar] [CrossRef] [PubMed]
- Hafner, L.M. Pathogenesis of fallopian tube damage caused by Chlamydia trachomatis infections. Contraception 2015, 92, 108–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bebear, C.; de Barbeyrac, B. Genital Chlamydia trachomatis infections. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2009, 15, 4–10. [Google Scholar] [Green Version]
- McGowin, C.L.; Spagnuolo, R.A.; Pyles, R.B. Mycoplasma genitalium Rapidly Disseminates to the Upper Reproductive Tracts and Knees of Female Mice following Vaginal Inoculation. Infect. Immun. 2010, 78, 726–736. [Google Scholar] [CrossRef] [PubMed]
- McGowin, C.L.; Totten, P.A. The Unique Microbiology and Molecular Pathogenesis of Mycoplasma genitalium. J. Infect. Dis. 2017, 216, S382–S388. [Google Scholar] [CrossRef] [Green Version]
- Wiesenfeld, H.C.; Manhart, L.E. Mycoplasma genitalium in Women: Current Knowledge and Research Priorities for This Recently Emerged Pathogen. J. Infect. Dis. 2017, 216, S389–S395. [Google Scholar] [CrossRef]
- Francis, I.P.; Islam, E.A.; Gower, A.C.; Shaik-Dasthagirisaheb, Y.B.; Gray-Owen, S.D.; Wetzler, L.M. Murine host response to Neisseria gonorrhoeae upper genital tract infection reveals a common transcriptional signature, plus distinct inflammatory responses that vary between reproductive cycle phases. BMC Genomics 2018, 19, 627. [Google Scholar] [CrossRef]
- Lenz, J.D.; Dillard, J.P. Pathogenesis of Neisseria gonorrhoeae and the Host Defense in Ascending Infections of Human Fallopian Tube. Front. Immunol. 2018, 9, 2710. [Google Scholar] [CrossRef]
- Rice, P.A.; Shafer, W.M.; Ram, S.; Jerse, A.E. Neisseria gonorrhoeae: Drug Resistance, Mouse Models, and Vaccine Development. Annu. Rev. Microbiol. 2017, 71, 665–686. [Google Scholar] [CrossRef] [PubMed]
- Sturdevant, G.L.; Caldwell, H.D. Innate immunity is sufficient for the clearance of Chlamydia trachomatis from the female mouse genital tract. Pathog. Dis. 2014, 72, 70–73. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.J.; Vieira-Neto, A.; Gobikrushanth, M.; Daetz, R.; Mingoti, R.D.; Parize, A.C.B.; de Freitas, S.L.; da Costa, A.N.L.; Bicalho, R.C.; Lima, S.; et al. Uterine Microbiota Progression from Calving until Establishment of Metritis in Dairy Cows. Appl. Environ. Microbiol. 2015, 81, 6324–6332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knudsen, L.R.V.; Karstrup, C.C.; Pedersen, H.G.; Agerholm, J.S.; Jensen, T.K.; Klitgaard, K. Revisiting bovine pyometra--new insights into the disease using a culture-independent deep sequencing approach. Vet. Microbiol. 2015, 175, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.-L.; Liu, M.-C.; Xu, J.; An, L.-G.; Wang, J.-F.; Zhu, Y.-H. Uterine Microbiota of Dairy Cows with Clinical and Subclinical Endometritis. Front. Microbiol. 2018, 9, 2691. [Google Scholar] [CrossRef] [PubMed]
- Lyman, C.C.; Holyoak, G.R.; Meinkoth, K.; Wieneke, X.; Chillemi, K.A.; DeSilva, U. Canine endometrial and vaginal microbiomes reveal distinct and complex ecosystems. PLoS ONE 2019, 14, e0210157. [Google Scholar] [CrossRef] [PubMed]
- Deane, J.A.; Cousins, F.L.; Gargett, C.E. Endometrial organoids: In vitro models for endometrial research and personalized medicine. Biol. Reprod. 2017, 97, 781–783. [Google Scholar] [CrossRef]
- Łaniewski, P.; Gomez, A.; Hire, G.; So, M.; Herbst-Kralovetz, M.M. Human Three-Dimensional Endometrial Epithelial Cell Model to Study Host Interactions with Vaginal Bacteria and Neisseria gonorrhoeae. Infect. Immun. 2017, 85, e01049-16. [Google Scholar] [CrossRef]
- Weimar, C.H.E.; Post Uiterweer, E.D.; Teklenburg, G.; Heijnen, C.J.; Macklon, N.S. In-vitro model systems for the study of human embryo–endometrium interactions. Reprod. Biomed. Online 2013, 27, 461–476. [Google Scholar] [CrossRef]
- Li, D.; Li, H.; Wang, Y.; Eldomany, A.; Wu, J.; Yuan, C.; Xue, J.; Shi, J.; Jia, Y.; Ha, C.; et al. Development and characterization of a polarized human endometrial cell epithelia in an air–liquid interface state. Stem Cell Res. Ther. 2018, 9, 209. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Characteristic | Vagina (Lower Genital Tract) | Uterus (Upper Genital Tract) |
|---|---|---|
| Sampling procedure | • Low invasiveness • Simple (vaginal swab) | • Invasive methods • Through the vaginal–cervical route, after hysterectomy or during laparoscopic surgery |
| Target patients | • Can be routinely performed in all type of patients | • Patients with endometrial pathologies (endometriosis or cancer) or reproductive issues |
| Contamination risk during sampling | • Low risk of contamination | • Risks of contamination by vaginal microbiota when accessing uterus through the vaginal–cervical route |
| Data interpretation | • Well-defined vaginal microbiota based on a large number of studies | • Scarcely studied • Comparison between studies is difficult due to the heterogeneity of targeted patient populations • Difficult to obtain healthy controls |
| Study | Study Targets | Sample Type | Sequencing Method and Target | Country | Major Genera | Impact on the Outcome 1 | Ref. |
|---|---|---|---|---|---|---|---|
| Franasiak et al. (2016) | Patients undergoing in vitro fertilization (IVF) (n = 33) • 18 in ongoing pregnancies • 15 without pregnancies | Embryo transfer catheter tips | Ion Torrent • V2,3,4,6,7,8,9 | USA | Flavobacterium spp. | NA | [28] |
| Lactobacillus spp. | NA | ||||||
| Limnohabitans spp. | NA | ||||||
| Polynucleobacter spp. | NA | ||||||
| Bdellovibrio spp. | NA | ||||||
| Chryseobacterium spp. | NA | ||||||
| Spirochaeta spp. | NA | ||||||
| Clostridium spp. | NA | ||||||
| Verstraelen et al. (2016) | Patients undergoing hysteroscopy (n = 19) • 11 with recurrent implantation failure • 7 recurrent pregnancy loss • 1 with both | Endometrial sampler | Illumina • V1–V2 | Belgium | Bacteroides spp. (dominant) | NA | [33] |
| Lactobacillus crispatus | NA | ||||||
| Pelomonas spp. | NA | ||||||
| Prevotella spp. | NA | ||||||
| Atopobium vaginae | NA | ||||||
| Mobiluncus curtisii | NA | ||||||
| Khan et al. (2016) | Patients with endometriosis (n = 32) • 16 with endometriosis • 16 controls without endometriosis | Endometrium and cystic fluid | Illumina • Custom primers | Japan | Lactobacillacae | + | [43] |
| Streptococcaceae | − | ||||||
| Staphylococaceae | − | ||||||
| Enterobacteriaceae | NA | ||||||
| Moraxellaceae | − | ||||||
| Fang et al. (2016) | Patients with endometrial polyps (n = 30) • 20 with endometrial polyps • 10 healthy women | Endometrial swabs and endometrial tissues | Illumina • V4 | China | Lactobacillus | − | [41] |
| Bifidobacterium | − | ||||||
| Gardnerella | − | ||||||
| Streptococcus | − | ||||||
| Alteromonas | − | ||||||
| Enterobacter | + | ||||||
| Pseudomonas | + | ||||||
| Moreno et al. (2016) * | Patients undergoing IVF (n = 70) • 35 undergoing IVF • 35 fertile women | Endometrial fluid | 454 pyrosequencing • V3–V5 | Spain | Lactobacillus spp. (dominant) | + | [27] |
| Gardenrella spp. | − | ||||||
| Bifidobacterium spp. | − | ||||||
| Streptococcus spp. | − | ||||||
| Prevotella spp. | − | ||||||
| Walther-António et al. (2016) * | Women undergoing hysterectomy (n = 31) • 17 with endometrial cancer • 4 with endometrial hyperplasia • 10 with benign gynecologic condition | Uterine biopsies | Illumina • V3–V5 | USA | Shigella spp. | NA | [38] |
| Barnesiella spp. | NA | ||||||
| Staphylococcus spp. | + | ||||||
| Blautia spp. | + | ||||||
| Parabacteroides spp. | + | ||||||
| Bacteroides spp. | − | ||||||
| Faecalibacterium spp. | − | ||||||
| Miles et al. (2016) * | Women undergoing a total hysterectomy and bilateral salpingo-oopherectomy (n = 10) • Patients with various pathologies | Endometrium tissue | 454 pyrosequencing • V1–V3 | USA | Lactobacillus spp. | NA | [30] |
| Corynebacterium spp. | NA | ||||||
| Staphylococcus spp. | NA | ||||||
| Acinetobacter spp. | NA | ||||||
| Blautia spp. | NA | ||||||
| Tao et al. (2017) | Patients undergoing IVF (n = 70) | Embryo transfer catheter tips | Illumina • V4 | USA | Lactobacillus spp. (dominant) | NA | [32] |
| Corynebacterium spp. | NA | ||||||
| Bifidobacterium spp. | NA | ||||||
| Staphylococcus spp. | NA | ||||||
| Streptococcus spp. | NA | ||||||
| Chen et al. (2017) * | Patients without infection (n = 110) | Endometrium | Ion Torrent • V4–V5 | China | Lactobacillus spp. (30.6%) | NA | [26] |
| Pseudomonas spp. (9.09%) | NA | ||||||
| Acinetobacter spp. (9.07%) | NA | ||||||
| Vagococcus spp. (7.29%) | NA | ||||||
| Sphingobium spp. (5%) | NA | ||||||
| Pelzer et al. (2018) | Patients undergoing operative hysteroscopy or laparoscopy (n = 145) | Endocervical swabs | 454 pyrosequencing • V5 and V8 | Australia | Lactobacillus spp. (dominant) | NA | [44] |
| Gardnerella spp. | NA | ||||||
| Veillonella spp. | NA | ||||||
| Prevotella spp. | NA | ||||||
| Sneathia spp. | NA | ||||||
| Jonquetella spp. | NA | ||||||
| Fusobacterium spp. | NA | ||||||
| Kyono et al. (2018) * | Patients with infertility (n = 109) • 79 undergoing IVF • 23 non-IVF cases • 7 fertile volunteers | Endometrial fluid | Illumina • V4 with modified primers | Japan | Lactobacillus spp. | + | [45] |
| Gardnerella spp. | NA | ||||||
| Streptococcus spp. | NA | ||||||
| Atopobium spp. | NA | ||||||
| Bifidobacterium spp. | NA | ||||||
| Sneathia spp. | NA | ||||||
| Prevotella spp. | NA | ||||||
| Staphylococcus spp. | NA | ||||||
| Kyono et al. (2019) | Patients undergoing IVF (n = 92) • 56 Lactobacillus-dominant microbiotas • 36 non-Lactobacillus-dominant microbiotas | Endometrial fluid | N/A | Japan | Lactobacillus spp. | + | [42] |
| Atopobium spp. | NA | ||||||
| Bifidobacterium spp. | + | ||||||
| Gardnerella spp. | NA | ||||||
| Megasphaera spp. | NA | ||||||
| Sneathia spp. | NA | ||||||
| Prevotella spp. | NA |
| Study | Study Targets | Sampling | Sequencing Method and Target | Country | Major Taxa | Ref. |
|---|---|---|---|---|---|---|
| Fallopian Tubes | ||||||
| Miles et al. (2016) | Women undergoing a total hysterectomy and bilateral salpingo-oopherectomy (n = 10) • Patients with various pathologies | Ampullary region of the transected fallopian tubes | 454 pyrosequencing • V1–V3 | USA | Firmicutes | [30] |
| Proteobacteria | ||||||
| Actinobacteria | ||||||
| Bacteroidetes | ||||||
| Chen et al. (2017) | Patients without infection (n = 110) | Laparoscopy and laparotomy | Ion Torrent • V4–V5 | China | Acinetobacter spp. | [26] |
| Comamonas spp. | ||||||
| Pseudomonas spp. | ||||||
| Dysgonomonas spp. | ||||||
| Vagococcus spp. | ||||||
| Delftia spp. | ||||||
| Pelzer et al. (2018) | Women undergoing total hysterectomy (n = 16) | Fallopian tube dissection | 454 pyrosequencing • V5–V8 | Australia | Staphylococcus spp. | [47] |
| Enterococcus spp. | ||||||
| Lactobacillus spp. | ||||||
| Pseudomonas spp. | ||||||
| Burkholderia spp. | ||||||
| Propionibacterium spp. | ||||||
| Zhou et al. (2019) | Women diagnosed with high-grade serous ovarian cancer (n = 50) • 25 cases • 25 controls | Distal fallopian tube tissues (used as control) | Illumina • V3–V4 | China | Proteobacteria | [48] |
| Firmicutes | ||||||
| Bacteroidetes | ||||||
| Actinobacteria | ||||||
| Ovary | ||||||
| Miles et al. (2016) | Women undergoing a total hysterectomy and bilateral salpingo-oopherectomy (n = 10) Patients with various pathologies | Ovary dissection | 454 pyrosequencing • V1–V3 | USA | Firmicutes | [30] |
| Proteobacteria | ||||||
| Actinobacteria | ||||||
| Bacteroidetes | ||||||
| Verrucomicrobia | ||||||
| Banerjee et al. (2017) | Women with ovarian cancer (primary and recurrent or metastatic tumors of ovarian origin, n = 139) • 99 ovarian cancer samples • 20 matched • 20 unmatched control | Ovarian cancer tissues | Illumina • Custom | USA | Proteobacteria | [49] |
| Firmicutes | ||||||
| Bacteroidetes | ||||||
| Chlamydiae | ||||||
| Spirochaetes | ||||||
| Tenericutes | ||||||
| Zhou et al. (2019) | Women diagnosed with high-grade serous ovarian cancer (n = 50) • 25 cases • 25 controls | Ovarian cancer tissues | Illumina • V3–V4 | China | Proteobacteria | [48] |
| Firmicutes | ||||||
| Bacteroidetes | ||||||
| Actinobacteria | ||||||
| Placenta | ||||||
| Aagaard et al. (2014) | Women at term and preterm (n = 48) • One-third with preterm delivery • One-third with remote antenatal infection • One-third women delivering at term | Placental sections located 4 cm from the cord insertion site | Illumina • V1–V3 | USA | Escherichia coli | [50] |
| Prevotella tannerae | ||||||
| Bacteroides spp. | ||||||
| Streptomyces avermitilis | ||||||
| Propionibacterium acnes | ||||||
| Rhodococcus erythropolis | ||||||
| Neisseria polysaccharea | ||||||
| Neisseria lactamica | ||||||
| Fusobacterium spp. | ||||||
| Doyle et al. (2017) | Rural pregnant women (n = 1097) | Fetal (chorionic and amniotic) membrane and placental tissue near the umbilical cord insertion | Illumina • V5–V7 | Malawi | Acinetobacter spp. | [51] |
| Enterobacteriaceae | ||||||
| Prevotella copri | ||||||
| Lactobacillus iners | ||||||
| Parnell et al. (2017) | Women at term (n = 57) | Samples from the placental villous, fetal membrane, and the basal plate | Illumina • V1–V9 | USA | Ralstonia insidiosa | [52] |
| Mesorhizobium spp. | ||||||
| Lactobacillus crispatus | ||||||
| Lactobacillus iners | ||||||
| LUreaplasma nucleatum | ||||||
| Seferovic et al. (2019) | Women at term and preterm (n = 53) • 26 term deliveries • 26 preterm deliveries • 1 positive control | Dissected placental core sections | Illumina • V4 | USA | Firmicutes | [53] |
| Proteobacteria | ||||||
| Actinobacteria | ||||||
| Bacteroidetes |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peric, A.; Weiss, J.; Vulliemoz, N.; Baud, D.; Stojanov, M. Bacterial Colonization of the Female Upper Genital Tract. Int. J. Mol. Sci. 2019, 20, 3405. https://doi.org/10.3390/ijms20143405
Peric A, Weiss J, Vulliemoz N, Baud D, Stojanov M. Bacterial Colonization of the Female Upper Genital Tract. International Journal of Molecular Sciences. 2019; 20(14):3405. https://doi.org/10.3390/ijms20143405
Chicago/Turabian StylePeric, Adriana, Jürgen Weiss, Nicolas Vulliemoz, David Baud, and Milos Stojanov. 2019. "Bacterial Colonization of the Female Upper Genital Tract" International Journal of Molecular Sciences 20, no. 14: 3405. https://doi.org/10.3390/ijms20143405
APA StylePeric, A., Weiss, J., Vulliemoz, N., Baud, D., & Stojanov, M. (2019). Bacterial Colonization of the Female Upper Genital Tract. International Journal of Molecular Sciences, 20(14), 3405. https://doi.org/10.3390/ijms20143405